Copyright 0 1997 by the Genetics Society of America
Identification of a New
Class
of Negartive Regulators Affecting
Sporulation-Specific Gene Expression in Yeast
Mei
Li
Benni* and Lenore Neigeborn*+**
Waksman Institute of Microbiology, +Department of Molecular Biology and Biochemistry and $The New Jersey Cancer Institute, Rutgers, the State University of New Jersey, Piscataway, New Jersey 08854-8020Manuscript received February 19, 1997 Accepted for publication July 21, 1997
ABSTRACT
We characterized two yeast loci, MDS3 and PMDl, that negatively regulate sporulation. Initiation of sporulation is mediated by the meiotic actintor IMEl, which relies on MCKl for maximal expression. We isolated the MDS3-1 allele (encoding a truncated form of Mds3p) as a suppressor that restores IMEl expression in mckl mutants. mds3 null mutations confer similar suppression phenotypes as MDS3-I,
indicating that Mds3p is a negative regulator of sporulation and the MDS3-1 allele confers a dominant- negative phenotype. PMDl is predicted to encode a protein sharing significant similarity with Mds3p.
mds3 pndl double mutants are better suppressors of mckl than is either single mutant, indicating that Mds3p and Pmdlp function synergistically. Northern blot analysis revealed that suppression is due to increased IMEl transcript accumulation. The roles of Mds3p and Pmdlp are not restricted to the MCKl
pathway because mds3 pmdl mutations also suppress IMEl expression defects associated with MCKl-
independent sporulation mutants. Furthermore, mds3 pmdl mutants express significant levels of IMEl
even in vegetative cells and this unscheduled expression results in premature sporulation. These pheno- types and interactions with , - RAS2-Val19 suggest that unscheduled derepression of IMEl is probably due - _ to a defect in recognition of nutritional status.
T
HE Saccharomyces cermisiae sporulation pathway con- sists of a complex series of events, the execution of which is cell-type dependent (restricted to a / a cells) and subject to specific environmental cues. The process initiates with premeiotic DNA synthesis followed by mei- osis, which is characterized by high levels of genetic recombination and reduction in chromosome content to generate four haploid nuclei. Continued progression through the developmental program is characterized by morphogeneses that result in packaging of the four meiotic products into prospores and enclosure of the prospores into a mature ascus resistant to a variety of harsh environmental conditions (ESPOSITO and KLAP-HOLTZ 1981; MITCHELL 1988; MALONE 1990; HONIG
BERG et al. 1993). Governing the execution of this path- way are a variety of regulatory mechanisms, including alterations in transcriptional gene expression patterns (CLANCY et al. 1983; HOLAWAY et al. 1987; SMITH and MITCHELL 1989; STRICH et al. 1989;
KIHARA
et al. 1991; VERSHON et al. 1992; BURNS et al. 1994; COE et al. 1994; MITCHELL 1994; BOWDISH, et al. 1995; HEPWORTH et al. 1995; FRIESEN et al. 1997), post-transcriptional control at the level of splicing and mRNA stability (ENGEBRECHT et al. 1991; MENEES et al. 1992; SUROSKY and ESPOSITO 1992; OGAWA et al. 1995) and the involvement of MAP kinase-mediated signal transmission (s) ( COSTIGAN andCmresponding author: Lenore Neigeborn, Waksman Institute of Mi-
crobiology, Rutgers, the State University of New Jersey, 190 Freling- huysen Rd., Piscataway, NJ 08854-8020.
E-mail [email protected]
Genetics 147: 1351-1366 (November, 1997)
SNYDER 1994; FRIESEN et al. 1994; KRISAK et al. 1994; HERSKOWITZ 1995). Thus, this model system represents an ideal tool for investigation of the control of develop mentally regulated cellular events.
A combination of two concurrent signals is required for yeast to initiate the sporulation program: establish- ment of the a / a cell type and nutrient deprivation (KAS SIR and SIMCHEN 1976; &NE et al. 1981; TATCHELL et al. 1985; MITCHELL and HERSKOWITZ 1986; GOUTTE and JOHNSON 1988; HERSKOWITZ 1988; DRANGINIS 1990;
MATSUURA et al. 1990), Both signals converge on stimu- lation of the meiotic-activator
Z M E l
(inducer of =io-sis) (KASSIR et al. 1988; SMITH and MITCHELL 1989; MAT-
SUURA et al. 1990; MITCHELL et al. 1990).
Z M E l
is essen- tial for sporulation-specific gene (SSG) expression and, hence, is required for all meiotic and subsequent sporu- lation events (KASSIR et al. 1988; SMITH and MITCHELL1989; MITCHELL et al. 1990; SMITH et al. 1990). Thus, control of
Z M E l
expression is a vital determining factor in triggering the sporulation pathway. Despite the criti- cal role played byZ M E l
in the control of meiosis and sporulation, little is known about the molecular mecha- nisms mediating the signals governing its expression. Indeed, several modulators ofZ M E l
expression have been identified genetically, but only one,R M E l ,
has been shown to act directly at theZ M E l
promoter (see below); the specific molecular roles of the others are still unclear.1352 M. Li Benni and L. Neigeborn
regulatory locus, MAT (KASSIR and SIMCHEN 1976; HER-
SKOWITZ 1988). Yeast heterozygous at the MAT locus (MATa/MATa diploids) express the negative regulator a l a 2 , which represses expression of haploid specific genes, among them being RMEl (regulator of meiosis) (MITCHELL and HERSKOWITZ 1986;
G o u m
and JOHN-SON 1988; DRANGINIS 1990). h e l p is expressed only in haploid cell types ( a and a cells) and functions di- rectly as a negative regulator of IMEl expression (COV-
ITZ et al. 1991; COVITZ and MITCHELL 1993). Hence, the a / a cell type is a positive regulator of sporulation because it leads to repression of a repressor of IMEI. The a / a cell type also activates I&%# expression and RES1 (-@ME1 escape) activity, which also promote the expression of IMEI, but in an RMEl-independent man- ner (SHAH and CLANCY 1992). From a biological per- spective, the a / a cell type is characteristic of the diploid phase of the yeast life cycle; thus this signal ensures that only diploid cells undergo meiosis, which would be lethal to haploid a and a cell types.
The response to nutrient deprivation represents a fundamental difference between the haploid and d i p loid cell types. Both reproduce vegetatively when nutri- ents are plentiful, however, nitrogen starvation, in the presence of nonfermentable carbon sources, leads to the cessation of cell growth and normal mitotic division (HERSKOWITZ 1988). Following starvation, a and a cells (haploids) enter a quiescent phase of the cell cycle considered to be analogous to Go (ESPOSITO and KIA-
PHOLTZ 1981). In contrast, a / a (diploid) cells activate IMEl expression and initiate the sporulation program (ESPOSITO and KLAPHOLTZ 1981; SMITH et aZ. 1990). The starvation signal triggering meiosis is partially trans- duced through decreased CAMP-dependent protein ki- nase (PJSA) activity (TATCHELL et aZ. 1985), which results in mitotic growth arrest that can lead to induction of IMEl expression (SMITH and MITCHELL 1989; MATSU- URA et al. 1990). However, this cannot be the only mech- anism, because CAMP levels alone do not appear to influence sporulation ability (MATSUMOTO et aZ. 1983; OLEMPSKA-BEER and FREESE 1987; CAMERON et aZ. 1988). The MCKl (meiosis and centromere regulatory
b-
nase) locus encodes a protein kinase that mediates the decision to initiate the sporulation program by func- tioning as a positive regulator of IMEl (NEIGEBORN and MITCHELL 1991). An mckl deficiency results in delayed and reduced levels of sporulation; excess MCKI gene dosage has a stimulatory effect. These phenotypes result as a direct consequence of altered gene expression of IMEI. MCKl is required for maximal IMEl transcript accumulation, as well as the expression of other SSGS.The mechanism controlling Mcklp activity has not been determined, however, it is clear that Mcklp does not contribute to cell type regulation. Its role in trans- mitting the starvation signal or another, as yet undeter- mined, signal remains to be elucidated. In addition, MCKl also promotes efficient ascus maturation and
centromere activity, but not via its role as an activator of IMEl expression (NEIGEBORN and MITCHELL 1991; SHERO and HIETER 1991; JIANG et al. 1995).
