Indian ~. Plant Physlol •• Vol. XXIX. No.3, p.p. 230-236 (September. 1'86)
EFFECT OF LOW LIGHT INTENSITY ON GROWTH AND YIELD OF
RAINFED COTTON
DEVENDRA SINGH
"
Division of Plant Physiology and Biochemistry. Central Institute for Cotton Researcb. P. B. No. 125, G. P.O., Nagpur
SUMMARY
The response of four cotton genotypes (AKH 4 of G. oboreum L. and SRT I, B 1007 and C 1412 of G. hlrsutum L.) to reduction in light intensity by shading the plants was studied under field conditions. Reduction in light intensity reduced the rate or photosythesis as indicated by reduced NAR, (net assimilation rate) and LAB (leaf area efficiency) but LAR (leaf area ratio) and SLA (specific leaf area) increased. Dry matter of all the genotypes increased significantly with 3S percent reduction in light intensity. When it was furtl>er reduced to 6S per cent the dry matter ofSRT J and B 1007 remained unchanged, that ()f HKA 4 was the same as under natural light but reduced significantly ,in C 1412. ThO' genotypes AKH 4 and C 1412 appeared outstanding as they maintained higher seed cotton yields and higher fruiting coefficients by apportioning more photosynthates for seed cotton production even when the light intensity was reduced to 6S percent.
INTRODUCTION
Sbantz (1913) was the first to record tbat the growth of the colton plant
was the best at 1/5 to 1/2 of the normal sun light. When the crop was artificially
shaded under field conditions, Knigbt (1935) found that the yield of American
cotton was reduced by 2/3 in Sudan. Eaton and Ergle (1954) observed that
reduction in light intensity to 32 per cent of the full exposure by muslin shades
reduced seed-cotton yield by 47 per cent. In Central [ndia where cotton is sown
with the onset of south-west monsoon. Dastur and Bbatt (1956) and Dastur
and Singh (1956) opined that reduction in hours of sunsbine, during early
. growtb. acts as a limiting factor for further growth and yield. Though Canney
(1924) felt bright sunshine was essential for growth and yield in cotton. Konstan
tinov (1940) observed that varieties of cotton differ in their sensitivity to light.
Since inadequate light during the pre-flowering phase might produce subnormal
growth, it would be interesting to know how cultivated varieties of cotton
I
LOW LIGHT INTENSI1Y ON COlTON
231
react to reduction in light intensity during flowering and bolling phases of
growth when production of drymatter is maximum.
MATERIALS AND METHODS
f
Four morphologically different cotton genotypes, viz. SRT I. C 1412 and
B
1007of
G.hirsutum and AKH 4 of
G.arbore'llm., were grown in the field
during
1979·80at main research farm, CICR, Nagpur. Three levels of light
.. intensity; i.e, normai sun light-control
(850Klux), about 35 per cent reduction
in normal light-Ll (550 Klux). and about 65 per cent reduction in normal light
Ls (299 Klux), were imposed at flower initiation (65 days after sowing) and
continued till boll development
(125days after sowing) by covering the plants
with standardised clother nets. The experiment was laid in a split plot design
with different levels of light intenSity in main plots and varieties in subplots.
Each plot was replicated three times.
Growth parameters were computed by the formulae described by William
and Joseph
(1973)and the area of the leaves was estimated by dry weight method
suggested by Singh
(1975) .. RESULTS AND DISCUSSION
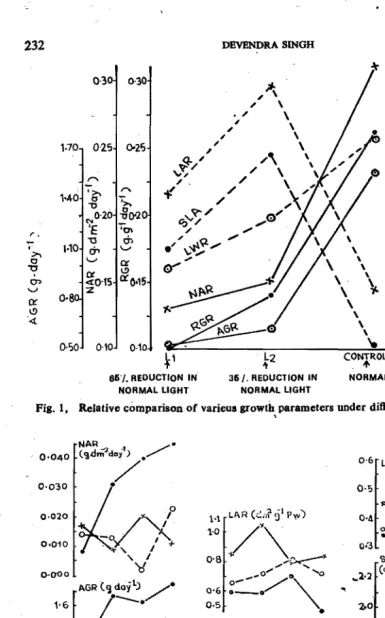
Responses of four genotypes of cotton AKH 4. SRT I, C 1412 and B 1007
to reductions in intensity of natural light under field conditions showed that
though net assimilation rate (NAR) decreased to about 66 percent (of the con
tron when light intensity was reduced either to 35 percent or 65 percent of the
normal, leaf area ratio (LAR) and specific leaf area (SLA) increased to about
50 and 40 per cent, respectively (Fig
I).
Similar findings have been reported
by Huxley
(1964);Rajan,
et
01.
(1973)and Pattarson,
et al.
(1978).C 1412
exhibited high rate of NAR (30 mg/droll/day). foJlowed by AKH 4 (21 mg/dm
2/day.) As compared to 8RT I and B 1007, the SLA was high in AKH 4 and
C
1412 (Fig 2). This indicates that leaves are thinner in AKH 4 and C 1412.
