The control of alternative splicing of the
leucocyte common antigen (CD45)
by
Christian Frank Zilch
A thesis submitted for the degree of
Doctor of Philosophy
March 1998
Imperial Cancer Research Fund
Tumour Immunology Unit
University College London
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10014437
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
Abstract
The leucocyte com m on antigen (CD45) is a high m olecular w eight transm em brane glycoprotein exclusively expressed on hematopoietic cells. It plays an important role in the signal transduction of T cells, in which various alternatively spliced isoforms are generated according to the stage of development and activation. Monoclonal antibodies recognising high and low molecular weight isoforms have been used to distinguish between naive and memory T cells. The splicing pattern of CD45 in T cells is altered in some individuals, who show continuous expression of high molecular weight isoforms containing exon A on activated T cells. A silent point mutation at position 77 of CD45 exon A was shown to be associated with the variant splicing pattern. This thesis provides strong evidence that this mutation is the cause of abnormal splicing.
In order to study the factors influencing CD45 alternative splicing, somatic cell hybrids of lymphocytes with a CD45 splicing defect and a mouse lymphoid line were produced. Clones expressing different isoforms o f CD45 were isolated in which expression o f the high molecular weight isoform containing exon A was associated with the mutation within exon A. In addition, minigenes including this mutation were constructed and transfected into various cell lines. Semi-quantitative RT-PCR showed a striking increase in splicing to CD45RA when com pared with the normal minigene. Taken together, these results demonstrate a causal relationship between the mutation in CD45 exon A and the variant splicing pattern observed.
Another important question is how the alternatively spliced exons of CD45 are controlled. The involvement of trans-acting splicing factors that interact with this region of CD45 pre- mRNA from minigenes was investigated by cotransfection assays of several cloned splicing factors belonging to the family of SR proteins. It was clearly shown that SRp 20 and SRp30c have antagonistic effects on CD45 alternative splicing. SRp 20 induced shifts in the CD45RA form, whereas SRp30c promoted splicing in the opposite direction in a dose dependent manner resulting in CD45RO expression.
Acknowledgements
A bove all, I am deeply indebted to my supervisor Professor Peter B everley for his extraordinary support, continuous motivation and fellowship throughout the project, without a too confining supervision. I am very grateful to all the my colleagues at the Tum our Immunology Unit for help in everyday work, especially the "CD45 team": Drs Lindsey Goff, Diana Wallace, Marcos Timon and Thomas Welge who always had an open ear and a helping hand. In particular I would like to thank Drs Mala Maini, Torben Lund and Hans Stauss for their enormous patience in critical discussions and for correcting my dreadful English. My compliments go to Mrs Christine Hughes whose efficient running of the laboratory saved me an awful lot of time and effort.
Furthermore I am very glad about to have become involved in fruitful collaborations with Dr Gavin Screaton, Andrew W alker and Gerdy ten Dam and the people who helped me during my stay at the N uffield Department of Clinical M edicine in Oxford and Bartholomew's Hospital Molecular Oncology Unit in London.
I very much appreciate the numerous blood donations which I obtained from Drs Deborah Morrison, Formwald, and Richard Jones. I should like to thank Professor Michel Streuli for his gift of various CD45 minigenes.
I am also grateful to the people of the UCL Department of Oncology for their patience and not letting me down during a smooth transition period of the good old TIU laboratory. A big "Cheers" to Lorna and Professor Avrion Mitchison for their warm hospitality.
Finally I w ould like to express my love to my family and all my friends who gave me encouragement and understanding during difficult times when I must have been unbearable.
Table of contents
A b stra ct ... 3
A cknow ledgem ents ... 4
L ist o f ab b rev iatio n s 12 D escription o f following c h a p te rs ... 16
C h a p te r 1. 17 G en eral In tro d u ctio n I. T he im m une sy ste m ... 17
Innate and adaptive immune responses... 17
Lymphocyte subpopulations...18
T lym phocytes...18
Major T cell subsets...19
Cytokine production of CD4 T cells (Th-type)...20
Naive and memory T-cell subpopulations...20
Markers to distinguish naive and memory T c e lls ... 21
Recognition of a specific antigen by the T C R ... 22
Mechanisms of T cell activation...24
Pathways of signal transduction...25
II. The Leucocyte Com m on A ntigen (C D 45)...26
Genomic structure of C D 45... 26
Glycoprotein structure of CD 45... 27
Role of CD45 in the development and activation of lym phocytes... 29
The function of variably spliced CD45 isoform s...31
CD45 in signal transduction...32
Tissue- and developmentally-specific isoforms of C D 45.... 32
Ligands of the CD45- isoforms... 33
Antibodies against isoforms of C D 45... 33
CD45 as a marker for m em ory... 34
Regulation of alternative splicing...35
III. The process o f pre-m R N A sp lic in g ... 36
Interrupted genes and molecular splicing ...36
The spliceosomal com plex...36
The mechanism of splicing...37
Cis-sequences required for constituitive splice-site recognition...41
Alternative splicing... 42
Cis-acting elements affecting splice site selection... 43
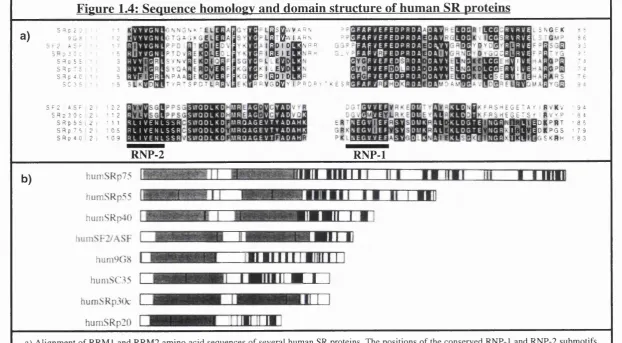
Members of the SR protein family of pre-mRNA splicing factors...44
Related proteins to the SR protein superfamily of splicing facto rs...47
RNA recognition motifs (R R M )...50
The function of SR proteins... 50
SR proteins excert non-redundant functions in vivo... 51
Activity of SR proteins in alternative splicing... 52
Alternative splicing enhancers... 52
Specific functions of RRM and SR d o m ains... 53
Shuttling of SR proteins...56
Interactions between SR proteins and RNA polymerase II... 57
SR proteins in disease states... 58
P urp o se of the study ... 58
C h a p te r 2. 59 G en eral m aterials an d m ethods M aterials 59 Media ... 59
Reagents ...60
O ligonucleotides...61
Plasmids ... 62
Yeast Artificial Chromosome (Y A C )... 62
Transgenic m ic e ...62
Bacterial strains... 63
Cell lines ...63
Human subjects... 64