The MCKl gene encodes a protein kinase that is con- stitutively expressed (DAILEY et al. 1990; NEIGEBORN and MITCHELL 1991). In vitro, Mcklp can autophosphory- late at tyrosine and serine, but phosphorylates exoge- nous substrates at only serine and threonine (LIM et al.
1993; L. NEIGEBORN, unpublished results). Since it is unlikely that Mcklp controls IMEl expression directly, we suggest that Mcklp activates an IMEI-specific trans- activator, either directly or indirectly, via a phosphoryla- tion cascade. Identification of the factors functioning downstream of Mcklp is complicated by the fact that an mcklA deficiency produces only a partial defect in activation of IMEI. This observation suggests that there may be other pathways sharing functional specificity with Mcklp. Indeed, genetic studies reveal that the RTMI, 8, 9, 13 pathway is one example of such a redun- dant pathway (LI and MITCHELL 1997; Su and MITCH-
To identify IMEl regulatory components functioning downstream of or parallel to Mcklp we screened for dosagedependent suppressors of an m k l deficiency. We obtained a clone harboring an unusual dominant- negative allele of a previously uncharacterized locus that we denote M D S 3 ( Z c k l dosage suppressor). We show here that M D S 3 and its paralog PMDl (Earalog of mS3) represent synergistic negative regulatory activi- ties that operate downstream of both the mckl and riml
pathways to curb IMEl expression.
ELL 1993a,b).
MATERZALS AND METHODS
Strains, media, genetic methods and assays: All of the yeast strains used in this study are isogenic and derived from the SKI genetic background (ME and ROW 1974). Rele- vant genotypes and sources are shown in Table 1. The mcklA4::TWl, imelA13-HIS3, rmelAS::LEU2, and riml muta- tions, as well as the arg6 and met4 auxotrophic markers and the I"lacZ::URA? reporter alleles have been described pre- viously (SMITH and MITCHELL 1989; NEICEBORN and MITCH-
ELL 1991; su and MITCHELL 1993a,b).
Yeast strains were grown at 30". Complete (YEPD and YEPAc), synthetic (minimal and dropout), and sporulation
media were prepared as described by SHERMAN et al. (1986) ;
carbon sources, as indicated (glucose or potassium acetate), were added to a final concentration of 2%.
Standard genetic procedures were used for mating, diploid selection, sporulation, and tetrad analysis (SHERMAN et ul.
1986). Yeast transformations were performed by the lithium acetate procedure ( I T 0 et al. 1983). Sporulation efficiency was
assessed in quadruplicate from plate cultures unless otherwise indicated: strains were first grown overnight on YEPD plates and then replica plated to sporulation plates and allowed to incubate at 30" for 48 hr. Quantitation of total asci formed was determined by microscopic examination; no less than 200 cells were counted per sample. The values are the average of four independent assays for each strain.
Growth rates in YEPAc were monitored by growing each
Negative Regulators of Sporulation 1353
determining the ODm of the culture every 2 hr until station- ary phase was attained.
For P-galactosidase assays, cells were grown to early log phase in YEPAc medium (0 time), washed and shifted to an equal volume of liquid sporulation medium (2% KAc), and incubated for 6 hr. Samples for assay were harvested by cen- trifugation, washed in water, resuspended in Zbuffer, perme- abilized by addition of SDS and chloroform, and assayed as described (STERN et al. 1984). Values are the average of at least four independent assays.
Selection for multicopy suppressors of mcklA: LNY266 (mcklA4::TRPl, imelAl3-HIS3) was transformed with a YEp24 genomic library (CARLSON and BOTSTEIN 1982) and 2400 independent Ura+ transformants were picked and screened for acquistion of the ability to grow in the absence of histidine. Twelve transformants were found to express plas- middependent histidine prototrophy. The plasmids were re- covered via passage through Escherichia coli and their activity was verified by retransformation into LNY266 followed by test- ing for growth in the absence of histidine. A combination of restriction mapping and Southern analyses revealed the presence of 11 different plasmids: two of the plasmids con- tained the genomic HIS3 locus but none contained the geno- mic MCKl locus. Control experiments demonstrate that MCKl clones do confer the His+ phenotype in our screen, however, Southern analysis of library DNA revealed that the genomic MCKl locus is underrepresented as compared to other unique loci (data not shown). The remaining nine plas- mids define five different genomic regions (two regions were represented by overlapping clones). Given the relatively low number of transformants screened and the fact that several loci were recovered only once and MCKl was not recovered at all, we conclude that the screen was not saturated.
Plasmid constructions: pMCH8 (&tigopy suppressor conferring His+, no. 8) is the original clone from the YEp24 library (CARLSON and BOTSTEIN 1982) that confers suppres- sion of the mcklA defect in PIMEI-HIS3 expression; it contains an 8.5-kilobase (kb) genomic insert (see Figure 1A). The fol- lowing plasmids contain the indicated subcloned fragments derived from pMCH8: pMLl contains the 6 k b CluI-BumHI fragment cloned into the polylinker of pRS426 (SIKORSKI and HIETER 1989; CHRISTIANSON et ul. 1992); pML2: contains the 3.3-kb EcoRI fragment cloned into pRS426; pML3: contains the 6.1-kb N h I fragment cloned into the SpeI site of pRS426; pML5: contains a 1.4kb NheI fragment spanning the yeast DNA/vector junction cloned into the SpeI site of pRS426; pMLlO: contains the 3.4kb NcoI fragment, which was made blunt ended by filling in the 3’ recessed ends using Klenow enzyme and then cloned into the SmuI site of pRS426; pMLl1 contains a 2.9-kb LEUkontaining fragment cloned into the BgnI site of pMLl. pML51: contains a 4kb Hind111 fragment spanning the yeast DNA/vector junction cloned into pRS426. We used PCR to obtain a 2.3-kb fragment containing the amino terminus of the genomic MDS3 locus. The following primers were used with genomic DNA as template: 5’-CTG ACTTCCGATI’CCAACGAG3’ and 5”TATAGAGGACCTTA GAAG3’. This PCR product was a used as a probe to identify a genomic library clone carrying the amino terminus of MDS3 via colony hybridization in E. coli (NEIGEBORN and M ~ C H E L L 1991). We used this method, as opposed to directly obtaining the region via PCR, to avoid inadvertent PCR-mediated mum- tion of MDS3. One positive library clone, pML41, was retained and confirmed by restriction analysis to contain the amino terminus of MDs3 as well as several kilobases of upstream DNA. pML43 was constructed by cloning the 4.6kb Sua-NheI fragment from pML41 (including 1.9 kb of upstream DNA and the amino-terminal 2506 nucleotides of AfDS3) into pML10, thus creating a clone containing full-length MDS3
under &e control of its own promoter. The integrity of this clone was confirmed by sequence analysis.