It
is further ob'erved that NAR and LAR were inversely associated (r
=-0.56)
as reported by Patterson,
et
01.
(1978)and Siog
(1975).232
..~
DBVENDRA SINGH
.,....;_L...
"";.;~'~1
.
,SRT.1 13 1007 AlCH4 C1412
~fL2 (6<;''Y.REOUCTIOI'III'l X - - l . NORMAl liGHT)
.~
-
"
~
"
) ,
0'4 '\. ".
OJ I I • 0'" , •
S~H 61007 AKII4 (1412
{
CONT~Ol. - - . {ttORtltAL llGMT
0·3 0-30
PO
0·25 0-251·40
r.
\·10
cr I.!>
~O'15
"M5:z
0·10 0·10
o·so
s',/.
REDUCTION IN 36/. REDUCTION IN2.-10 -eO
,·60
1·50
0-50NORMAL LIGHT NORMAL LIGHT
Fig. I, Relative comparison of varieus growth parameters under different light conditions.
,
LOW LIGHT 1NJE,N81TY ON COTION
233
is indicated. The genotypes AKH 4 and C 14]2 appeared outstanding as they
maintained bigber seed-cotton yield and fruitiog co-efficient by apportioning
more pbotosyntbates for seed cotton 'production even wben,the ligbt intensity
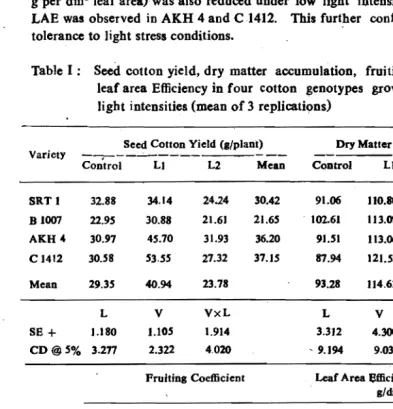
was reduced to 65 percent of the control. Leaf area
efficiency~LAE (dry matter
g per dmll leaf area} was also reduced under low light intensity Table
O.
High
LAE was observed in AKH 4 and C1412. This furtber confirms tbeir relative
tolerance to ligbt stress conditions.
Table I: Seed cotton yield, dry matter accumulation, fruiting coefficient and
leaf area Efficiency in four cotton genotypes grown under differebt
light intensities (mean of 3 replications)
Seed Cotton Yield (./plant) Dry Matter (llplant) Variety
Control LI L2 Mean Coatrol Lt L2 Mean
SRTl 32.88 34.14 24.24 30.42 91.06 110.80 IOpl 103.19
t
B1007 22.95 30.88 21.61 21.65. Un..
61 113.09 114.26 109.99AKH" 30.97 45.70 31.93 36..20 9l.S1 113.06 91.29 98.(i2 ...
~
.,
CI412 30.58 53.55 27.32 37.15 87.94 121.58 63.73 91.09'"
r
Mean 29.35 40.94 23.78 93.28 114.63' 94.25.' L V VxL L V VxL
(
S8+
].180 1.105 1.914 3.312 4.300 7.470CD@5% 3.277 2.322 4.020 ' !U94 9.035 15.695
J
•
I•
•
Fruitin. Coefficient Leaf Area Sfficiency (Dry Matter ./dm'LA)
SRT I 0.362 0309 O.~ 0.300 6.261 3.422 2.452 4.047
81007 0.225. 0.273 0.100 0.199 5.377 3.674 1.927 3.659
AICH4 0.336 0.408 0.347 0.364 10.11' 6.515 3.759 . 6.798
C 1412 0.351 0.449 0.425 0.408 9.328 5.494 4..639 6.487
Mean 0.319 0.358 0.275 7.773 4.776 3.194
L V VxL L V VxL
S8m ::I: 0.0022 0.0067 0.012 0.280 0.641 l.llO
CD@5% 0.006 0.014 0.024 0.778 1.347
NS
Reduction in chloropbyll (Fig. 3) 'was perbaps one of the contributing
factor for reduced NAR and LAE under low light intenlity. Higher rates of
234
; I.¥<
DEVENDRA SINGH
II
CONTROL (NORMAL LIGHT)O
l1
(35'Y.,
REDUCllON INNORMAL LIGln)
I
L2 (65'Y.'Ri:DUCTlON INNORMAL LIGH')
3!
u.
(T\
-"'
0"1100
-
.,.,E
B-1
00" C-1412.Fic. 3: Chlorophyll content of different cotton genotypes under different ligbt conditions.
dry matter production might be attributed to high LAR which more than
compensated the reduced NAR under reduced light intensities, as reported by
ludwig
(1965)-Further, increase in fruiting coefficient under reduced light
intensities lIlay be due to more mobilisation of assimilates from the leaf to the
boll (as revealed by increase in SLA) and better sink capacity due to more num
ber of bolls retained. High drymatter associated with fruiting coefficient resulted
in increased seed cotton yield (Asana, 1975; Bhardwaj,
et al.,
1971 and 1975).