M ethods 64 Tissue culture m ethods...64
Preparation of peripheral blood mononuclear cells (PBMCs) ... 65
PH A stimulation of P B L ...65
Determination of cell number and the viab ility...65
Freezing c e lls ... 65
Fractionation of cell populations using immunomagnetic beads...6 6 Production of somatic cell hybrids...6 6 Cell fu sio n ...6 6 Selection of h y b rid s...6 6 Immunofluorescence and flow cytometric analysis... 67
Lymphocyte staining for flow cytometric analysis... 67
Lipofection of adherent cells...6 8 K aryotypes...6 8 Biotinylation of DNA to serve as a probe for F IS H ... 69
Agarose gel electrophoresis...70
Purification of DNA fragments from agarose g e ls ... 70
Estimation of nucleic acid concentration...70
Ligation of restriction fragments into plasmid v ecto rs 71 Transformation of B. coli with products of ligation... 71
DNA digestion with restriction enzym es...71
Preparation of genomic D N A ... 72
Preparation of plasmid D N A ... 72
DNA sequencing... 73
Polyacrylamide gel electrophoresis of sequencing samples ...73
Site directed mutagenesis...74
Oligo labelling of DNA probes... 74
Polymerase Chain Reaction (PG R )...75
RNA methods 77 RNA preparation... 77
Reverse transcription polymerase chain reaction (R T-PC R)...77
Protein methods 78 Protein gel electrophoresis (SD S-PA G E)...78
Western blotting... 78
C h a p te r 3. E xpression o f CD45 isoform s on resting an d PH A stim ulated T cells n o rm al a n d v a ria n t p a tte rn of CD45 isoform e x p re ssio n ... 79
In tro d u ctio n 79 R esults 81 Assessment of the incidence of the variant CD45 phenotype in a normal human population... 81
Kinetics and phenotype of the variant pattern of CD45 isoform expression after T cell stim ulation... 81
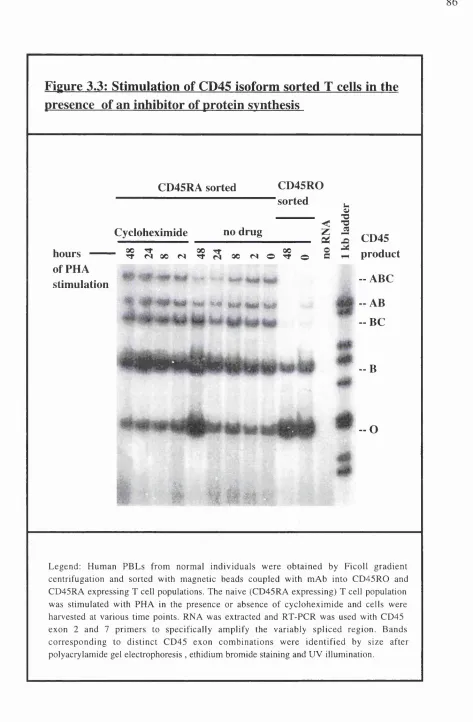
Mitogen induced CD45 isoform switch can be prevented by inhibitors of protein synthesis... 82
Discussion ...8 6 C h a p te r 4. P ro d u ctio n of som atic cell hyb rids an d th eir c h a ra c te ris a tio n ... 90
In tro d u ctio n ... 90
R esults ...93
Production and isolation of CD45 expressing somatic cell h y b rid s...93
Surface expression of human CD45 isoform s... 93
The expression of murine CD45 isoforms in the hybrids .... 94
The expression of various other human surface m arkers 95 Chromosomal analysis o f CD45 expressing hyb rid s... 97
Both CD45RO and CD45RA/RO expressing hybrids contain only one copy of the CD45-gene...97
No correlation between chromosome content and CD45 isoform expression...98
CD45RA-expression on somatic cell hybrids is associated with a mutation in exon A ... 99
Other potential mutations causing variant CD45 splicing in patients with H L H ...100
Inhibition o f CD45RA splicing in CD45-minigenes
containing the exon A mutation ... 116
Introduction 116
Results 119
Minigenes containing the exon A point mutation
show strong inhibition of splicing to C D 45R O ... 119 The exon combination ABC is predominately
retained in mutant splice p ro d u cts...1 2 0
Discussion 129
Chapter 6. 133
SR proteins affect CD45 alternative sp licin g ...133
Introduction 133
Results 135
Dose dependent promotion of CD45RO splicing
by SR p30c... 135 Effects on the alternative splicing pattern of
pSE C -LC A l-7...136 Protein expression of SRps in COS c ells... 137 Theoretical considerations for SRp30c/30a
chimeric domain swap- and SRp30c deletion-mutants 137 The atypical RRM of SRp30c is responsible for
the specific interaction with exon A ...139 Intracellular distribution of SRp30c and deletion mutants ..140
Discussion 152
Chapter 7. 158
Conclusions 158
Appendix I 168
Buffers and solutions
Appendix II 172
List of reagents
Appendix III 175
Disposals and equipment Chapter 8.
List of tables and figures
Chapter 1.
General Introduction
Tables:
Table 1.1: SR- and related proteins... 48
Figures: Figure 1.1: Some important signalling molecules on T cells... 23
Figure 1.2: CD45 genomic organisation and differential exon usage of mRNA 28 Figure 1.3: Mechanism of pre-mRNA splicing... 38
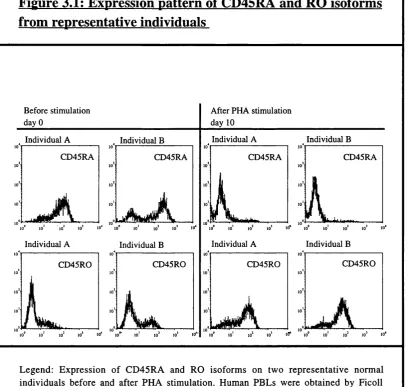
Figure 1.4: Sequence homology and domain structure of human SR proteins 46 Chapter 2. General materials and methods Tables: Table 2.1: Antibodies 60 Results Chapter 3. Expression of CD45 isoforms on resting and PHA stimulated T cells: normal and variant pattern o f CD45 isoform expression Figures: Figure 3.1: Expression pattern of CD45RA and RO isoforms from representative individuals 84 Figure 3.2: Variant phenotype of CD45 isoform expression... 85
Figure 3.3: Stimulation of CD45 isoform sorted T cells in the presence of an inhibitor of protein synthesis... 8 6 Chapter 4. Production of somatic cell hybrids and their characterisation Tables: Table 4.1 : Expression of human CD45 isoforms... 96
Table 4.2: Expression of murine CD45 isoforms... 96
Table 4.3: Expression of human surface antigens... 96
Table 4.4: Human chromosomes in human/mouse... 96
Figures:
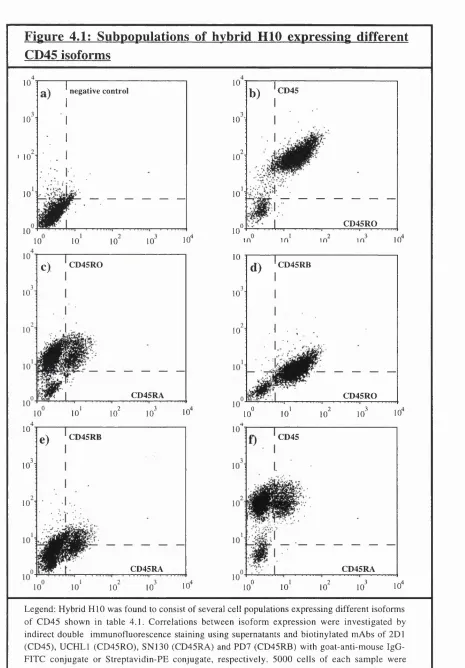
Figure 4.1: Subpopulations of hybrid HID expressing different CD45 isoforms 101 Figure 4.2: Expression of high and low molecular weight CD45 isoforms on
somatic cell hybrids H10RA+ and HI OR A-...102 Figure 4.3: Cloning of different CD45 isoform expressing HIO hybrids... 103 Figure 4.4: Detection of human CD45 in somatic cell hybrids...104 Figure 4.5: FISH studies using hybrid-probes on normal human chromosome
spreads 105
Figure 4.5.a: Chromosome content of hybrid H10RA+...106 Figure 4.5.b: Chromosome content of hybrid H I ORA-...107 Figure 4.6 : Sequence of CD45 exon A and position of C to G transition... 108 Figure 4.7: Association of the CD45 variant splicing pattern with an
exon A mutation 109
Figure 4.8: Detection of a C>G transition at position 77 of CD45 exon A ... 110 Figure 4.9: No association of the CD45 exon A mutation and members of
a family with HLH I l l
Chapter 5.
Inhibition o f CD45RA splicing in CD45-minigenes
containing the exon A mutation
Figures:
Figure 5.1: Schematic representation of the CD45 minigenes LCA-2, LCA-18, and
L C A l-7 122
Figure 5.2: Alternative splicing patterns of the minigenes
LCA-18, LCA-2 and L C A l-7 ...123 Figure 5.3: CD45 minigene splicing assay... 124 Figure 5.4: Inhibition of exon A splicing in the presence of the C>G m utation... 125 Figure 5.5: Effect of exon A (C>G) point mutation on LCA-2 splicing in CHO
and HeLa cells 126
Figure 5.6: Effect of exon A (C>G) point mutation on pSEC-LCA l-7 alternative
splicing 127
Figure 5.7: Important sequence elements within CD45 alternative exons A and C...128
Chapter 6.