The vector pADC416 contains the constitutive yeast A D c l promoter (including the transcriptional start site) cloned into the XhoI-Hind111 sites within the polylinker of pRs416 (SIKOR- SKI and HIETER 1989; CHRISTIANSON et al. 1992). This plasmid was constructed by subcloning the 1.42-kb %I-Hind111 frag- ment containing the ADCl promoter from pPC97 (CHEWY and NATHANS 1992) into the XhoI-Hind111 sites of pRs416. pML46 was constructed by cloning a PCRgenerated MDS3 fragment extending from the initiator ATG through the ter- mination codon (containing S m d sites at both ends) into the
S m d site of pADC416, thus creating an MDS3 allele driven by the ADCl promoter. To avoid PCR-mediated introduction of mutations into this construct, we then swapped everything from the CluI site (at nucleotide +1241 from the translational start) to beyond the end of the clone with the analogous non-PCRderived MDS3 DNA. pML47, pML48, pML49, and pML50 were constructed in the same way, except that each clone initiates from sequentially carboxy-terminal in-frame ATGs (see Figure 1B; pML48 and pML49 contain the same insert DNA, except in opposite orientations relative to the ADCl promoter). For pML48 and pML49, non-PCRderived wild-type DNA was swapped from the xbd site to the end of the clone; in pML50 non-PCRderived wild-type DNA was swapped from the Me1 site to the end of the clone. PCR- derived sequences retained on all of these clones were verified by sequence analysis not to contain any mutations. The follow- ing PCR primers were used in these constructions: 5‘46 5’- T A T T C C C G G G A T G C C T C A - 3 ’ ; 5’47: 5’-TAT
TCCCGGGATGCATGTAAAGAACGAMAT-3‘; 5’48: 5‘-ACC
CCCGGGATGTCTTCTATCCCT-3’; 5’50: 5”ACCCCC
GGGATGAGCATCCCGTCTGGT-3‘; and 3’46-50: 5”ACC CCCGGGGGTTTCATI’AGCCTITGGGAA-3’.
Construction of the mds32::LEU2 and mdr3P3::W al-
leles: A 2.9-kb genomic fragment including the LEU2 se- lectable marker was inserted at the BgllI site within the MLlS3- 1 coding region of pMLl to generate the allele mds3-2::LEU2 in plasmid pMLl1. A fragment containing the mds3-2::LEU2 allele was cleaved from the plasmid by digestion with NcoI and used to perform a one-step gene replacement (ROTH-
STEIN 1983) by transforming the diploid strain LNY357 and
selecting for leucine prototrophy. Integrity of the mutation was confirmed by Southern blot analysis.
To confirm that mds3-2::LEU2 confers the null phenotype, we also constructed the mds3A3::UM3 null allele that lacks most of the coding sequence. The 2.6kb CZuI-B@I fragment of pML43 was deleted and replaced by a 1.5kb ClaI-Bud1 fragment containing the cTRA3 gene to generate plasmid pML45 containing the mds3A3::UM3 allele. pML45 was then digested with Sac1 and PVuI to liberate linearized mds3A3::URA3, which was purified and used to perform a one-step gene replacement by transforming LNY357 to uracil prototrophy. The integrity of the mutation was confirmed by Southern blot analysis (data not shown). The phenotypes associated with this allele were indistinguishable from those associated with mds3-2::LEU2, thus, we conclude that mds3- 2::LEU2 confers a null phenotype (data not shown).
1354 M. Li Benni and L. Neigeborn
TABLE 1
Yeast strains
LNY266 LNY339
LNY342
LNY357
LNY746
LNY747
LNY748
LNY749
LNY763
LNY764
LNY765
LNY766
LNY767
LNY768
LNY807
LNY808
LNY809
LNY810
LNY890
LNY891
LM1897
LNY899
LNY1007
LNYl008
MATa arg6-2032S mlA5::LEU2 imelAl3-HIS3 mcklA4::TRPl MATa/MATa a@2032S/ARG6 met4-44S/MET4 imelA13-HIS3/IMEl
M A T d M A T a arg62032S/ARG6 met4-44S/MET4 imelAl3-HIS3/IMEl
MATa/MATa ar@2032S/ARG6 met444S/MET4 imelAl3-HIS3/IMEl
MATa/MATa arg6-2032S/ARG6 met4-44S/MET4 mds3-2::LEU2/MDS3
MATa/MATa arg62032S/ARG6 met4-44S/MET4 mds3-2::LEU2/MDS3
MATa/MATa arg6-2032S/ARG6 met4-44S/MET4 imelAl3-HIS3/IMEl
MATa/MATa arg6-2032S/ARG6 met4-44S/MET4 imelA13-HIS3/IMEl
MATa/MATa arg6-2032S/ARG6 met444S/MET4 imelAl3-HIS3/IMEl mcklA4::TRPl/MCKl mds4Al::LEU2/mds4Al::LEU2
mcklA4::TRPl/mklA4::TRPl mds4Al::LEU2/mds4Al::LEU2
mcklA4::TRpI/MCKl
imelAl3-HIS3/IMEl mcklA4::TRPl/MCKl
mcklA4::TRPl/mcklA4::TRPl imelAl3-HIS3/IMEl
m&3-2::LEU2/ mds3-2::LEU2 mckl A4::TWl/ MCKl
mcklA4::TRPl/mcklA4::TRPl mds3-2::LEU2/mds3-2::LEU2
mds3-2::LEU2/ mds3-2::LEU2 mds4A2::LEU2/mds4A2::LEU2
mcklA4::TRPI/mcklA4::TRPl
mds3-2::LEU2/ mds3-2::LEUL’ mds4A2::LEU2/ mds4A2::LEU2 mcklA4::TRPI/MCKl
rncklA4::TRPl/MCKl mds3-2::LEU2/MDS3 IME2-5-lacZ::URA3/
MATa/MATa arg62032S/ARG6 met4-44/MET4 imelAl3-HIS3/IMEl MATa/MATa ar&-2032S/ARG6 met4-44S/MET4 imelA13-HIS3/IMEl
MATa/MATa arg62032S/ARG6 met4-44S/MET4 imelAl3-HISjl/IMEl
IME2-5-lacZ::URA3
mcklA4::TRPI/mcklA4::TRPl IME2-5”lacZ::URA3/IME2-5-
lacZ::URA3 mds3-2::LEU2/MDS3
mds3-2::LEU2/ mds3-2::LEU2 IME2-5-lacZ::URA3/ IME2-5-lacZ:: URA3 mcklA4::TRPl/MCKl
MATa/MATa met4-44S/MET4 a@2032S/arg62032S imelA13-HIS3/ IMEl mcklA4::TRPI/mklA4::TRPl mds3-2::LEU2/mds3-2::LEU2 IME2-5-lacZ::URA3/IME2-5-lacZ::URA3
MATa/MATa arg62032S/ARG6 met4-44S/MET4 imelAl3-HIS3/IMEl
