Mutations that Affect the Efficiency of Translation of mRNA for the
cIZ Gene of Coliphage Lambda
Ed DUI, Michael
E.
Mahoney and Daniel
L.
Wulff
Department of Biological Sciences, State University of New York, Albany, New York 12222 Manuscript received January 20, 1986
Revised copy accepted December 15, 1986
ABSTRACT
Starting with the
XpRE-
strain Xctrl cy3008, which forms clear plaques, we have isolated two mutant strains, hdya2 ctrl cy3008 and Xdya3 ctrl cy3008, that form plaques with very slightly turbid centers. T h e dya2 and dya3 mutations lie in the region of overlap between the Pm promoter and the ribosome recognition region of the cZZ gene, and have nucleotide alterations at positions -1 and +5 ofp,,
and alterations in cZZ mRNA at -16 and -21 nucleotides before the initial AUG codon of the gene. Both mutations destabilize a stem structure that may be formed by cZZ mRNA, and dya2 also changes the sequence on cZZ mRNA that is complementary to the 3’-end of 16 S rRNA from 5’-UAAGGA-3’ to 5’-UGAGGA-3’.-The dya2 and dya3 mutations, along with the ctrl mutation, which destabilizes either of two alternate stem structures which may be formed by cZZ mRNA (these being more stable stem structures than the one affected by dya2 and dya3), were tested for their ability to reverse two ell-mutations that are characterized by inefficient translation of cZZ mRNA. These are cZZ3088, an A + G mutation four bases before the initial AUG codon, and clZ3059, a GUU + GAU (Val2 + Asp) second codon mutation. It was found that ctrl completely reverses the translation defects of these two mutations, while dya2 partially reverses these translation defects. T h e dya3 mutation has no effect on translation efficiency under any condition tested. However neither the ctrl mutation nor the dya2 mutation has much effect on translation efficiency in an otherwise cZZ+ background, indicating that other factors must limit the rate of translation of CZZ mRNA under these conditions.
ARIOUS mutations in the ribosome recognition
V
region and NHZ-terminal region of thecZZ
gene of bacteriophage lambda decrease the efficiency of translation of cZZ mRNA. These include cZZ3059, a GUU -+ GAU (Val2 -+ Asp) mutation in the second codon (PLACE et al. 1984); cZZ3086, an AUG + GUG mutation in the initiation codon (WULFF et al. 1984); and cZZ3088, an A + G mutation four bases on the 5’side of the AUG initiation codon (WULFF et al. 1984). T h e cause of the translation deficiency in XcZZ3059 is unknown, but XcZZ3086 is probably deficient because it creates an AUG codon two bases upstream from the proper start codon, and XcZI3088 may be deficient because it creates a UUG codon in the region between the Shine and Dalgarno sequence and the proper AUG initiation codon. In both cases the codons cre- ated by the mutations may interfere with initiation at the proper start codon. Alternatively, the translation deficiency in XcZZ3088 may result from a new stem structure which may be formed by cZZ3088 mRNA (WULFF et al. 1984).