SR proteins affect CD45 alternative splicing
Figures:
Figure 6.2: Co-transfection CD45 minigene splicing assay... 142
Figure 6.3: Effect of cloned SR proteins on CD45 minigene splicing... 143
Figure 6.4: Dose dependent effect of SRp30c on LCA-2 minigene splicing...144
Figure 6.5: Expression of SRp30c in COS cell transfectants... 145
Figure 6.6: Dose dependent effect of SRp30c on LC A l-7 alternative minigene splicing ... 146
Figure 6.7: Cloning strategy for SRp30a/30c domain-swap chimeras and SRp30c deletion mutants 147 Figure 6.8: SRp30a/c chimeras and SRp30c deletion mutants and their activity in CD45 minigene alternative splicing ... 148
Figure 6.9: Effects of SRp30a/30c chimeras and SRp30c deletion mutants on alternative L C A l-7 minigene splicing... 149
Figure 6.10: Expression of the SR protein 30a/30c chimeras and SRp30c deletion mutants 150 Figure 6.11: Role of structural domains in the intracellular localisation and distribution of SRp30c and deletion mutants ... 151
List of abbreviations
Ab antibody
Ac acetate
APC antigen presenting cell
ASF alternative splicing factor
ATP adenosine triphosphate
ATTC The American Type Culture Collection
auto autologous
B-LCL B lymphoblastoid cell line
BBP branch point binding protein
bp basepairs
Bq becquerel (1 disintegration/sec)
BSA bovine serum albumin
CC commitment complex
CD cluster of differentiation
cDNA complementary DNA
Ci curie (3.7 x lOl® Bq)
CMV cytomegalovirus
Con A concanavalin A
cpm counts per minute
CTD carboxy terminal domain
CTL cytotoxic T lymphocyte
Da Dalton
DAG diacyl glycerol
dATP deoxyadenosine triphosphate
DC dendritic cell
dCTP deoxycytidine triphosphate
DEPC diethyl pyrocarbonate
dOTP deoxyguanosine triphosphate
dist. distilled
DM variant individual D M.
DMEM Dulbecco's modification of Eagle's medium
DMSO dimethyl sulphoxide
DNAse deoxyribonuclease
DNA deoxyribonucleic acid
dNTP deoxyribonucleoside triphosphate
dsx DTT dTTP
E. coli
EBV EDTA EM EMBL ERE EtBr EtOH FACS ECS EEL FITC FTOC HAT hCD45R HIV HLA HLH hnRNP HPRT ICAM ICRF IFN Ig Ig IL IP3 IPTG IT AM kb kDa LB LCA LCL LG
double sex dithiothreitol
deoxythymidine triphosphate Escherichia coli
Epstein Barr virus
ethylenediamine tetraacetic acid electron microscopy
European Molecular Biology Laboratories exonic recognition element
ethidiumbromide ethanol
fluorescence activated cell sorter fetal calf serum
familial erythrophagocytic lymphohistiocytosis fluorescein isothiocyanate
fetal thymus organ cultures
Hypoxanthine, Aminopterin, Thymidine human CD45R
human immunodeficiency virus human leucocyte antigens
haemophagocytic lymphohistiocytosis high nuclear ribo-nucleo-particle
hypoxanthine guanosine phosphoribosyl transferase
intercellular adhesion molecule Imperial Cancer Research Fund interferon
immunoglobulin immunoglobulin interleukin
inositol-1,4,5, triphosphate isopropyl-thio-8-galactoside
immunoglobulin receptor family tyrosine-based activation motif
kilobase kilodalton loading buffer
leucocyte common antigen (CD45) lymphoblastoid cell line
LMA M-MLV mAb mCD45R MHC mol Mu-MLV NK NPC OD260 OD280 P PAGE PAMP PBL PBMC PBS PCR PE PEG PHA PKC PEG Pol PRR PTK PTP pu PY R RT R.T. RFLP RNA RNAse rpm RRM RT SDS SEM
low melting point agarose Moloney murine leukemia virus monoclonal antibody
mouse CD45R
major histocompatibility complex molar
murine moloney leukaemia virus natural killer
nuclear pore complex optical density at 260 nm optical density at 280 nm protein
polyacrylamide gel electrophoresis pathogen associated molecular patterns peripheral blood lymphocytes
peripheral blood mononuclear cells phosphate buffered saline
polymerase chain reaction phycoerythrin
polyethylene glycol phyto haemagglutinin protein kinase C phospho lipase C polymerase
pattern recognition receptor phospho tyrosine kinase phospho tyrosine phosphatase purine
polypyrimidine receptor
reverse transcriptase room temperature
restriction fragment length polymorphism ribonucleic acid
SF splicing factor
SH2/3 src-homologous domain 2/3
snRNP small nuclear ribo-nucleo-particle
ssc
saline sodium citratesv
simian virusSV40 Simian virus type 40
T thymine or thymidine
TAB tris-acetic acid EDTA - buffer
Taq Thermus aquaticus
TCR T cell receptor
TE tris-EDTA buffer
TEMED N,N,N',N'-Tetra methylethylenediamine
Th T helper
TIU Tumour Immunology Unit
TNF tumour necrosis factor
Tris tris (hydroximethyl-) aminomethane
Ts T suppressor
UCL University College London
UCLH University College London Hospital
UV ultra violet
V volt
vis visible
vol volume
X-Gal 5 -bromo-4-chloro- 3
-indolyl-galactopyronoside
YAC yeast artificial chromosome
Note: This thesis uses the nomenclature for CD45 reagents suggested by the 6th International
W orkshop and Conference on Leucocyte Differentiation Antigens, Kobe, Japan, November 1996.
Description of following chapters
In chapter 3 of this thesis the CD45 isoform switch in stimulated T cells was investigated and im m unofluorescence was employed to detect the variant phenotype of CD45 alternative splicing. Furthermore the incidence of this genetic defect was evaluated.
The data presented in chapter 4 provides evidence for the involvement o f the structural gene of CD45, but not a postulated splicing factor in the emergence of the variant splicing pattern. Somatic cell hybrids were produced to separate the two alleles o f CD45 and a mutation in exon A was found to be associated with hybrids displaying high molecular weight isoforms of CD45.
The proof for this mutation being the underlying cause of the variant phenotype in humans is provided in chapter 5 following the transfection of several different minigenes into which the mutation had been introduced that failed to splice to CD45RO forms properly.
The regulation o f CD45 alternative splicing was studied in chapter 6: particular splicing factors belonging to the family of SR proteins were shown to have antagonistic effects on alternative CD45 splicing, either promoting or inhibiting the exclusion of exon A. Domain swap and deletion experiments extended the analysis of SR proteins in their ability to change alternative CD45 splicing providing evidence for the specificity of different domains within one individual splicing factor (SRp30c).
Chapter 1
Introduction
I. The immune system
We encounter an environment that contains a great variety of infectious microbes which can cause life threatening disease, if they multiply uncontrolled. Therefore we need a biological defence system which combats pathological invaders and can distinguish between self and non self. This ability to recognise and respond to the diversity of infectious agents in a controlled way is one of the hallmarks of our immune system.