mds3-2::LEU2/mds3-2::LEU2 mcklA4::TRPI/mcklA4::TRPl r i m l A / rim1 A
MATa/MATcu arg62032S/ARG6 met4-44S/MET4 imelAl3-HIS3/IMEl mds3-2::LEU2/MDS3 mcklA4::TRP1/MCKl rimlA/rimlA
MATa/MATa ar&-2032S/ARG6 met444S/MET4 imelAl3-HIS3/IMEl mds3-2::LEU2/MDS3 r i m l A / r i m l A mcklA4::TRPl/mcklA4::TRPl MATa/MATa a@2032S/ARG6 met4-44S/MET4 imelAl3-HIS3/IMEl mcklA4::TR?’l/MCKl r i m l A / r i m l A mds3-2::LEU2/mds3-2::LEU2
MATa/MATa a@2032S/ARG6 met4-44S/MET4 imelAl3-HIS3/IMEl pmdlAl::LEU2/pmdlAl::LEU2 mds3-2::LEU2/mds3-2::LEU2
mcklA4::TRPI/MCKl
pmdlAl::LEU2/pmdlAl::LEU2 md.s3-2::LEU2/mds3-2::LEU2
mcklA4::TRPl/mcklA4::TRPl
MATa/MATa arg6-2032S/ARG6 met444S/MET4 imelA13-HIS3/IMEl mcklA4::TRPI/mcklA4::TRPl pmdlAI::LEU2/pmdlAl::LEU2 MATa/MATa ar&2032S/ARG6 met444S/MET4 imelAl3-HIS3/IMEl
pmdlAl::LEU2/pmdlAl::LEU2 rncklA4::TRPUMCKl
MATa/MATa arg62032S/ARG6 met444S/MET4 imelAl3-HIS3/lMEI m&3-2::LEU2/MDS3 pmdlAl::LEU2/PMDl mklA4::TRPl/MCKl r i m l A / R I M l
pmdlAI::LEU2/pmdlAl::LEU2 mds3-2::LEU2/MDS3
MATa/MATa arg6-2032S/ARG6 met444S/MET4 imelAl3-HIS3/IMEl
MATa/MATa arg6-2032S/ARG6 met444S/MET4 imelAl3-HIS3/IMEl
M A T a / / M A T a met4-44S/MET4 imelAl3-HIS3/IMEl rimlA/rimlA
This laboratory This laboratory
This laboratory
This laboratory
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
This study
Negative Regulators of Sporulation
TABLE 1
continued
1355
Strain
~~~ ~
Genotype Source
LNYl009
LNYlOlO
LNYlOll
LNYl102
LNYl104
LNYl105
LNYl106
LNYl119
LNYl121
MATa/MATa a@2032S/ARG6 &444S/MET4 imelA13-HIS3/IMEl
prndlAI::LEUZ/pmdlAl::LEU2 mklA4::TRPl/mcklA4::TRPl
r i m l A / r i m l A mds3-2::LEU2/MDS3
MATa/MATa imelAl3-HIS3/IMEl met444S/met444S rimlA/rimlA
pmdlAI::LEU2/pmdlAl::LEU2 mds3-2::LEU2/mds3-2::LEUZ
mcklA4::TRPl/MCKl
pmdlAl::LEU2/pmdlAl::LEU2 mds3-2::LEU2/mds3-2::LEU2 mklA4::TRPl/mcklA4::TRPl r i m l A / r i m l A
pmdlAl::LEU2/PMDl mds3A3::URA3/MDS3 mcklA4::TRPl/ MCKl
pmdlAl::LEUZ/PMDl mcklA4::TRPl/MCKl mds3A3::URA3/ mds3A3::URA3
pmdlAl::LEU2/pmdlAl::LEU2 mds3A3::URA3/MDS3 mcklA4::TRPl/MCKl
pmdlAl::LEU2/pmdlAl::LEUZ mds3Ajr::URA3/mds3A3::URA3
mcklA4::TRPl/MCKl
MATa/MATa ar@2032S/ARG6 met444S/MET4 mcklA4::TRPl/
MCKl imelAl3-HIS3/imelAl3-HIS3 pmdlAl::LEU2/
pmdlAl::LEU2 mds3A3::URA3/mds3A3::URA3
MATa/MATa a@2032S/ARG6 met444S/MET4 mklA4::TRPl/
MCKl imelAl3-HIS3/imelAl3-HIS3 pmdlAl::LEU2/PMDl
mds3A3::URA3/MDS3
MATa/MATa ar&Z032S/ARG6 met4-44S/MET4 imelAl3-HIS3/IMEl
MATa/MATa ar@2032S/ARG6 met4-44S/MET4 imelA13-HIS3/IMEl
MATa/MATa a@2032S/ARG6 met444S/MET4 imelAl3-HIS3/IMEl
MATa/MATa arg6-2032S/ARG6 met4-44S/MET4 imelAl3-HIS3/IMEl
MATa/MATa arg62032S/ARG6 met444S/MET4 imelAl3-HIS3/IMEl
This study
This study
This study
This study
This study
This study
This study
This study
This study
All yeast strains were derived from the SK1 background and harbor the following markers in addition to
~ ~~
those listed above; ura3SK1, h2-hisG, lys2SK1, ho::LYS2, tql-hisG, and his3.
PMDl locus via colony hybridization in E. coli (NEIGEBORN and MITCHELL 1991). We were unable to obtain a library clone carrying the PMDl locus in its entirety, therefore we used PCR with the following primers to obtain the carboxy terminal half of PMDl: 5’GCGTCAAGhlGGTCTTCTCA’IT- 3’ and 5’-TGCGCATGTAAC’ITCG’ITCA-3’. The resulting PCR product was cut with PVuII and Sac1 and cloned into pML28 generating pML29, which contains the PMDl locus reconstructed in its entirety. A 7.6kb ChI-EcoRI fragment from pML29 containing PMDI was cloned into pRS426 to create pML30. A 4.5-kb BgnI fragment within the PMDl cod- ing sequence was deleted from pML3O and replaced by the
LEU2 selectable marker to create the pmdlAl::LEU2 allele in pML31. Digestion of pML31 with SmaI and ApaI released a pmdlA1::LEUkontaining fragment that was used to create the chromosomal pmdlAl::LEU2 allele via the standard one- step gene disruption procedure. The integrity of this disrup tion was confirmed by Southern blot analysis (data not shown).
DNA sequencing, PCR reactions, and oligonucleotide syn-
thesis: Both single- and double-strand sequencing techniques were used. To sequence MDS3-1, Exonuclease 111-generated nested deletions were made from one end of pMLlO using the Erase-a-Base kit as recommended by the manufacturer (Promega). The deletion clones were then sequenced by the dideoxy method using the Sequenase Kit (Amersham) and the universal primer provided by the manufacturer. In addi- tion, single-strand pMLlO was sequenced using appropriate synthetic primers. All other clones containing PCRderived inserts were also sequenced to confirm the integrity of the
inserts. Computer analysis of the sequences was performed using the University of Wisconsin Genetics Computer Group nucleic acids sequence analysis programs (DEVEREUX et al.
1984; GENETICS COMPUTER GROUP 1991) and the Saccharo- myces Genome Database (CHERRY et al. 1996). The following PCR primers were used to obtain the 0.46kb DNA fragment derived from the genomic ACTl locus: ACT974 5’“rTCGA ACAAGAAATGGS’ and ACT1924: B’-AGMCACTTGTG GTG3’.