Two mutations have been isolated that reverse the translation deficiency in XcZZ3059. One of these, c t r l , is a GCA + ACA (Ala4 -+ Thr) change in the fourth
cZZ
codon, and the second, ctr2, is a CGT + CGC change in the third cIZ codon (PLACE et al. 1984). Since ctr2 does not result in an amino acid change,Genetics 115 585-590 (April, 1987)
and since XcZZ3059 ctr2 makes a fully active
cZZ
protein, the cZZ- phenotype of XcZZ3059 can be explained entirely by the deficiency of translating cZZ mRNA. Both the c t r l and ctr2 mutations destabilize a stable stem structure that can be formed bycZZ
mRNA, and this destabilization was proposed to be the cause of the increased translation efficiency (PLACE et al. 1984). Although the c t r l mutation greatly increases effi- ciency of translation of cZZ mRNA in a cZZ3059 genetic background, it has little if any effect on translation efficiency in an otherwise cZZ+ strain (PLACE et al. 1984). In this paper we show that the c t r l mutation not only reverses the translation defect of XcZZ3059, but that it also reverses the translation defect in a second strain, XcZZ3088. We also report that dya2, a new mutation in the ribosome recognition region of the cZZ gene, partially reverses the translation defects of both XcZZ3059 and XcZZ3088, but has little effect in a cZZ+ genetic background. These results point to the existence of other factors which limit translation rates in cIZ+ genetic backgrounds. A second new mutation in the same region, dya3, was found to have no effect on translation rates under any conditions.MATERIALS AND METHODS
586
2
I
I
Is.0.I
FUET VAL A R G A A ASN Lrs AR A N GLU ALA 5'-C A A T T G T T A T C T A Z G A A AT A C T T A C A T A T G G T T C G T G C C G A G G C T-
3' 3'- G T TA A C A A T A G A T T C C T T T T A A T A T A T G A AT G T A T--'bA
AC C A A G C A C G T T T E T9
T TG C G T b T G C ' T C C G A-
5'A C
AAA
C G-10 region -35 region
A
PREFIGURE ].-DNA sequence of the pRE promoter and NH,-terminal region of the c l l gene (SCHWARZ et al. 1978; ROSENBERG el al. 1978; SCHMEISNER et al. 1980). The 6-base consensus sequences for the -10 and -35 regions of prokaryotic promoters are indicated between strands (ROSENBERG and COURT 1979; SIEBENLIST, SIMPSON and GILBERT 1980; HAWLEY and MCCLURE 1983). The TTGC sequences recognized by c l l protein are outlined. Transcription from pRE initiates at either of two sites, as indicated by the arrow (SCHMEISSNER et al. 1980). The c l l gene is transcribed from the p R promoter, which lies several hundred nucleotides to its left. The line labeled "S.D." indicates the SHINE and DALGARNO (1974) homology for the c l l gene. Hyphens have been omitted from the sequence for clarity. Relevant mutations are indicated. The ~113059, ctrl and ctr2 mutations are described in PLACE et al. ( I 984), the cl13088 and cy3008 mutations in WULFF et al. (1984), and the dya2 and dya3 mutations in this study.
work, and for determinations of plaque morphologies. For selection of various recombinants from phage crosses we used appropriate lysogenic derivatives of UC4 185 (BELFORT and WULFF 1971) and of N99 (ENQUIST and WEISBERC 1977). Strain UC6183 (F- galK- hfl recA- str- supo) (FIEN
et al. 1984) was used to construct lysogens for galactokinase assays.
Phage: The
pRE-
strain Xctrl cy3008 is described in PLACEet al. (1 984). The cII?088, cII?059, and ctrl mutations are described in WULFF et al. (1984) and PLACE et al. (1984). Other phage markers used in this study are described in WULFF (1976), WULFF et al. (1980), COURT et al. (1980), and FIEN et al. (1984).
General: Media, general phage techniques and phage crosses are described in WULFF (1976).
Construction of phage strains: T h e bacteriophage strains were constructed by suitable stepwise crosses from appro- priate starting strains by methods similar to those described by WULFF et al. (1980) and COURT et al. (1980). DNA
sequence determination was used to confirm genotypes where appropriate. Key crosses in the construction of var- ious single and double mutants are outlined below.
dya2: This mutation was separated from its original ge- netic background through the cross Ximm4'4 dya2 ctrl cy3008 Oam205 X Xcy3048, followed by selection for i ~ n m " ~ O+ plaques. A deeply turbid plaque of genotype dya2
was picked and the phage were crossed with hcIts857 cinl
Oam205, followed by selection for immA O+ plaques at 32".
A Cin+ plaque of genotype XcZts857 dya2 was picked. dya2 cZZ3059: The cross XcIts857 dya2 Oam205 X
cy3048 ~ 1 1 3 0 5 9 was followed by selection for imm' O+ plaques at 32". Clear (or almost clear) plaques were picked and tested f o r complementation with Xcy42 with a spot test.
A cy+ plaque o f genotype XcIts857 dya2 cIZ3059 was identi- fied.
dya3 cZZ3059: We attempted to construct hcIts857 dya?
by the same method as described above for XcIts857 dya2.