Innate and adaptive immune responses
Immunity is provided by specialised cells and the molecules they produce and secrete. Two different, but interrelated types of immunity, depending on the specificity and mechanism of the response can be distinguished: the innate and the adaptive immune systems. Nonspecific components of innate immunity protect the host from pathogen associated effects and are characterised by rapid response kinetics. This first barrier o f protection is based on nonspecific cellular as well as soluble factors, including the complement system, natural killer cells, antibacterial and viral products secreted by phagocytes, including granulocytes and macrophages. The innate immune system operates via germline-encoded receptors, which are commonly referred to as pattern recognition receptors (PRRs) (M edzhitov and Janeway, 1997a; M edzhitov and Janeway, 1997b). These receptors are able to recognise pathogen associated m olecular patterns (PAMPs), a set o f conserved m olecular structures that are shared by a large number of pathogens. They include humoral proteins circulating in the plasma, endocytic receptors expressed on the cell surface and signalling receptors that are either cell membrane associated or localised intracellularly (Fearon and Locksley, 1996). A recent example of a conserved defence pathway found in both insects and vertebrates is mediated via the Toll receptor, that can induce the production of cytokines and costimulatory molecules by induction of NFkB upon stimulation (Lemaitre et al., 1996; M edzhitov et al.,
1997). The protection provided by the innate immune system however is limited due to its restricted specificity and inflexibility to adapt to pathogens that developed escape mechanisms through a high mutational rate.
against previously encountered pathogens. Immunological memory is the capability of the immune system in a repeated confrontation with the same antigen to mount a secondary immune response even after years. In contrast to the primary immune response, the secondary response is faster and the reaction to the antigen is stronger and more prolonged than at the first antigenic contact. The adaptive immune system uses receptors that are generated by somatic rearrangement of their genes, creating an immense repertoire for antigen recognition. Therefore the specificities of receptors expressed on a distinct subset of cells is not predeterm ined and an immune response will only be generated if this cell encounters an appropriate antigen. However, immunopathologies may develop if the adaptive immune system is inappropriately activated. Consequently it requires signals that provide information about the origin of the antigen, which appears to be a function of the innate immune system (Medzhitov and Janeway, 1997a). These signals are provided by APCs and epithelial cells and include immunological mediators, such as inflammatory cytokines and chemokines as well as the co-stimulatory molecules, which activate the specialised cells of the adaptive immune system. Adaptive immunity is mediated mainly by two types of lymphocytes acting together with phagocytic cells, macrophages and dendritic cells, which take up foreign material, process and present antigens to T lymphocytes.
Lymphocyte subpopulations
Lym phocytes are divided into two m ajor subgroups, the T- and B -lym phocytes. B- lymphocytes mature in the bone marrow and migrate between the blood and lymphoid tissues. B-cells produce antibodies, which can be membrane bound or secreted, bind antigens and trigger effector mechanisms for their destruction and removal. T-lymphocytes develop in the bone marrow and subsequently migrate to and mature in the thymus. Once matured they circulate between the blood, lymphoid and non-lymphoid tissues and execute a variety of functions. One group interacts with B-cells and helps them to divide, differentiate and make antibody. Another group interacts with phagocytes supporting them to destroy pathogens. These two groups are called T-helper cells. In contrast cytotoxic T-cells are responsible for destruction of host cells which become infected by viruses or other intracellular pathogens and even tumour cells.
T lymphocytes
thymus, either promoting selection for maturation to CD4+/CD8+ single positive thymocytes or depletion by apoptosis (Anderson and Perlmutter, 1995; Jameson et a l, 1995; Sprent et al., 1988) Both positive and negative selection must depend an TCR interaction with self peptide in conjunction with the MHC complex as this is all there can be in the thymus. Once mature, T cells migrate out of the thymus and recirculate between the blood and lymphoid tissues. W hen T cells encounter an antigen they undergo clonal expansion and acquire effector and memory functions.
Antigens are normally presented as peptides on the major histocompatibility complex (MHC) expressed by antigen presenting cells (APC) such as macrophages, dendritic cells or B cells. T cells in turn can provide help to these other cellular components of the immune system by the secretion o f a variety of mediators or directly eliminate pathogens depending on the subset. After massive increase in circulating antigen-specific T cells after stimulation, the majority of these cells are cleared as they undergo apoptosis (Russell et al., 1991; Zhang et al., 1995) or persist in a relatively quiescent memory state (Sprent and Miller, 1976; Sprent and Tough,
1994). Apoptosis is associated with down-regulation of expression of the protooncogene Bcl- 2 (Cory, 1995), which can be triggered by deprivation of certain cytokines (Akbar et al., 1996; Maini et al., 1998 : submitted)
Major T cell subsets
T-lym phocytes consist of heterogeneous subpopulations of cells which can be identified phenotypically by their expression of different cell surface molecules. All T cells express on their surface the T-cell-receptor complex (TCR-complex) for the recognition of antigens presented by APCs as peptides bound to M H C -m olecules (Fink and B evan, 1978; Zinkernagel et al., 1978). Two major functionally distinct lineages of T cells are defined by the expression of co-receptors CD4 or CD8. The CD4 molecule identifies T-helper cells (Th)
and the CD8 molecule the cytotoxic T-cells (Tc) subpopulation. Suppressor cells (Ts) can be
identified only by their function and not by the expression of any unique surface molecule.
C D8 and CD4 m olecules interact w ith domains o f the M HC I- or MHC Il-com plex,
positively affects the response of B and T cells and another type can induce delayed type hypersensitivity responses.
Cytokine production of CD4 T cells (Th-type)
T cells expressing CD4 can be further subdivided into ThO, T hl and Th2 cells according to their profile of cytokine secretion (Mosmann et ah, 1986). T h l type T cells predominately activate macrophages mainly by the release of y-IFN, IL-2, IL-12 and TNF-p. In contrast, Th2 type T lymphocytes generally provide help for B lymphocytes to differentiate into plasma cells by the secretion o f IL-4, IL -6, IL-10 and TGF-P (Abbas et ah, 1996; Fearon and
Locksley, 1996; Romagnani, 1992; Scott, 1993).
Although, the immune modulatory effects of different cytokines are extremely complex, many autoimmune diseases involving tissue damage are characterised by T h l type responses, whereas allergic reactions are associated with Th2 type responses. The ThO type is believed to represent both a Th2 and T h l type cytokine profile. There is evidence that suggests that this bipolar phenotype can be polarised by the influence of certain cytokines (Hsieh et ah, 1992). IL-12 and y-IFN were shown to direct the bipolar cell towards T h l, whereas IL-4 promotes the production of the Th2 phenotype. Furthermore y-IFN negatively affects Th2 type cytokine production, whereas IL-10 indirectly inhibits the T hl cytokine pattern.
Naive and memory T-cell subpopulations
Peripheral mature T-cells can also be divided into naive and memory cells, depending on their stage of development. Naive T-cells have not been in contact with any specific antigen, whilst memory T-cells have already encountered an antigen and show enhanced responsiveness to recall antigens (Cerottini et ah, 1974). Immunisation can substantially increase the frequency of T cells responsive to specific antigen (MacDonald et ah, 1980), consistent with the theory of clonal expansion of antigen primed T cells in vivo (Jerne, 1984). The duration o f antigenic stimulation determines whether T cells become activated and this requirement differs largely between naive and memory T cells (lezzi et ah, 1998).
Schittek and Rajewsky, 1990). There is accumulating evidence from studies in mice though, that the ability o f the immune system to create memory cells does not require the preceding formation of effector cells (Bemer et ah, 1995; Liu et ah, 1997).
A central question is whether the cells responsible for immunological memory itself are long lived or if memory is rather determined by specific clones that persist for long periods, but contain rapidly cycling cells. The first studies in humans using mathematical models revealed that the lifespan of the memory T cell population is paradoxically shorter than in the slower cycling naive T cell population (Michie et al., 1992). This was further supported by the observation that CD45RO+ T cells expressed high levels of Fas (CD95) and downregulated Bcl-2 expression, rendering them prone to apoptosis (Akbar et al., 1993; Salmon et al., 1994). These observations have proven difficult to reconcile with the view that the carriers of memory are imbued with the quality of long life, but are consistent with the idea that memory is carried in long lived clones of T cells. New evidence from clonally expanded CD8+ T cells
in acute infectious mononucleosis shows a paradoxical preservation of telom ere length accompanied by profound upregulation of telomerase, suggesting that these T cells can be rescued from apoptosis by certain cytokines hence entering the memory pool with a high capacity for continued cell division and allowing longterm maintenance of immunological memory (M. Maini, 1998 submitted).