Northern blots and probes: Cells were cultured overnight to early log phase inYEPAc. For sporulation, cells were shifted from YEPAc when they were in the early log phase to an equal volume of Spo medium; samples were harvested at various time points for RNA isolation (SMITH and MITCHELL 1989). Standard methods were used for Northern blot preparation (SMITH and MITCHELL 1989). The following “P-labeled probes were made by Random-Prime Labeling (Boehringer Mannheim Biochemicals, Indianapolis, IN): a 0.6kb HindIII-
EcoRI fragment of IMEl (SMITH and MITCHELL 1989), a 0.4
kb BamHI-EcoRI fragment of IME2 (SMITH and MITCHELL 1989), a 1.0-kb BamHI-Sac1 fragment of HOP1 (HOLLINGS
WORTH et al. 1990), a 0.46kb fragment of ACTl corresponding
to bases 974-1434 of the coding region (CHERRY et al. 1996), and pC4 used as a loading control (LAW and SEAGALL 1988).
1356 M. Li Benni and L. Neigeborn
A
m c k l AS R X HXh RCBg NXh Nh BgR N B BXhHBgXhRBg H Nh SUppreSSiOn
Chrom I I I I I \ \ I / / I I I I I I I / I 1
VI1
///ps3y////////////+j
I
I I k b I e
pMCH8 / / / / / I / / / / / / / fl-b
+
pML3 v////////-
pML2
pMLl
+
-
-
p M L l l
-
pMLSI - / / / / / / / / / / / / / -
+
pMLlO pML5
+/-
-
pML43
-
FIGURE 1,"Delineation of the m c k l A suppressor locus (A4DS3). LNY266 was transformed with the indicated plasmids. The ability of a given plasmid to suppress mcklA was determined by scoring P,,v,;I-HIS3 expression by monitoring growth on SC-His- Ura medium.
+,
growth in 2 days; +/-, growth in 3 days; -, no growth. B, BnmHI; Bg, BglII; C, CluI; R, EcoRI; H, HindIII; N,NcoI; Nh, NheI; S, SalI; Xb, XbnI; X, XhoI. (A) The open bar is a restriction map of the genomic region surrounding MDSZ the il.IDS3 predicted ORF is indicated by the hatched box. The lower, thick horizontal bars indicate the pMCH8-derived yeast DNA insert carried on the indicated plasmid (hatched areas indicate the il.1DS3 coding regions predicted to be expressed); thin horizontal bars denote that the pMCH8-derived yeast DNA insert carried with it flanking vector sequences derived from the E p 2 4 tetracycline resistance locus (containing fortuitous promoter activity). The arrowhead on pMCHS indicates that the clone contains additional genomic sequences beyond those indicated. (B) Truncated forms of A4DS?were cloned behind the constitutive A D C l promoter (PADCI). The hatched boxes indicate the extent of m S 3 coding sequences present; Mds3p in-frame initiator ATGs are shown. The transcript arising from pML49 is derived from the noncoding strand and probably does not encode a meaningful translation product.
Printer. ZlllE2 quantitation was determined using the IPLab- Gel software; the pC4 signal served as a normalization control.
RESULTS
Identification of MDS3-1 as a dosage-dependent sup- pressor of mckl: To identify genetic loci functioning downstream of or parallel to Mcklp in the regulation of sporulation we exploited the role of MCKl in the activation of IMEl expression by using the reporter gene PTMEI-HIS3 (NEIGEBORN and MITCHELL 1991). The PIM;,-HIS3 allele consists of the HIS? coding sequence situated downstream of the IMEl promoter and mRNA start site. PIM;I-HIS3 strains deficient at the chromo- somal his? locus are histidine prototrophs only under conditions in which the IMEl promoter is active. Thus, wild-type strains are phenotypically His' while rncklA mutants are His- because the rnckl deficiency inhibits even basal levels of transcription driven by the IMEl promoter (NEIGEBORN and MITCHELL 1991).
We screened a yeast genomic multicopy library for sequences capable of restoring histidine prototrophy to
the rncklA PrMF;,-HIS3 strain LNY266 (NEIGEBORN and MITCHEL~L 1991). We reasoned that increased expres- sion of a substrate or target of Mcklp might be able to restore P,,,-HIS3 expression in an rncklA mutant; likewise, overexpression of positive regulators acting in MCKl-parallel pathways might also suppress an mcklA defect. Among the suppressor clones obtained in this screen was pMCH8, which contains the 8.5-kb yeast ge- nomic insert shown in Figure 1A.
Negative Regulators of Sporulation 1357
result strongly suggests that it contains at least part of the locus.
Sequence analysis of the 3.6-kb NcoI fragment re- vealed an apparent open reading frame (ORF) of 295 amino acids (see Figure 3) immediately upstream of GCNl on chromosome VI1 (MARTON et al. 1993). Com- parison of this region with that derived by the Saccharo- myces Genome Project revealed that this short reading frame actually represents an amino-terminal truncated portion of the larger ORF YGL197w (CHERRY et al. 1996). In other words, pMCH8 contains only the car- boxy-terminal portion of this locus, which we have named MDS3 (Eckl dosage Suppressor).
Surprisingly, the fragment of the gene retained on pMCH8 confers an mckl suppression phenotype despite the absence of -1 kb of coding sequence, the initiator methionine, and a promoter. Two possibilities can ex- plain this result: the activity responsible for mckl sup- pression is not due to expression of a gene product, but rather to increased dosage of the DNA itself (titration of a regulatory factor, for example) or a truncated version of the gene product is fortuitously expressed under the influence of a cryptic promoter, probably located in adjacent vector sequences. We favor the latter model for several reasons. The region conferring activity contains only protein coding sequence and it is unlikely (albeit, not unprecedented) that DNA control elements are present. More compelling, however, is the observation that a simple insertion at the unique BglrI site within the coding region is sufficient to eliminate suppressor activity even though large fragments spanning this site do not possess activity (Figure lA, compare pMLl and pML3 to pMLl1 and pML43). Finally, “flipping” the relative orientation of the truncated coding region abol- ishes suppressor activity (see Figure l B, compare PML48 and pML49). This is the expected result if a cryptic promoter were driving expression of the gene fragment. Indeed, the plasmid region adjacent to the yeast DNA insert contains the tetracycline resistance locus, which has been previously reported to contain nonspecific fortuitous promoter activity (STRUHL and
DAVIS 1981).