However, the strain proved to be of genotype XcIts857 dya? ctrl by DNA sequence determination. To obtain Xclts857 dya? 1913059, the cross Xclts857 dya? c t r l Oam205 X Ximm4j4 cy3048 clI3059 was followed by selection for immA O+ plaques at 32" and testing clear plaques for complementa- tion with Xcy42. A cy+ plaque of genotype Xclts857 dya? cI1?059 was identified.
cy3048 was followed by selection for imm' O+ plaques at dya3: The cross XcZts857 dya3 cII?059 Oam205 X
32". A deeply turbid plaque of genotype Xclts857 dya? was picked.
dya2 ctrl cy3008 Oam205
X hc113088 was followed by selection for imm434 O+ plaques. T h e desired Ximm4j4 dya2 ~113088 recombinants formed plaques with lightly turbid centers.
dya2 cl13088 was followed by selection for imm' 0' at 32".
A clear plaque of genotype Xclts857 dya3 ~ 1 1 3 0 8 8 was picked. cZZ3088 ctrl: We first constructed plasmids containing
DNA fragments from XcI1?088 and XcIts857 c t r l . Each plasmid contained a X DNA fragment, extending from a Hind111 site in the CI gene to the EcoRI site in the 0 gene (DANIELS et al. 1983), cloned into the Hind111 and EcoRl sites of a pBR322 derivative that lacked an NdeI site. We then used a unique NdeI site between the cII3088 and ctrl
mutations (DANIELS et al. 1983) and the Hind111 site to exchange the NdeI-Hind111 fragments of these two plasmids. E. coli C600 carrying a recombinant plasmid of genotype
~ 1 1 3 0 8 8 ctrl was infected with Xcl7, and Xcl7+ recombinants were selected by plating on the h f A l strain UC6183. A
recombinant of genotype XcI13088 ctrl was identified by
DNA sequence determination. This strain also had an uni- dentified clear plaque mutation, which was removed by crossing it with Aimm4j4 cII3088 Oam205, and picking a turbid plaque of genotype Ximm4j4 ~113088 c t r l .
Construction of lysogenic bacteria: A set of lysogenic UC6 183 strains was constructed and tested as described in FIEN et al. (1984). T h e starting strains were either in a genetic background of hint6 red? clts857 Pam? (for dya2, dya?, c t r l , c11?088 c t r l , and ~ 1 1 3 0 5 9 c t r l ) , or hint6 cIts857 cro27 Pam? (for ~ 1 1 3 0 8 8 , ~ 1 1 3 0 5 9 , dya2 ~113088, dya2 ~ 1 1 3 0 5 9 , dya? cZ13088, and dya3 ~ 1 1 3 0 5 9 ) . In addition to the tests described in FIEN et al. (1984), immh P+ recombinants were obtained from the lysogenic bacteria by infection with
c l - and incubation until lysis, and were tested for the presence of the desired dya, ctr and cII mutations by
DNA sequence analysis.
Mutagenesis: Xctrl cy3008 was passed through an E. coli mutD mutator strain (FOWLER, DECNEN and Cox 1974) by the method of ENQUIST and WEISBERG (1 977).
DNA sequence determinations: DNA sequences of Sam7
derivatives of phage strains were determined according to the method of MAXAM and GILBERT (1 980) as described previously (WULFF et al. 1980, 1984).
Plasmids: The plasmid pKM2, in which the X
pRE
pro- dya2 cZZ3088 T h e crossLambda mRNA Translation 587
moter governs expression of the E. coli galK gene, is de- scribed in FIEN et al. ( 1 984).
Galactokinase measurements: Galactokinase was assayed at 30 min after shifting a lysogenic UC6183[pKM2] deriv- ative from 32", a temperature at which cZl gene expression is repressed, to 42 O , as described by FIEN et al. ( 1 984).