Reduction in photosynthetic productivity at reduced ligh t intensity has also
been reported by Huxley
(1964);Lamaeva,
et al.
(1972)and Lamaeva and
Khallyeva, (1973). Reduction in photosynthesis was also found associated with
the reduction in plant growth and seed cotton yield (Bhatt and Nathan, 1917;
Bhatt and Ramanujam, 1975 and Sorour and Rassoul, 1974). Tbe effect appeared
to be more pronounced when plants
we~e
shaded during flowering and boll for
. mation phase (Lamaeva,
etal., 1972).
235
/
f
:
rLOW LIGHT INTENSITY ON COTTON
ACKN OWLEDGEMENT
Author is
highly
grateful to Director and Head Division of Plant Physio
logy and Biochemistry. CICR, Nagpur for the facilities and help provided to
conduct this study.
REFERENCES
Asana. R. D. (1975). Physiological Research in Cotton-Past and present. Indian farming. (Advances in Cotton Research). 24 : 17-19. .
Bhardwaj, S. N., Nath, V. and Mehra, R. B. (1971). Plant type in relation to fruiting coeffi cient in upland cotton (G. hlrsutum L.) Indian J. agric. Sci., 41 : 519-523.
Bhardwaj S. N., Singh, M. and Wankhede, N. P. (1975). Suitability of upland cotton varieties to closer planting and higher yield. Indian J. agric. Sci., 45 : 239-245.
Bhatt. J. O. and Nathan. A. R. S. (1977). Studies on the 8l'owth ofG. barbadoue cottons in India. Response to environmental stresses. Turrialba, rT : 83-92.
Bhatt. J. O. and Ramanujam. T. (1975). Response of the cotton plant to variations in the intensities of natural light. TU"ia/ba. 25 : 440-444.
Canney, E. E. (1924). Rain grown cotton and climate. Shirley InStitute Memoirs 3 (14) :
U.K.
Dastur, R. H. and Bhatt, J. O. (1956). Effect of chemical hormones OD carbohydrate and nitrogen content of the cotton plant. Indian Jour1lll/ of Agricultura] Science 26 : 39-79.
Dutur, R. H. and Singh, K. (1956). A study of the growth of American upland cottons in the Malwa tract of Madhya Bbarat. Indian Journal of Agricultural Science, 26: 133-192.
Eaton, F. M. and Ergle, D. R. (1954). Effects of shade and partial defoliation on carbo bydrate levels and the growth, fruitins and fibre properties of cotton pJant. Plant Physiology,29 : 39-44 •
..
Huxley. P. A. (1964). Some effects of artificialshadingoD growth of upland cotton seedings. Emp. COli. Grog. Rev., 41 : ItO-Ill.
Knight, R. K. (1935). The effect of sbade on American cotton. Empire Jour1lll/ of Experi mental Agriculture, 3 : 31-40.
Konstantinov, N. N. (1940). Light intensity as a factor in tbedeve)opment or various forml of cotton. Comptes Rendus (Doklady) de I' Academic des Science. de l' USSR, 47 : 607-609.
Lemaeva, A. M. and KhaJlyeva. K. ll973). Effect ofsbading on carbohybrade metabolism in the five fibred cotton. Invest/yo Akademi: Nauk Tuskmenollcai SSR. Bi%gi cheskikh Nauk, 2 : 26-30:
236
,.-:
~EVBNDR.A SINGH
Ludwis. L. J.. ~acld. T • .!'Dd Evans. L. T. (1965). Pbotosyntbesis in artificial communities of cotton plant io relation to leaf area. I. Experiment witb prolressiYe defaliation of mature plants. AIlSI. I. Bioi Sci. 11: 1103-1118.
Patterson. D. T., Duke. S. O. and Hoogland R. B. (1978). EfI'eet of irradiance during growtb on adaptive photosyntbetic characteristics of velvet leaf and c oUon. Plant Physiol., 61: 402-405.
Rajan. A. K.. Batteridle, B. and Blackman, G. B. (1973). DifI'erences in tbe interacting efl'ects of light and temperature on the growtb of species in the vegetatiye pbalie. Ann. Bot•• 37: 287-313.
Singh. D. (1975).Phnsiological ~analysis of yielding ability in American cottods (Golsypium hlrsutum L) ph.D. thesis IARt New Delhi. pp. 108.
Sbantz, H. L. (19l3). Tbe efl'ect of artificial shading on plant growth in Louisiana. U.S.D.A. Bureau 0/P.IQllt Indll.try Bullllln-279.
Sorour. F. A .• Rassoul. S.P.A. (1974). Meet ofsbading at different stagcs of growth of the cotton plant onllowering and fruiting. boH sbedding, yield ofseed cotton and earlincss, LibYQ1l Jr. of A,ri., 3 : 39-43.
William. C. N. and Josepb. K. T. (1973). Light use in tropical crops. measurement of growth and light in crop production in climate soil and crop production in the Humid Tropics II Bd. Oxford. Univ. Press. pp. 177 : 59-97".