Another recent debate concerns the requirement of antigen for persistence of a memory T cell population. The original assumption for the existence of a longlived resting T cell population was first questioned by observations following transfer of T cells that their survival critically dependent on the persistence of antigen. (Gray and Matzinger, 1991; Gray and Skarvall, 1988). However, this field has been highly controversial, since later studies based on virus specific C D8+ T cells show either antigen dependent survival (Gehen et al., 1992; Zimmerman et al., 1996) or memory endurance in the absence of antigen (Hou et al., 1994; M ullbacher, 1994). However these discrepancies may be due to the fundamentally different experimental models (Kundig et al., 1996), questioning the reliability of in vitro techniques to study this phenomenom.
Markers to distinguish naive and memory T cells
and CD44 isoforms) the lymphocyte homing receptor (CD62L), Fas (CD95), co-stimulatory molecules (CD27, CD28), certain cytokines (IL-2, IL-4, IFN-y, T N F -a) and their receptors (IL-2R ). The surface m olecules are not exclusively expressed on one or the other subpopulation, as is the case for CD4 and CDS cells, but rather expressed at different densities.
Apart from the change in expression levels, differences in the pattern of alternative splicing (especially for CD45) have been associated with a functional change from naive to m em ory/effector T cells. The enhanced expression of surface adhesion m olecules and alterations in alternative splicing (especially for CD44) on memory T cells results in a different circulation pattern in the organism. Both activated and resting T cells are found in efferent lymphatics and re-enter the blood via the thoracic duct. However, whilst naive T-cells are mostly found in blood vessels and enter lymph nodes, memory T-cells can leave the blood to traffic through tissues and pass via afferent lymphatics to enter lymph nodes (Mackay,
1993; Mackay, 1991; Mackay et al., 1990; Mackay et al., 1992)
Recognition o f a specific antigen by the TCR
T- and B-cells have basic differences in the way they recognise an antigen. B cells recognise and bind antigens in their native form. T-cells are only able to recognise antigens via the TCR, when they have been processed and presented by accessory cells in association with M HC-molecules (Sharma et al., 1996; Shimonkevitz et al., 1983; Zinkernagel, 1978). Only if the TCR has the appropriate binding site is the T-cell activated through the TCR/CD3 complex. The interaction of the TCR with the appropriate MHC complex is referred to as self- M HC-restriction (Zinkernagel and Doherty, 1974).
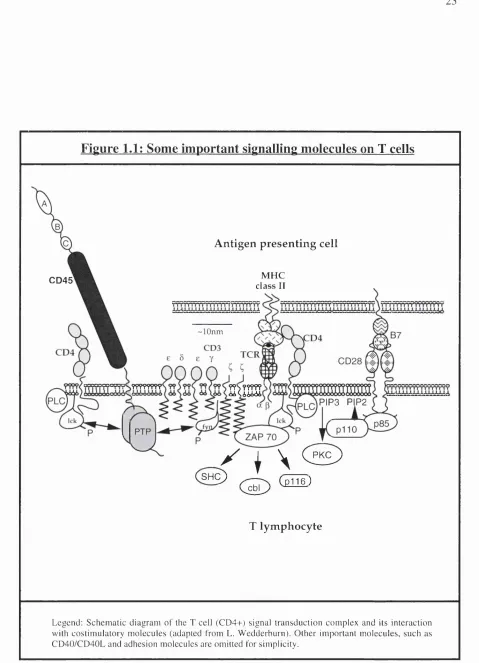
Figure 1.1: Some important signalling molecules on T cells
A n tig en p re s e n tin g cell
M H C
class II C D 45
-lOnm
????????
KIHÏÏ
P IP 3 P P 2
( S H ^
/
I \ (
2
^
( S )
( p 1 1 6 )T ly m p h o cy te
The diversity o f the TCR repertoire is generated by rearrangement of the V, D, J and C segments during T cell ontogeny, to produce a continuous V-D-J sequence, which is then spliced together with the C region after transcription. The germ line DNA consists of approximately 50 V a and 60 Vp segments, over 70 J a and Jp segments, one Dp segment, a single C a and two Cp segments creating a enormous variety o f more than 10^^ different com binations for the ab TCR by recombination, if N region heterogeneity is taken into account (Moss et al., 1992).
Mechanisms of T cell activation
In order to generate a T cell response, the TCR must have appropriate specificity and sufficient affinity for peptides presented in conjunction with the self-restricted MHC receptor. During this process only specific clones of T cells become activated. However, mitogenic products of some bacteria and plants, termed superantigens, can produce a polyclonal response within a subset of a T cell population that contains V gene products with which they interact (Choi et al., 1989; Bowness et al., 1992). Normally, the functional outcome o f TCR ligation is dependent on the presence or absence of costimulatory signals provided by APCs (Janeway and Bottomly, 1994; Linsley and Ledbetter, 1993). As a result of this additional requirem ent, antigenic stimulation can either lead to a productive cellular response and acquisition of effector function or in its absence, to a state of anergy, which is characterised by long lasting unresponsiveness (Jenkins and Schwartz, 1987; M ueller et al., 1989).
A critical role in the prevention of or induction of anergy is brought about by costimulatory molecules such as the B7 family, B7-1 (CD80) (Freeman et al., 1989), and B7-2 (CD8 6) (A zum a et al., 1993), CD40 (Grewal and Flavell, 1996) a large num ber o f adhesion molecules, such as CD2 (Springer, 1990), CD5 (Ceuppens and Baroja, 1986; Osman et al., 1993), CD54 (Altmann et al., 1989) and CD58 (Moingeon et al., 1989), as well as the soluble ligands (Ceuppens et al., 1988; Lorre et al., 1994).
The B-7 family of molecules display a restricted pattern of expression on APCs and can be induced by a variety of stimuli (Boussiotis et al., 1994). Both B7-1 and B7-2 interact with either CD28 (Harding et al., 1992) or CTLA-4 (Brunet et al., 1987; Linsley et al., 1991) expressed on T cells constitutively or after activation, respectively. However, different functional effects were suggested, as B7-1 preferentially induces Th2, whereas B7-2 predominantly influences T hl-m ediated responses (Kuchroo et al., 1995; Lenschow et al.,
B 7-CD 28 interaction appears to be closely linked to CD 40-CD40L interaction, since signalling through CD40, expressed on most APCs, can trigger upregulation of B7 (Caux et al., 1994; Kiener et al., 1995). CD40-CD40L engagement is not only critical for B cell growth, isotype switching and Ig-synthesis (Banchereau et al., 1994), but can also lead to T cell activation enhancing both T h l and Th2 type responses (Peng et al., 1996; van Essen et al., 1995). In the light o f this recent data a reciprocal stim ulation model for T-B cell interaction has been proposed, in which TCR-Ag/M HC engagement can induce CD40L upregulation on T cells, which in turn interacts with CD40 resulting in increased B7 expression on the cognate B cell. As a consequence of B7 upregulation, B cells reciprocally trigger the T cell via CD28. The two principal costimulatory signals are therefore provided by B7-CD28 and the CD40-CD40L interactions whereas adhesion molecules predominantly exert their costimulatory role by increasing the avidity of T cell and APC contact (Foy et al.,
1996).