RESULTS
The dyat and dya3 mutations: T h e clear plaque
pRE-
strain Xctrl cy3008 (PLACE et al. (1984) (Figure1) was mutagenized by passage through an E. coli mutD mutator strain, and the strains Xdya2 c t r l cy3008 and Xdya3 c t r l cy3008 were isolated as variants that form plaques with very lightly turbid centers. T h e plates must usually be incubated
20-24
hr for the turbidity to become detectable, and even then turbid- ity can be seen only under optimal lighting conditions. T h e turbidity is so light that these two strains would ordinarily be classified as clear plaque mutants. In fact the strain Xcy3008, which is standardly classified as a clear plaque strain, forms plaques with very light turbidity, indistinguishable from those of Xdya2 c t r l cy3008 and Xdya3 c t r l cy3008, and a third mutant strain with very lightly turbid plaques that was isolated after mutagenesis of Xctrl cy3008 proved to be of genotype Xcy3008 according to DNA sequence analy- sis.Sequence changes of dyu2 and dyu3 mutations:
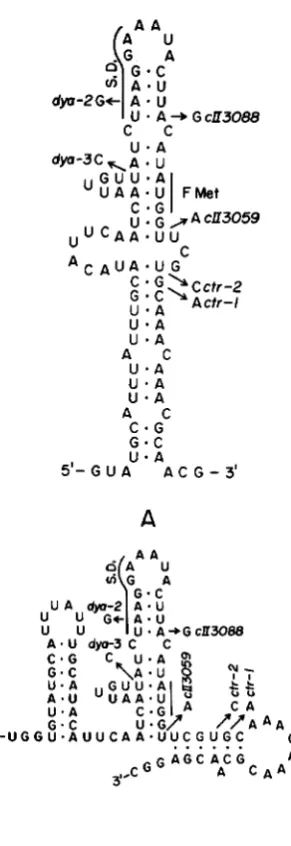
DNA sequence determination of the dya mutant strains revealed, in addition to the original c t r l and cy3008 alterations, single base changes in the ribosome recognition region of the cll gene. T h e d y a 2 mutation is an A + G change on c l l mRNA 16 bases 5' to the initial AUG codon, and the d y a 3 mutation is a U + C change 21 bases 5' to the initial AUG codon (Figure
Both the d y a 2 and d y a 3 mutations decrease the stability of a potential stem and loop structure that may be formed by cll mRNA (Figure
2).
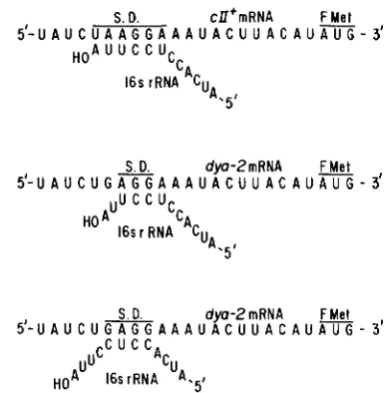
In addition, the d y a 2 mutation creates a poorer SHINE and DAL-GARNO (1974) sequence (the region of c l l mRNA that is complementary to the 3'-end of 16 S rRNA), al- though it preserves the 5'-AGGA-3' portion of the sequence that is the critical element for ribosome recognition (STORMO, SCHNEIDER and GOLD 1982). T h e d y a 2 mutation also results in the sequence 5'- GAGG-3', which constitutes an alternate pairing mode with the 3'-end of rRNA (Figure 3). One would not expect the d y a 2 ribosome recognition sequence to be appreciably less effective than that of ell+, and it could be more effective because of the two possible pairings.
T h e X c l l protein binds to a specific recognition site on the X
pRE
promoter to activate transcription frompRE
(SCHMEISSNER et al. 1980; SHIMATAKE and ROSEN-BERG 1981; Ho, WULFF and ROSENBERG 1983). T h e
p,,
promoter overlaps with the ribosome recognition 1).A
C . A
U A . C ' U * U
* A + G
C ' c l l 3 0 8 8
U . A
A C
U . A U * A U . A
A C
C - G G - C U S A 5 ' - G U A A C G - 3 '
A
A A L j f A U d \ G h
u u U * A +G CII3OBB A . U dyad C C
C * G C U * A cn G - C \ A - U
3
Y i 'L +
U . A
A - U U . A
u8ii:i1
C A G * C0 0
U . G ~ P P A A A 5'-U G G U * A U U C A A . U U C G U . . . . _ _ . . . G C C
A G C A C G A
3" C G G A C A A
B
FIGURE 2.-Two alternate stem and loop structures that can be formed by c l l mRNA in the NHn-terminal reigon of the c l l gene. The SHINE and DALGARNO sequence and the initial cII codon are indicated. The sequence changes of mutations used in this study are indicated. The stem structures affected by the dyu2 and dyu3
mutations (the upper stem of structure A and the middle stem of structure B) are identical in the two formulations, while the stem structures affected by the ctrl and ctr2 mutations (the lower stem of structure A and the right-hand stem of structure B) are different and mutually exclusive. Structure A is identical to the structure proposed in Figure 5 of PLACE et al. (1984), but with some weak additional features. Structure B was suggested by Dr. G. STORMO.