Figure 1A shows the restriction map of the full-length
MLX3 locus and indicates the “coding” region ex- pected to be expressed in pMCH8derived clones (as- suming that translation of the fortuitous transcript initi- ates at the first AUG, see below); we denote the allele present on pMCH8 as MDS3-1. To confirm the model that expression of m S 3 - 1 leads to mckl suppression and to determine whether this phenomenon is related to the function of the wild-type allele, we retrieved the full-length locus from a yeast genomic library by colony hybridization using a probe derived from pMCH8 (see M.4TERJAL.S AND METHODS). various constructs, using ei- ther the native MDS3 promoter (pML43) or the consti- tutive ADCl promoter (Figure l B ) , were tested for abil- ity to confer mckl suppression. Figure 1B summarizes
TABLE 2
enrpl.eseion suppresses the
4 1 A
sporulation defectSporulation efficiency
Clone Clone Clone Clone
Plasmid 1 2 3 4 Average
Vector 1.6 22 22 22 16.9
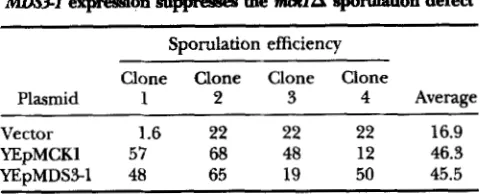
YEpMCKl 57 68 48 46.3 12 YEpMDS3-1 48 65 19 50 45.5
Strain LNY747 was transformed with YEp24 (vector), pAM301 (YEpMCKl), or pMCH8 (YEpMDS3-1) and four in- dependent transformants of each (clones 1-4) were trans- ferred to sporulation medium and ascus formation was quanti- tated after 48 hr.
our findings. Neither the full-length gene under the control of its own promoter (Figure lA, pML43) or the constitutive ADCl promoter (Figure IB, pML46) was capable of restoring Pzml-HIS3 expression to m k l A mutants. In contrast, a construct containing the same MDS3 coding information as that provided by MDS3-1 under the control of the ADCl promoter (pML48) is an efficient suppressor of mcklA, whereas the reverse orientation of this construct is not (pML49). Th’ IS ex- periment proves that MDS3-1 mediated suppression ac- tivity is, indeed, a consequence of expression of a trun- cated form of the MDS3 gene product. Furthermore, mckl suppression represents a novel, dominant phene type, because expression of the full-length gene does not behave in the same manner as MDs3-1 and MDS3-1 confers its phenotype in an MDS3 genetic background.
MDS3-1 expression suppresses the quautitative sporu-
lation defect associated with ntcR1A: To verify the bio- logical significance of the suppression activity conferred by MDS3-1, we tested the ability of this allele to suppress the sporulation defect conferred by mcklA. A diploid strain homozygous for the mcklA4::TRPl deficiency was transformed with plasmids harboring either m S 3 - 1 , MCKl, or vector alone and subjected to sporulation conditions followed by microscopic quantitation of sporulation efficiency. The results shown in Table
2
demonstrate that MDS3-1 restores sporulation levels to those comparable to that observed in the presence of MCKl (-46% as compared to 17% in the mcklA mu- tant carrying vector alone). It should be noted that the absolute sporulation level achieved in these strains is somewhat lower than those observed in strains carrying the genuine chromosomal MCKl allele. This is due to the inability to maintain plasmid selection under sporu- lation (nonvegetative) conditions. The result is an accu- mulation of a subpopulation of cells that have lost the plasmid and, therefore, lose complementation (or s u p pression) activity. Nevertheless, this experiment con-
1358 M. Li Renni and L. Neigeborn
Relevant
GenotvDe
PlMEI-HIS3
Exuression
Wild-type
mckl
mds3
mds3 mckl
+++
+++
++
SDorulation
75
%15
%75 %
40 %
FIGURE 2.-mds3::IJW2 suppresses the m c k l A sporulation defecn. Strains homozygous for the indicated mutations (wild tvpe, LNY746; mckl, LNY747; md.d, LNY748; mckl mds3, LW749) were assayed for their ability to express Pr,\ler-HI.Y3 and for their ability to sporulate. Pr,\II_,-Hl.S3 expression is shown in the photographic insert after 3 days incubation on
SCUra medium.
+++,
growth in 1 or 2 days;++,
growth in 3 days; -, n o growth. Sporulation data is an average of three independent trials for the each genotype; SD < 13%.MDS3 is a negative regulator of sporulation: To un- derstand the relationship between the roles played by the wild-type MIX3 and the MLX3-1 gene products, we created the chromosomal mds3”2::IJ<U2 allele. This al- lele represents an insertion of the I E U 2 selectable marker into the second BgAI site within the MDS3 cod- ing sequence. Although this did not necessarily repre- sent a null mutation, we chose this site for disruption because we had previously found it to be required for MDS3-I suppression activity (see Figure lA, pML11). Therefore, this mutation represent5 a perturbation of the activity defined by the MDS3-1 allele. Furthermore, subsequent experiments with the mds3A3::URA3 null allele (see MATERIALS AND METHODS) indicated that
mds3”2::IXU2 does indeed confer the null phenotype. Following replacement of the chromosomal MDS3locus with m(ts3-2::IEU2, standard strain construction meth- ods were used to generate the md~3-2::1fW2 strains listed in Table 1. Both haploid and diploid mds3 mu- tants are viable and display no prominent growth de- fects under vegetative conditions.
mds3”2::IEU2 strains were assayed for both Pl,,l,.~l-Hl.T3 expression and sporulation efficiency. If Mds3p is a pos- itive regulator of IMEI expression and sporulation (which dominant suppression of mckl suggests), a loss
of function allele should result in decreased Pl,~ll~l-Hl.T3 expression and sporulation efficiency. However, as the data provided in Figure 2 indicate, this is not the case. The results in Figure 2 demonstrate that the mds3- 2::IJW2 allele confers no observable reduction in the ability of Pl.,,l~l-HLS3 strains to grow in the absence of histidine. Furthermore, quantitation of sporulation ef- ficiency of mds3-2::IdXJ2 homozygous diploids also re- vealed no deficiency (Figure 2). Thus, the mds3”2::LEU2 mutation does not block entry into the sporulation pro- gram. There are several possible explanations to ac- count for this observation: mds3-2::LEU2 is not a null mutation; the role played by M D s 3 is only significant
in the absence of MCKI, or 1bfD.73 function is redun- dant. The possibility that rnds3-2::IXU2 is not a null mutation was ruled out by the fact that a deletion of MILT3 displays the same phenotype as mds3”2::IJZJ2 (data not shown).
To address the possibility that the phenotype con- ferred by mds3-2::141:‘U2 is masked in the presence of a functional MCKI allele (in this scenario, MDS3 would be a component of one of the Mcklpparallel path- ways), we analyzed the behavior of m c k l A mds3-2::1.1N2 double mutant strains (Figure 2, line 4). If the M I X 3 dependent pathway positively regulates IMI.‘l expres- sion and sporulation, we expect the double mutant to be even more defective in sporulation ability than the m c k l A single mutant. However, the result is exactly the opposite: rnrl.s3-2::IJW2 is a recessive supp-pssor of the mck1A defect. This observation is manifested both by restored l’l,~,l.~l-HlS3 expression and nearly threefold in- creased sporulation efficiency (Figure 2). Thus, bo!h the dominant MDS3-I and recessive rnd~3-2::IJ,‘U2 alleles confer the same phenotype: suppression of m c k l A . Since the suppression phenotype conferred by m.dy3- 2::IJW2 is recessive (data not shown), this must repre- sent the loss of function phenotype. Therefore, the wild-type role of 1 W S 3 must be negative regulation of IMEI expression and sporulation, and, MDS3-I repre- sents a dominant-negative allele.
m&3-2::LEU2 leads to increased expression of ZME2 To expand the biological relevance of MdsSp function, we analyzed the expression of the early SSG
IME2, whose expression is completely dependent on Imelp function (SMITH and MITCHELL 1989; YosHlnA
PI nl. 1990). We followed IME2 expression using the previously described IML2-5-lncZ::URA3 reporter gene (MITCHEI.I. P/ nl. 1990; SMITH P/ 01. 1990; SV and MITCH-
ELL 1993b) and assayed 0-galactosidase expression quantitatively (Table 3). Our results reveal that mds3- 2::IJXJ2 m c k l A double mutant strains display signifi- cantly higher levels of sporulation-specific IME2-lncZex- pression than that observed in mck1A strains; this result is in agreement with the l’l,,ll~l-HlS3 and sporulation data. In addition, this assay uncovered a modest, but consistently reprodltcible, increase in IME2”lncZexpres- sion in mds3-2::I.EU2 mutants as compared to wild type (Table 3, compare m d . ~ 3 and wild type). All these data support the model that MdsSp is a negative regulator of Imelpdependent SSG expression.