Pathways o f signal transduction
Binding of MHC/peptide complexes to TCR/CD3 molecules mediates signalling, perhaps via oligomerisation and conformational changes, that can result in activation, anergy or apotosis, depending on the state of T cell differentiation and the influence of co-receptors (Cohen, 1992; M iller and Morahan, 1992; Weiss, 1991). The strength of these signals and the presence or absence of costimulatory signals determines the activation o f downstream src-family members, such as p59^y^, p56l^^ and src-like ZAP-70, all tyrosine kinases that couple the TCR to downstream pathways of signal transduction via CD3 and Ç molecules (Cantrell, 1996; Chan et al., 1994; Penninger et al., 1993). The tyrosine kinase p59fy^ directly associates with the components of the TCR/CD3 complex, whereas p56^^^^ is associated with CD4 or CD8, which bind the appropriate MHC-complex with their extracellular domain . The
activation o f this so called phosphotyrosine kinase (PTK) pathway results in tyrosine phosphorylation of a variety of cellular proteins (Samuelson et al., 1986; Hsi et al., 1989). The sequence o f these events is activation of src kinases, that catalyse the phosphorylation of IT AMs (immunoglobulin receptor family tyrosine-based activation motifs), which then recruit and activate ZAP-70/syk kinase by phosphorylation (Iwashima et al., 1994; Thome et al., 1995; Weil et a l, 1995).
A mong those proteins that undergo tyrosine phosphorylation is the y l isoform of phospholipase C (PLC) increasing its catalytic activity (Park et al., 1991; Weiss et a l, 1991) and inducing p36 tyrosine phosphoprotein engagement, which is necessary to recruit PLCyl to the plasm a membrane (Sieh et al., 1994). The enhanced activity of PLCyl leads to the h y d ro ly sis o f the m em brane phospholipids p h o sph o ino sitolp h osph ate (P IP2) to inositoltrisphosphate (IP3) and diacylglycerol (DAG) (Nisjibe et a l, 1990; Kim et al., 1991).
kinase C isozymes (PKCs) and mobilisation of intracellular stored calcium ions (Berridge et ah, 1984; Imboden et ah, 1985). These proximal events subsequently lead to activation of ras, raf and M AP kinase pathways further transmitting the signal to transcription factors such as fos and jun. These bind directly to the DNA strands and recruit polym erases to start transcription of genes that are important for cellular proliferation and differentiation (Cantrell,
1996).
An alternative signalling pathway from the TCR involves inositol lipid metabolism substrates, which are phosphorylated by the large catalytic subunit o f phosphatidylinositol 3'-hydroxyl kinase (Ptdlns 3-kinase) (Ward et ah, 1992), upon T cell activation. CD28 is thought to provide a costimulatory signal which is essential for the optimal activation of this enzyme (Ward et ah, 1993). The small regulatory subunit of Ptdlns 3-kinase can directly associate with the cytoplasmic domain of CD28 (Pages et ah, 1994; Truitt et ah, 1994), which may result in direct interaction with the TCR Ç chain (Exley et ah, 1994) or alternatively src- kinases such as p59fy^ and p56^c^^ that interact with components of the TCR complex (Prasad et ah, 1993a; Prasad et ah, 1993b). The downstream signal transduction cascades for Ptdlns 3- kinase, w hich result in the cellular responses, of differentiation and growth are very complicated and not completely understood (Cantrell, 1996). However, another protein kinase p70S6, involved in protein synthesis as well as members of the PKC family have been suggested as targets (Liscovitch and Cantley, 1994; Ming et ah, 1994; Monfar et ah, 1995). Important accessory molecules of the T cell acting at the most proximal point to potentiate of this TCR signalling cascade are CD2, CD4, CD8, C D lla , CD28, (CD54, CD58) and the
leucocyte common antigen, CD45.
II. The Leucocyte Common Antigen (CD45)
CD45 is a membrane associated high molecular weight glycoprotein expressed exclusively by hem atopoietic cells, which is im portant for signal transduction in T as well as in B lymphocytes (Thomas, 1989; Thomas and Lefrancois, 1988; Trowbridge and Thomas, 1994).
Genomic structure of CD45
1997). The extracellular domain of CD45 is encoded by exon 3, the amino terminus, and exons 7 through 15 (exons 1 and 2 are untranslated regions).
The CD45 isoforms arise by alternative splicing of exons 4, 5 and 6 (also designated as A, B
and C), which are used differentially to generate potentially eight different mRNAs (Thomas et al., 1987; Saga et al., 1987; Streuli et al., 1987). Six of the eight possible transcripts have been isolated as cDNAs from three different species: human, mouse and rat (Ralph et al., 1987; Saga et al., 1987; Streuli et al., 1987). The most prominent isoforms contain exons A, B and C, only exon B or none of the variably spliced exons. The transmembrane peptide is encoded by exon 16. Exons 17 through 24 encode the first and exons 25 through 32 the second of the two homologous domains of the cytoplasmic region. The carboxy terminus is encoded by exon 33, which also includes the entire 3' untranslated sequence o f CD45 (for illustration see figure 1.2).
Glycoprotein structure of CD45
The complete sequence for human, mouse and rat CD45 has been determined from the analysis of cDNA clones (Barclay et al., 1987; Ralph et al., 1987; Thomas et al., 1987; Saga et al., 1987; Streuli et al., 1987). The glycoprotein is composed of an amino-terminal, external domain ranging in length from 391-552 amino acids, depending on the isoform, a membrane spanning region of 22 amino acids and a large cytoplasmic domain of 705 amino acids. The cytoplasmic domain is highly conserved between species and is the largest reported to date corresponding to an approximate molecular weight of 80 kDa (Thomas, 1989).
Figure 1.2: CD45 genomic organisation and differential exon usage
of mRNA
CD45 gene 130 kb
19 20 22 30 31 (A )5 (B ^ 6 (C ^ || Y | ) l | 1^ I j l j t 1^ 16T M ^1 | 18^ 2 1 || 3 2 4 ^ 5 ^ 6 27.2
//-i-m
□ □ #
II
3 A
mRNA splicing ev en Is
C 7
C 7
glycoprotein Fragment length m.w. (kD) afler PCR (bp)
1)CD45R ^
----2) C045R
3) C045R
4) C045R
3 A
AB
AC
BC
5) C045R ^
6) C045R 0
7) C045R,
8) CD45R
3 A
mouse human 2 2 0 5 8 0 6 7 5
2 1 0 4 3 9 531
2 1 0 4 3 3 5 3 4
2 1 0 451 4 7 7
1 9 0 2 9 2 3 9 0
1 9 0 3 1 0 3 3 3
1 9 0 3 0 4 3 3 6
1 8 0 1 6 3 1 8 2
The external part of CD45 had been most extensively studied in rat thymocytes (Sunderland et ah, 1979; Thomas et al., 1985; Barclay et al., 1987; Brown et al., 1981). It is much less conserved than the cytoplasmic domain and divides into at least three different subdomains: a region containing O- linked carbohydrate and two separate cysteine rich regions. It contains at least 14 potential N-linked carbohydrate sites, all but one of which are heavily glycosylated. The sequence, encoded by the variable exons A, B, and C, is rich in serines and threonines. The proline content o f about 14 % and the absence of cysteines indicates a random structure, which is characteristic o f 0-linked carbohydrate sites (Davis et al., 1986). The usage o f the three variable exons changes the number of potential 0-glycosylation sites. The largest form o f the molecule is distinguished from the smallest by an insert of 161 amino acids after the first eight amino-terminal residues.
Role of CD45 in the development and activation o f lymphocytes
CD45 was found to be a growth regulator required for antigen stimulated proliferation. The first indications for this role were provided by antibody crosslinking studies, in which the ligation o f CD45 during T cell stimulation resulted in either potentiation or inhibition of T cell activation, depending on the CD45 antibody or T cell line examined (Ledbetter et ah, 1988; M arvel et al., 1989; Schraven et al., 1989; Turka et al., 1992). This is in line with studies using CD45-deficient cell lines indicating that CD45 is a positive regulator o f TCR signalling that cannot be substituted by other PTPs expressed by T-cells (Koretzky et al., 1990; M cFarland et al., 1993; Pingel and Thomas, 1989; W eaver et al., 1991). N orm al responses were restored by the transfection of different CD45 constructs into the deficient cell lines (Koretzky et al., 1990) (Shiroo et al. 1992; Desai et al., 1993).