region and NH2-terminal end of the cll gene (Figure l), and mutations in the region of overlap can affect
pRE
promoter strength, cll protein activity, or both. (The cll gene is transcribed in the opposite direction from P E E , from a promoter which lies well outside theregion of overlap.) T h e d y a 2 and d y a 3 mutations lie near the start site of transcription for the P R E pro-
S. D. cU'mRNA F M e t 5'-U A U c U A A G G A A A U A c U U A c A U AUG
- 3'
HOA U U C C U
c,
UA 16srRNA CU
A.5'
TABLE 1
Effects of various mutations on cZZ activity
Relative Relative Geno- Relative Genotype activity Genotype activity type activity
S. D. d y a - P m R N A F M e t
5'-U A U C U G =A A A U A C U U A C A U
-
3''5'
S. D. d y a - 2 m R N A F M e t 5'-U A U C U - A A A U A C U U A C A U A m
-
3'CU uCc ' A
HOA" 16srRNA
FIGURE 3.-Possible pairings between the NH~terminal region of c l l mRNA and the 3'-end of 16 S rRNA (SHINE and DALGARNO
1974) for ell+ mRNA and dya2 mRNA.
change at +5 (Figure 1). In this region of prokaryotic promoters, DNA sequence does not have a strong influence on promoter activity, and no mutations which influence promoter activity have been reported near promoter start sites in any prokaryotic system. On the other hand, compilation of promoter DNA sequences reveals a nonrandom distribution at certain positions near the start sites (HAWLEY and MCCLURE
1983), and RNA polymerase certainly "covers" the DNA at the locations of the dya2 and dya3 mutations. In the following paper (MAHONEY and WULFF 1987) we show that both mutations result in increased p R E
activity.
Effects of the dyu2 and dya mutations on transla- tion efficiency: Since the ctrl mutation reverses the translation defect in XcZ13059 but has little effect on the efficiency of translation of the c l l gene in an otherwise wild-type genetic background (PLACE et al. 1984), we reasoned that any effects of the dya2 and dya3 mutations on translation efficiencies might be most apparent in a strain containing the dl3059 mu- tation, or some other c l l mutation which results in a translation deficiency, such as ~113088. We therefore constructed strains containing either the cZZ3059 mu- tation or the ~113088 mutation and one of the three mutations c t r l , dya2 o r dya3, in all possible pairwise combinations. We also constructed strains with dya2
and dya3 alone. These were then incorporated into
single copy prophages in order to test the amount of cll-activated transcription from a multicopy plasmid in which the X
pRE
promoter governs expression of the E. coli galK gene (FIEN et al. 1984).T h e results are shown in Table 1. A cl13059 muta- tion in the prophage results in only 1.4% of the galactokinase activity generated by a c l l + prophage.
However this does represent low level cll-dependent transcription from
pRE:
less than 0.5% of the cZZ+cI13059 1.4 ~113088 14 C I S 100
~113059 dya3 1.6 ~113088 dya3 16 dya3 126 cl13059 dya2 8 ~113088 dya2 43 dya2 150 ~113059 ctrl 120 cl13088 ctrl 200 ctrI 160
An appropriate lysogenic derivative of UC6183 carrying pKM2, a multicopy plasmid in which the Xp, promoter governs expression of the E. coli galK gene (FIEN et al. 1984), was grown to log phase at 32" and then shifted to 42" to induce c l l gene expression. At 30
min extracts were made and galactokinase activities determined (FIEN et al. 1984). For determination of high activities (e.g., more than 50% of the ell' activity), the lysogenic strains were of genotype
UC6183 (Xint6 red3 clts857 Pam3) and carried, in addition, the indicated mutations on their prophages. For determination of low activities (e.g., less than 50% of the c l l + activity), the lysogenic strains also had cro27 mutations in order to increase the sensitivity of the assay system. All the observed activities were normalized to
UC6183(Xint6 red3 clts857Pam3) equal to 100 for cro' strains, and to UC6183(Xint6 clts857 cro27 P a d ) equal to 100 for cro27 strains. The observed galactokinase activities were 6 6 units for the cro+
strain and 517 units for the cro27 strain. The value observed for
~113059 is about four to five times higher than the value observed for a corresponding ~113067 strain. [The ~113067 mutation is an AUG + ACG initiation codon mutation; WULFF et al. (1984).]