Suppression by mds3 is not restricted to Mcklpde-
M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 p P m d l p M d s 3 o
1 1
7 7
8 1
1 6 1 1 3 3 2 4 1
2 1 3 3 0 8 2 9 3 3 5 4 3 7 3 4 3 3 4 5 3 4 9 1 5 3 3 5 6 8 6 1 3 6 4 8 6 8 8 7 2 5 7 6 0 7 7 8 8 3 0 8 4 4
9 1 0 9 2 4 9 8 6
1 0 6 6 9 6 6 1 0 4 3 1 1 4 6 1 1 2 3 1 2 2 6 1 2 0 1 1 3 0 6 1 2 7 5 1 3 8 5 1 3 4 8 1 4 6 4 1 4 2 6 1 5 4 1
1 6 2 1
P m d l p 1 7 0 1
FIGURE 3.-Comparison of Mds3p and Pmdlp predicted amino acid sequences. Identical residues are highlighted in black boxes and conserved residues are highlighted in gray boxes.
*,
the initiator methionine (Mds3p amino acid 517) of the peptide corresponding to the m S 3 - I allele.of the lacZ gene product). Thus, loss of Mds3pmedi- ated repression results in increased ZMEl, ZME2, and SSG expression. We are unable to see this phenomenon
TABLE 3
mnk3-Z::LEUZ suppresses the m c k l A defect in IMEZ-lac2 expression
Relevant genotype IM2-lacZ expression Wild type
mck 1
mds3 mckl m k 3
90
150 60 6.1
Strains of the indicated genotype (wild type, LNY765; mckl, LNY766 m k 3 , LNY76f; and mckl md.73, LNY768) were assayed for sporulation-specific ,&galactosidase expression as de- scribed in MATERIALS AND METHODS. Values are expressed in Miller Units and are the average of at least three independent assays; SDs were < 15%.
when monitoring actual sporulation efficiency because
our wild-type strains sporulate with high efficiency. Thus, it is only manifest in strains compromised for sporulation induction, such as mcklA mutants. Accord- ingly, it is possible that mds3-2::ZXU2can suppress other mutations affecting ZMEI expression that define
Mcklpindependent pathways.
1360 M. Li Benni and L. Neigeborn
A
Wild Type mds3A mds3 mckl mcklA mds3 pmdl mcklHours in Spo
IME2
-
I M E l
+
HOPI +
-w
ACT1 +
Control+
B
Relative [ M E 2 60
Expression 5 0
E
-.
.o"
E
lower sporulation levels than that attributable to either mutation alone (Su and MITCHELL 1993b). Therefore, Mcklp and the Rimlp group are thought to function in separate pathways that share functional redundance
(SU and MITCHELL 1993b). A similar situation was re- vealed for the MDS4 locus (defining a third indepen- dent pathway), which was identified in the same screen as MDS3 (L. NEIGERORN, unpublished results).
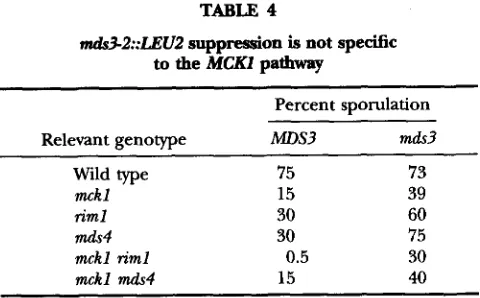
To determine whether mds3-2::LEU2 can also sup- press the I M I expression defects conferred by mds4A and r i m l a , we constructed the appropriate double and triple mutant strains. Table 4 summarizes our findings.
As expected, the r i m l A and m d ~ 4 A mutations confer reduced sporulation efficiencies: 30% as compared to
FIGL-RIC 4.-MD.Y3 and I'M111 negatively control early meiotic gene expres- sion. ( A ) Homozygous diploid strains of the in- dicated genotype (MT, LNY746; m h 3 , L.NY748; m 1 . d mrltl, LNY749; mrltl, L.NY747; rnh3 p m d l mrkl. LNY891) were grown to early log phase i n W A C medium (time 0) and then shifted to sporulation me- dium. At the indicated times, samples were re-
moved and total RNA was prepared, fraction- ated, and subjected to Northern analysis as de- scribed in MATICRIAIS ASD
M E T H O I X A single blot was probed for both IMEI and
IME2 simultaneously and then sequentially stripped and reprobed for HOPI,
A C T I , and the loading control probe pC4. ( R ) Quantitation of the data shown in A for IME2.
75% for the wild type. In contrast, both the m,d.s3 riml and mds3 mds4 double mutants demonstrated better sporulation efficiency: 60% and 75%, respectively. In- deed, the md.~3-2::IXU2 mutation could even relieve, to a certain extent, the requirement for both Mcklp and any one of these additional positive regulators ( m d ~ 3 mckl riml and mds3 mckl mds4 triple mutants sporulate much more efficiently than the mckl riml and mckl mds4 double mutants). These results indicate that m d ~ 3 - 2::ZEU2 bypasses the requirement for several indepen- dent positive regulators of I M H and sporulation, sug- gesting that MdsSp functions downstream or indepen- dently of all three of these pathways.
Negative
Regulators of Sporulation 1361
TABLE 4
nU"2::LEU2 suppression is not specific
to the MCKZ pathway
Percent sporulation
Relevant genotype m s 3 mds3
Wild type 75 73
mckl 15 39
riml 30 60
mds4 30 75
mckl riml 0.5 30
mckl mds4 15 40
Strains of the indicated genotype (wild type, LNY746; mckl,
LNY747; d 3 , LNY748; riml, LNY808; mds4, LNY339; mckl riml, LNY809; mckl mds4, LNY342; mckl mds3, LNY749; riml mds3, LNY810; mds4 mds3, LNY764; mckl riml mds3, LNY807; and mckl mds4 m h 3 , LNY763) were assayed for sporulation efficiency after 48 hr. Values are the average of at least three independent quantitations; SD were <15%.
to repress SSG expression: A search of the Saccharo- myces Genome Project database revealed that Mds3p shares a considerable degree of homology to the pre- dicted product of OW YER132c (CHERRY et al. 1996), an uncharacterized locus located on chromosome V
adjacent to GLC7 (FENG et al. 1991). We named this locus PMDl (Earalog of mS3). Figure 3 shows a com- parison of the predicted amino acid sequences of MDS3 and PMDl; they share 47% identity and 63% similarity over their entire lengths. Neither protein shows signifi- cant similarities to other proteins in the GenBank or SwissProt databases nor are any significant functionally identified motifs present.