These studies were incompatible with results from aggregation of the TCR and CD45 with antibodies, which consistently resulted in the inhibition of T cell activation, suggesting CD45 to be a negative regulator of TCR signalling (Ledbetter et al., 1991).The paradox arising of these contradictory data was explained by the need for TCR oligomerisation for proper signal transduction (Ledbetter et al., 1986; Ratcliffe et al., 1992). Any perturbation that prevents this outcome, such as CD45/TCR crosslinking, was believed to inhibit signalling (Alexander et al., 1992). This hypothesis was supported by careful studies using CD3/CD45 antibody constructs, demonstrating that inhibition of signalling does not occur. The same result was achieved when multivalent IgM mAbs were used (Shivnan et al., 1992).
splicing. Possibly as a result of this increased frequencies of mature T cells were observed in some organs. Therefore differences between the two models have to be considered cautiously.
Still, both CD45 - "knockout" mouse models showed severe impairment in thymic T cell development, which was arrested in the CD4+/CD8+ double positive stage before positive and negative selection occurs. Two distinct checkpoints in thymic development appear to be affected by the lack o f CD45. Firstly, the ability to transit from the CD4-/CD8- double negative, TCR- negative to the CD4+/CD84- double positive, TCR+ positive thymocyte is greatly reduced. Secondly, m aturation o f this im m uature into the positively selected thymocyte giving rise to single positive CD4-CD8-I- or CD4+CD8- cells is inhibited The few mature T cells that were detected in CD45 -/- null mice showed no response to CDBmAb mediated stimulation. This defect is probably due to inactivation of the src- kinases p59^Y^ and p5 6^^k by hyperphosphorylation, losing their capability to recruit downstream ZAP-70
kinase to the TCR-Ç chain.
Even though signal transduction through the TCR is still possible in the absence o f CD45, its presence appears to reduce the overall threshold of TCR mediated activation of T cells (Stone et al., 1997). This is in line with experiments using CD45 deficient cell lines, in which the basal level of p59fy^ and p56^^^ phosphorylation was inversely related to kinase activity, suggesting that the role of CD45 is to control these kinases by dephosphorylation (Hurley et al., 1993; McFarland et al., 1993; Ostergaard et al., 1989). Both p59^Y^ and p56^^^ have been shown by immunoprécipitation experiments to colocalize with CD45, supporting the idea that these kinases represent the major in vivo substrates for CD45 (Mustelin and Altman, 1990; Schraven et al, 1991; Mustelin et al, 1992; Shiroo et al, 1992).
Both, CD45 exon 6 and exon 9 CD45 -/- null mouse models have also provided insights into the relevance of CD45 in B cell maturation and activation. Surprisingly, B cell development was not affected by the absence of CD45. However, in mature B cells, signal transduction through the B cell antigen receptor was greatly im paired (K ishihara et al., 1993) but proliferation in response to CD40-mediated signals appeared to be normal (Byth et al., 1996). The increase in the number of B cells that was consistently detected in CD45 exon 9 -/- null mice, but not in CD45 exon 6 -/- null mice was suggested to be a result of the expansion of distinct B cell subpopulations with unique characteristics, such as high expression levels of IgM (Byth et al., 1996). In conclusion, CD45 is an important positive regulator for T cell generation as well as for apoptosis. In contrast, it is clear that CD45 is not involved at any stage during the development of B cells. However in mature lymphocytes it appears to play an critical role in signal transduction through both the BCR and TCR.
The function of variably spliced CD45 isoforms
Although the functional importance of CD45 is well established, much less is known about the role o f individual CD45 isoforms. There is substantial evidence that CD45 can associate with a number transmembrane signalling molecules on T cells (Altevogt et al., 1990; Mittler et al., 1991; Schraven et al., 1989; Torimoto et al., 1991). Distinct isoforms may differ in their ability to engage in TCR mediated signalling, as some were found to interact preferentially with the TCR and its coreceptor CD4 (Dianzani et al., 1990). M oreover, several studies indicated that elevated expression of CD45RO isoforms correlate with increased sensitivity to antigenic stimulation in CD4 T cells (Byrne et al., 1988; Sanders et al, 1989; Horgan et al,
1990) and that activation requirements differ in CD45RO versus CD45RA expressing T cells (Luqman and Bottomly, 1992; Patel et al., 1994; Robinson et al., 1993).
new role for specific CD45 isoforms in T cell trafficking. These results suggest that a change in CD45 isoform expression from CD45RA to CD45RO may render the TCR and accessory m em brane receptors more accessible, thereby increasing previously low M HC/Ag-TCR affinities, resulting in the more efficient activation of T cells at a considerably lower threshold and less requirement for co-stimulatory signals.
CD45 in signal transduction
The src related protein p56^^^ was found to be one of the physiological substrates o f CD45 dem onstrated by analysis of tyrosine phosphoprotein patterns in CD45 deficient cells (O sterg aard et al., 1989; Sieh et al., 1993). Supporting data cam e from co- immunoprecipitation studies, in which Ick coprecipitates with CD45 (Schraven et al. 1991). Also p59fy^ is regulated by CD45 in associating to the TCR/CD3 complex (Shiroo et al. 1992; R othstein et al., 1993; M ustelin et al., 1992). These two substrates appear to be specifically dephosphorylated by CD45, because CD45 was unable to exhibit its phosphatase activity in vivo on src, ubiquitious membrane bound protein kinase (Hurley et al., 1993). These results together suggest that the previously described signalling defect in CD45- deficient cells is due to an inactivation of Ick and/or fyn by hyperphosphorylation o f a negative regulatory C-terminal tyrosine residue interacting with the SH2 regions of the proteins. The generation of CD45 -/- null mice has provided further insights into the function o f CD45 in signalling pathway. These results are largely consistent with those of earlier studies already described.
Tissue- and developmentally-specific isoforms of CD45
The extracellular domains of CD45 are highly polymorphic between different leucocyte lineages ranging in molecular masses from 220 kDa on most B-cells to 180 kDa on T cells. The expression o f alternatively spliced isoforms depend on the cell type, the stage of developm ent and activation (Johnson et al., 1989; Streuli et al., 1987). M ost thymocytes express the lowest molecular weight isoforms of 180 kDa and 190 kDa (B only isoform) (Lefrancois et al., 1986; Janossy et al., 1989), but the more m ature cells found in the medullary region carry the CD45RA isoform on their surface. T lymphocytes can be divided into two functionally distinct subpopulations on the basis of CD45 isoform expression (Akbar et al., 1988; Morimoto et al., 1985; Rudd et al., 1987). Stimulatory signals result in a change of the isoform expression from CD45RA to CD45RO in mature T cells (M erkenschlager and Fisher, 1991).
expression o f only one or two of the variable exons (Hathcock et al., 1992). The lowest molecular weight isoform CD45RO is characteristic of granulocytes and monocytes, whereas CD45RB isoform expression was found on m ost lym phocytes (Pulido et al., 1988). Interestingly, no differences were detected among T cell populations and lines in the relative expression of particular variable exons, as exon B is always found in greater abundance than exons A or C. Among the possible combinations of two variable exons, AB and BC but not AC were detected in T cells (Rogers et al., 1992).
Ligands o f the CD45- isoforms
The search for ligands of the CD45 isoforms has not been successful to date. In 1992 some experim ents suggested an interaction between CD45RO and the B lymphocyte adhesion molecule CD22 (Stamenkovic et al., 1991), but subsequent studies have shown that CD22 recognises a -2 -6 sialic acid containing carbohydrate structures displayed by multiple cell surface molecules (Engel et al., 1993; Powell et al., 1993; Sgroi et al., 1993).