activity is seen with a prophage containing the initia- tion codon mutation ~113067 (AUG .--) ACG) (MAHO-
NEY and WULFF 1986). In a cl13059 genetic back- ground, the ctrl mutation results in nearly a 100-fold increase in cll-dependent transcription, to yield ga- lactokinase levels close to those observed with ell+.
T h e dya2 mutation results in a much more modest increase in cll-dependent transcription in a cl13059 background, namely about sixfold to 8% of the cZl+ level. T h e dya3 mutation has no measurable effect on the level of transcription associated with the cl13059 mutation.
A cl13088 mutation in the prophage results in about a sevenfold decrease from the cZZ+ level, which is less extreme than the decrease caused by the cZZ3059 mutation. T h e c t r l mutation completely reverses the cl13088 effect, while the dya2 mutation in a cl13088 genetic background results in a more modest three- fold increase in galactokinase activity, to 43% of the cZl+ level. T h e dya3 mutation has no measurable effect on c l l expression in a ~113088 genetic background.
However, in the absence of a mutation reducing translation efficiency (e.g., in an otherwise ell+ back- ground), the ctrl and dya2 mutations have much smaller effects, amounting to about a 50% increase over the galactokinase activity with a c l l + prophage.
Lambda mRNA Translation 589
are slightly less turbid than those of A+; while plaques of Xctrl, XcZZ3088 c t r l , and Xdya2 have fully turbid centers which are indistinguishable from those of A+.
All dya3 derivatives form plaques which are indistin- guishable from their dya3+ counterparts.
DISCUSSION
Secondary structure and the efficiency of trans- lation of cIZ mRNA: T h e dya2 mutation partially reverses the translation deficiency associated with either the cZZ3088 or the cZZ3059 mutations, while the dya3 mutation has no effect in either a cZZ3088 or cZZ3059 genetic background. T h e ctrl mutation com- pletely reverses the translation deficiency associated with either the cZZ3088 or the cZZ3059 mutations, and the ctr2 mutation was previously shown to reverse completely the translation deficiency associated with the cZZ3059 mutation (PLACE et al. 1984). (The ctr2 mutation has not been tested in combination with cZZ3088.)
T h e four mutations dya2, dya3, ctrl and ctr2 all destabilize potential stem structures in the ribosome recognition region and NH2-terminal end of cZZ mRNA (Figure
2 ) .
This destabilization has been pro- posed to be the cause of the increased translation efficiencies associated with the c t r l and ctr2 mutations (PLACE et al. 1984). T h e potential stem structure which is affected by the dya2 and dya3 mutations is less stable than the structures affected by the ctrl and ctr2 mutations [using the simple rules of TINOCO et al.(1 973) to calculate stabilities], a statement which is valid for either of the two alternate stem structures shown in Figure
2.
In fact it is by no means clear that the potential structure affected by the dya2 and dya3 mutations would form in vivo even in a wild-type genetic background. T h e observations that the ctr I and ctr2 mutations strongly reverse translation defi- ciencies, while the two dya mutations have little (dya2) or no (dya3) effects in translation deficient back- grounds, are consonant with the predicted relative strengths of these stem structures.T h e translation deficiency in XcZZ3088 could be caused by the creation of a UUG codon which inter- feres with recognition of the proper AUG start codon, or, alternatively, by the formation of an additional stem structure, consisting of hydrogen bonding be- tween the sequence immediately preceding the AUG initiation codon, 5’-UUGCAU-3’, and a sequence positioned 9 nucleotides downstream, 5’-GUGCAA- 3’ (WULFF et al. 1984). T h e cZZ3088 A + G change is central to the formation of this stem structure, which is mutually exclusive with either of the two structures shown in Figure
2 .