Given the high degree of similarity shared by Mds3p and Pmdlp, it seemed reasonable that PMDl might also play a role in controlling IMEl expression. To gain a better understanding of how PMDl functions, we cre- ated the chromosomal pmdlAl::LEU2 mutation. As w a s the case for mds3 mutants, pmdl mutants are viable, express PIMEI-HIS3 normally, and sporulate as efficiently as wild type (Table 5). But, unlike mds3, @ d l was un- able to suppress the mckl or riml sporulation defects or restore P,MEI-HIS~ expression ( r i m l A data not shown). However, mutants containing both a pmdl and mds3 allele (pmdl mds3 mckl and pmdl mds3 riml triple mu- tants, for example) displayed even higher sporulation levels than that of their PMDl counterparts, indicating that Pmdlp does play a role in sporulation efficiency when observed under the appropriate conditions (com- pare Table 5, line 6 with line 8 and Table 4, line 5 with Table 5, line 9). Indeed, sporulation efficiency in the pmdl mds3 mckl riml quadruple mutant strain is re- stored to a level as high as that of the wild type (75%
sporulation, as compared to only 0.5% in the mckl riml double mutant and 30% in the mds3 mckl riml triple mutant). Similar results were also obtained for the com- binatorial effect of ma33 and pmdl on mds4 and mds4
TABLE 5
MDs3 and P m l function synergistically to regdate
sporulation efficiency
PImI-HIS3 Percent
Relevant genotype expression sporulation
Wild type
+++
75mckl
-
15mds3
+++
75p m d l
+++
75mds3 pmdl
+++
86mckl mds3
++
40mckl pmdl
-
20mckl mds3pmdl
+++
85riml mds3 pmdl ND 75
riml mckl mds3pndl ND 75
Homozygous diploid strains of the indicated genotype (wild type, LNY746; mckl, LNY747; mds3, LNY748; p n d l , LNY899;
mds3pmdl LNY890; mckl d 3 , LNY749; mckl W d l , LNY892 mckl m h 3 p m d l , LNY891; riml mds3 p d l , LNYl010; and
riml mckl mds3 p m d l , LNYlOll) were scored for P1m1-HIS3
expression by monitoring growth on SGHis-Ura medium. Sporulation efficiency was quantitated after 48 hr as described in MATERIALS AND METHODS. Values are the average of at least three independent quantitations; SD <15%.
+++,
growth in 1 or 2 days;+
+,
growth in 3 days; -, no growth.mckl mutants (data not shown). The observation that coincident mutation of both mds3 and pmdl leads to greater suppression of mckl, riml, and mds4 than that of either single mutation alone suggests that Mds3p and Pmdlp function synergistically to repress sporulation. Although the effects of mds3 and pmdl on sporulation efficiency and the expression of the PjmI-HIS3 and IME2-lacZ reporter alleles suggests that these regulators exert their effects at the level of transcript accumula- tion, we confirmed this by monitoring SSG expression via Northern analysis of sporulating cultures. Diploids were grown vegetatively to late log phase, shifted to sporulation medium (Spo) at time 0, and RNA was pre- pared after varying terms of incubation (Figure 4A). In the wild-type strain, IMEl transcript levels were low in vegetative cells and induced and peaked following 4 hr in Spo. IME2 transcript was undetectable in vegetative cells and appeared after 2 hr in Spo, reaching a peak at 4 hr. In the mckl mutant the patterns of both IMEl and IME2 transcript levels showed delayed kinetics and decreased overall accumulation, as previously observed
M. Li Benni and L. Neigeborn 1362
A
5
4
0
s3
r3 0
2
1
0
""---"---...""""".."""
.."--.."-..""".__.___.____
""""-.""..._._
.""".""...
.."""""".."
""""_""._."".""..
"".""...
0 5 10 15 20 25 30 35
Time
in
W A C
(hours)
B
Relevant
%
Sporulation
Genome
in Rich Medium
Wild Type
0.2
mds3
1.0
pmdl
0.2
mds3 pmdl
10.0
C
0 5 10 15 20 2 5 30 35
Time
in
YPA.c
(hours)
FIGURE 5.-mds3 pmdl double mutants are defective in sensing nutrient availability. (A) Homozygous diploids of the indicated genotype were grown to saturation in WD, diluted into WAC to an ODm = 0.05 (time = 0 ) , and monitored for growth over a 3@hr period. Wild type, LNYl102 0; imel,
LNYl1210; md.~3,LNYl104m;pmdl,LNYl105 +; mds3pmd1,
LNYl106 0 ; mds3 pmdl imel, LNYl119
..
(B) Homozygous diploids of the indicated genotype were grown to saturation in W D , diluted into WAC to an ODm = 0.05 (time = 0), and incubated for 24 hr. Ascus formation was monitored mi- croscopically. Wild type, LNYl102; mds3, LNYl104; pmdl,never observed in the wild type. Similar results were obtained for HOPI, another early SSG, whereas no al- teration was observed in actin gene expression, indicat- ing that the effect is specific for SSGs. Mutation of only p m d l had little-or no effect on SSG expression in either MCKl or mckl strains (data not shown). Figure 4B illus- trates the significance of the unscheduled IME2 expres- sion via quantitation of the IME2 data shown in Figure
4A. Here we see that mckl mutants harboring an mds3 mutation express increased levels of IME2 and this ex- pression is accelerated relative to wild type. Further- more, the combination of mds? and p m d l results in the appearance of significant levels of vegetative IME2 expression, an occurrence never oberserved in the wild type (similar results were observed for the HOP1 tran- scripts; see Figure 4A). Indeed, this result is likely to represent an undersestimate of vegetative early SSG ex- pression since mature asci were already visible at the time RNA samples were prepared, suggesting that early
SSG expression might already have been down-regu- lated (see Figure 5B and discussion below). Thus, in the absence of the Mds3p and Pmdlp gene products, unscheduled expression of early SSGs occurs. There- fore, Mds3p and Pmdlp are bona
$de
negative regula- tors of early SSG expression and we conclude that the increase in sporulation observed in mckl mds3 p m d l mutants is a consequence of removal of this negative regulation.mds3 pndl double mutants are defective in recogni-
tion of their nutritional status During the course of our analyses, we observed that mds3 pmdl double mu- tants displayed a marked growth defect, as compared to mds3, p m d l , or wild-type strains, under specific culture conditions. When grown in rich medium in the pres- ence of glucose as the sole carbon source, the wild-type, mds3, p m d l , and mds3 pmdl strains formed colonies on solid medium at comparable rates (data not shown). However, in rich medium containing a nonfermentable carbon source (acetate), the mds3pmdl double mutants displayed a dramatic growth defect (Figure 5A). This defect was observed in both haploid and a / a diploid mds3 p m d l strains (data shown for a / a diploids only). Microscopic inspection of these cultures revealed that the a / a diploid mds3pmdl strains had not only arrested growth, but a proportion had initiated the sporulation program and formed mature asci (Figure 5B). Wild type and mds3 or p m d l single mutants, in contrast, grew normally and did not sporulate significantly. Thus, the premature and unscheduled vegetative meiotic gene
LNYl105; mds3pmd1, LNYl106. ( C ) Homozygous diploids of the indicated genotypes and harboring plasmids containing either the wild-type RAS2 gene (open symbols) or the RAS2-
Vu119 allele (filled symbols) were grown to saturation under selective conditions and then diluted into WAC to a final
ODeo0 = 0.05. Growth was monitored over a 33hr period. Plasmid loss was <30% in all strains. Wild type, LNY746 0