Antibodies against isoforms of CD45
A num ber of monoclonal antibodies to epitopes of the alternative spliced exons A, B, C and invariant regions of human CD45 have been isolated (Smith et al., 1986; Dalchau and Fabre, 1981; Coffman and W eissman, 1981; Spickett et al., 1983; Streuli et al., 1988) and allow determination of isoform expression on human leucocytes. The mAb U CHLl reacts with the lowest molecular weight isoform CD45RO of 180 kD; several mAbs detect the CD45RA epitope present in high molecular weight isoforms of 205 and 220 kD (Akbar et al., 1988) (Serra et al., 1988; Clement et al., 1988; Lee et al., 1990; Clement et al., 1992).
et al., 1988; Akbar et al., 1989) and appears to progress through interm ediate CD45RB isoforms (Salmon et al., 1994).
CD45 as a marker for memory
The view that CD45RO expression correlates with the memory/effector type T cell had been widely accepted for a long time (Mackay, 1991; Sanders et al., 1988; Sanders et al., 1988), but has recently been challenged (Zimmerman et al., 1996). The first evidence that raised doubts of the suitability of low molecular weight CD45 isoforms as memory markers was provided by in vivo data in rats showing that high molecular weight isoforms could be re expressed after the acquisition of memory functions (Bell et al., 1998; Bell et al., 1992). The memory T cell population in mice is most likely to follow a similar fate (Sprent, 1993). However naive and memory type T cells are defined differently in human, mice and rats, depending on the specificities of the mAbs used and may therefore not represent similar subsets of T cells.
Nevertheless, a body of evidence that this notion may also apply to humans has accumulated showing that T cells can revert from the CD45RO to the expression o f double positive CD45RA/RO phenotype (Rothstein et al., 1991; Sarawar et al., 1993; Warren and Skipsey,
1991). Furtherm ore the existence of antigen specific CD84- T cells that lack CD45RO expression has been described (Unutmaz et al., 1994). More recent studies concentrating on the role of CD45 isoforms as differentiation markers provided evidence that human CD8-f- memory/effector type T cells can be similarly subdivided into two subpopulations, depending on the expression of CD45RA, the absence of co-stimulatory molecules and the ability to secrete IL-2 and IL-4 (Hamann et al., 1997). Taken together it appears that the slow cycling CD84- population expressing CD45RA isoforms may be predom inantly responsible for immunological memory, but is less clear in case of the CD4-I- T cell subset, though. In addition it appears that the change in CD45 isoform expression after T cell stimulation is generally bi-directional and reversible.
Regulation of alternative splicing
The mechanism o f alternative splicing was investigated by transfecting several human CD45 mini gene constructs into mouse B and thymocyte cell lines. The results suggest that cells expressing the smallest CD45 isoform contain negatively trans-acting factors that allow the alternative exons to be skipped, but that the full length isoform containing all three alternative exons is the default pattern of splicing (Rothstein et ah, 1991). The same observation was made following transfection of a yeast artificial chromosome containing the entire human CD45 gene into a mouse B cell line, which expressed the low molecular weight isoform. The transfectants also expressed exclusively the low molecular weight isoform o f human CD45 (Fernandez Luna et al., 1991).
The opposite observation was made in earlier experiments by transfection o f total genomic DNA or a CD45 minigene containing the variably spliced exons into a fibroblast line. The removal o f the alternative exons by splicing between exon 3 and 7 suggests that the default pathw ay of splicing in these cells does not include exons A, B or C, because non- hematopoietic cells would not be expected to express the factors required for the regulation of CD45 alternative splicing (Raschke et al., 1985). The same postulate arises from later experiments using mouse CD45 mini gene constructs in fused T/B cells, which exhibited B- cell specific Ly-5 (mouse homologue of human CD45) transcripts (Saga et al., 1990). Also fusion of a mouse pre-B cell line expressing the high molecular weight isoform and a mouse T cell line expressing the low molecular weight isoform resulted in hybrids that expressed the high m olecular weight isoform, indicating that positive factors are required for the high molecular weight expression of CD45 (Saga et al., 1990).
Apart from the postulated transacting factors, sequences flanking the alternative exons as well as sequences within the exons are required for proper splicing. This was shown by the transfection of CD45 minigenes into hematopoietic cells. (Saga et al., 1990; Streuli and Saito,
1989). The elimination of almost all the intron sequences associated with exon 6 had no effect on the alternative splicing, while the full length exon 6 sequence was essential for alternative splicing (Tsai et al., 1989). However, alternative splicing of different CD45 isoforms is highly regulated after activation, and the conversion from CD45RA to CD45RO may be reversible rather than unidirectional and irreversible (Rothstein et al., 1991).
Stim ulation experim ents in culture suggest that transitions in CD45 isoform mRNA expression occur within the first 24 h after activation, and CD45RA persistence in T cells until day 4 resulting from slow turnover o f surface CD45RA glycoprotein (Deans et al.,
switches in alternative splicing, but also involves regulation of turnover of these isoforms from the cell membrane (Yamada et ah, 1992).
III. The process of pre-mRNA splicing
Interrupted genes and molecular splicing
The discovery of split genes 20 years ago has marked a critical step towards the understanding o f gene expression in eucaryotic organisms (Berget et ah, 1977; Chow et ah, 1977). In contrast to procaryotes it was found that m ost o f the protein-coding genes in higher eucaryotes are interrupted by multiple introns. As a result of this a significant proportion of the genome requires the accurate removal of intron sequences from the pre-mRNA by a process called RNA splicing. Therefore primary transcripts undergo an extensive series of processing reactions inside the nucleus before the mature RNA is exported into the cytoplasm.
As an indication of the extent of this accomplishment, one should be aware that specific splice sites can be separated by large intervening sequences varying in size from about 60 to more than ten thousand bases. The mechanism by which the exons are brought together and accurately ligated is achieved in a two step pathway as shown in early in in vitro and in vivo
experiments by a number of groups (Domdey et al., 1984; Padgett et al., 1984; Pikielny et al., 1983; Rodriguez et al., 1984; Ruskin et al., 1984; Zeitlin and Efstratiadis, 1984).
During the first step of this reaction the 5' site (donor) is cleaved, resulting in an exon with a 3' end and a lariat RNA in which the 5' end of the intron forms a phosphodiester bond with the 2 -OH o f an adenosine residue near the 3' splice site (acceptor). During the second step the intron is released as a lariat and the exons are ligated together. Experimentally, these two steps can be uncoupled by fractionation of the splicing components, by heat treatment of nuclear extracts from HeLa cells or by the use of mutant substrates (Reed and M aniatis,
1985). These genetic and biochemical methods have been used to address the question how this operation is accomplished.
The spliceosomal complex
chromatography (Bindereif and Green, 1986; Grabowski and Sharp, 1986; Pikielny and Rosbash, 1986)
Identification and characterisation com ponents o f the m am m alian spliceosom e is a prerequisite for understanding the mechanism of splicing control. In recent years the structure and function o f many components of the spliceosome have been determined. Five small nuclear ribonucleo-particles (snRNPs) and more than 50 additional proteins (W ill and Luhrmann, 1997), that range in molecular mass from smaller then 14 to 200 kDa (Reed,
1990), have been isolated. Some constituents were shown to be required for in vitro splicing. SnRNPs have been identified by com plem entation assays (Green, 1986; K rainer and Maniatis, 1985; Padgett et al., 1986), and they include the small ribonucleoprotein complexes (snRNPs) U l, U2, U4/6 and U5, and a large number of non-snRNP essential splicing factors (Krainer and Maniatis, 1985; Kramer, 1993; Lamm and Lamond, 1993; M aniatis and Reed,
1987; Moore and Sharp, 1993; Rio, 1993; Steitz, 1988).
The mechanism of splicing
The mechanism of the splicing reaction has been intensively investigated. The spliceosome is a highly dynamic complex, requiring ordered assembly on the pre-mRNA, rearrangement of its components and eventually dissociation from its substrates during the action of splicing (figure 1.3) (Green, 1986; Lamm and Lamond, 1993; Madhani and Guthrie, 1994; Padgett et al., 1986; Sharp, 1994; Stanley and Guthrie, 1998).
Figure 1.3; Mechanism of pre-mRNA splicing
ATP
U6
u:
U3 U6
® © ®
ATP?
a j5
ATP