T h e ctrl mutation, in addition to destabilizing the stem structures shown in Figure 2,also destabilizes the potential stem structure associated with the cZZ3088 mutation (the complementary se-
quence is altered from 5’-GUGCAA-3’ to 5’-GUA- CAA-3’). T h e full reversal of the cZZ3088 translation deficiency in XcZZ3088 ctrl may result from the de- stabilization of both stem structures by the ctrl mu- tation.
T h e dya2 and dya3 mutations should each have strongly destabilizing effects on the weak stem struc- ture they affect, but in actuality the dya2 mutation, by disrupting a stretch of four consecutive base pairs, may have a greater destabilizing effect on secondary structure than the dya3 mutation, which leaves these four base pairs intact. This could explain why the dya2 mutation, but not the dya3 mutation partially reverses translation deficiencies.
The SHINE and DALCARNO sequence and the ef- ficiency in translation of cZZ mRNA: T h e reversal of translation deficiencies by the dya2 mutation may be unrelated to its possible effects on mRNA secondary structure, but be caused by its alteration in the SHINE and DALCARNO homology instead. At first inspection one would predict that the dya2 mutation, by decreas- ing the length of this homology from six bases to four, should have the opposite phenotype from that actually observed. But the critical 5’-AGGA-3’ sequence is left intact (STORMO, SCHNEIDER, and GOLD 1982), and an alternate pairing mode with a 5’-GAGG-3’ sequence is introduced by the mutation (Figure 3). This could improve translation efficiencies either by increasing the probability of initial interaction of ribosomes with the translation initiation region, or by providing an alternate alignment of mRNA on the ribosome. We conclude that the effect of dya2 on translation could result from either its effect on mRNA secondary structure or from the alteration in the SHINE and DALCARNO sequence.
Factors controlling translation efficiency: In PLACE et al. (1984) we predicted that ctrl would compensate for other translation deficiencies besides that associated with the cZZ3059 mutation. In this paper we show that it does indeed fully reverse the translation deficiency associated with the cZZ3088 mu- tation. Similarly, the dya2 mutation described in this work is generally effective in increasing translation efficiencies in a genetic background associated with a translation deficiency. But like c t r l , the dya2 mutation has little effect in a translation proficient (cZZ+) back- ground, and, under these conditions, other factors apparently limit the extent of cZZ protein production. A similar situation was found by MUNSON et al. (1 984) for ZacZ translation initiation mutations, in that mu- tations changing the length of the SHINE and DAL- CARNO sequence had much smaller effects on transla- tion rates under translation proficient conditions (re- duced secondary structure) than under translation deficient conditions (increased secondary structure).
secondary structure for cI2 mRNA. We thank G. PINCHBECK for technical assistance and K. SCHUFF for typing the manuscript. This research was supported by grant GM28370 from the National Institutes of Health.
L I T E R A T U R E C I T E D
BELFORT, M. and D. L. WULFF, 1971 A mutant ofEscherichia coli
that is lysogenized with high frequency. pp. 739-742. In: The Bacteriophage Lambda, Edited by A. D. HERSHEY. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York. CAMPBELL, A., 1961 Sensitive mutants of bacteriophage A. Virol- COURT, D., C. BRADY, M. ROSENBERG, D. L. WULFF, M. BEHER, M.
MAHONEY and S. IZUMI, 1980 Control of transcription ter- mination: a rho-dependent termination site in bacteriophage lambda. J. Mol. Biol. 138: 231-254.
DANIELS, D. L., J. L. SCHROEDER, W. SZYBALSKI, F. SANGER, A. R. COULSON, G. F. HONG, D. F. HILL, G. B. PETERSON and F. R. BLATTNER, 1983 Complete annotated lambda sequence. pp. 519-676. In: Lambda II, Edited by J. HENDRIX, J. ROBERTS, F.
STAHL and R. WEISBERG. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
A genetic analysis of the att-int-xis region of coliphage lambda. J. Mol. Biol. 111:
97-120.
FIEN, K., A. TURCK, I. KANG, S. KIELTY, D. L. WULFF, K. MC- KENNEY and M. ROSENBERG, 1984 CII-dependent activation of the pnE promoter of coliphage lambda fused to the Esche- richia coligalK gene. Gene 32: 141-150.
Mutational specificity of a conditional Escherichia coli mutator, mutD5. Mol. Gen. Genet. 133: 179-191.
Compilation and analysis of Escherichia coli promoter DNA sequences. Nucleic Acids Res. 11: 2237-2255.
Bacteriophage X protein cII binds promoters on the opposite face of the DNA helix from RNA polymerase. Nature 304: 703-708.
Mutations that improve the pnE promoter of coliphage lambda. Genetics 115: 591-595.
Sequencing end-labelled DNA with base-specific chemical cleavages. Methods Enzymol. 65:
499-559.
MUNSON, L. M., G. D. STORMO, R. L. NIECE and W. S. REZNIKOFF, 1984 lacA translation initiation mutations. J. Mol. Biol. 77:
663-683. ogy 14: 22-32.
ENQUIST, L. W. and R. A. WEISBERG, 1977
FOWLER, R., G. DEGNEN and E. Cox, 1974
HAWLEY, D. K. and W. R. MCCLURE, 1983
Ho, Y. S., D. L. WULFF and M. ROSENBERG, 1983
MAHONEY, M. E. and D. L. WULFF, 1987 MAXAM, A. and W. GILBERT, 1980
PLACE, N., K. FIEN, M. E. MAHONEY, D. L. WULFF, Y. S. Ho, C. DEBOUCK, M. ROSENBERG, M.-C. SHIH and G. N. GUSSIN, 1984 Mutations that alter the DNA binding site for the bacteriophage XcII protein and affect the translation efficiency of the cII gene. J. Mol. Biol. 180: 865-880.
Regulatory sequences in- volved in the promotion and termination of RNA transcription. Annu. Rev. Genet. 13: 319-353.
ROSENBERG, M., D. COURT, H. SHIMATAKE, C. BRADY and D. L. WULFF, 1978 T h e relationship between function and DNA sequence in an intercistronic regulatory region in phage lambda. Nature 272: 414-423.
SCHMEISSNER, U., D. COURT, H. SHIMATAKE and M. ROSENBERG, 1980 Promoter for the establishment of repressor synthesis in bacteriophage A. Proc. Natl. Acad. Sci. USA 77: 3 191-3 195. SCHWARZ, E., G. SCHERER, G. HOBOM and H . KOSSEL,
1978 Nucleotide sequence of CTO, cII and part of the 0 gene in phage X DNA. Nature 272: 410-414.
Purified X regulatory protein cI1 positively activates promoters for lysogenic devel- opment. Nature 292: 128-132.
SHINE, J. and L. DALGARNO, 1974 T h e 3’-terminal sequence of Escherichia coli 16s ribosomal RNA: complementarity to non- sense triplets and ribosome binding sites. Proc. Natl. Acad. Sci. USA 71: 1342-1346.
SIEBENLIST, U., R. B. SIMPSON and W. GILBERT, 1980 E. coli RNA polymerase interacts homologously with two different pro- moters. Cell 2 0 269-28 1 .
STORMO, G. D., T. D. SCHNEIDER and L. M. GOLD, 1982 Characterization of translational initiation sites in E. coli. Nu- cleic Acids Res. 10: 2971-2996.
TINOCO, I., P. N. BORER, B. DENGLER, M. D. LEVINE, 0. C. UHLENBECK, D. M. CROTHERS and J. GRALLA, 1973 Improved estimation of secondary structure in ribonucleic acids. Nature New Biol. 246: 40-41.
Lambda cin-1, a new mutation which en- hances lysogenization by bacteriophage lambda, and the ge- netic structure of the lambda cy region. Genetics 82: 401-416. WULFF, D. L., M. BEHER, 2. IZUMI. J. BECK, M. MAHONEY, H.
SHIMATAKE, C. BRADY, D. COURT and M. ROSENBERG, 1980 Structure and function of the cy control region of bacteriophage lambda. J. Mol. Biol. 138: 209-230.
WULFF, D. L., M. MAHONEY, A. SHATZMAN and M. ROSENBERG, 1984 Mutational analysis of a regulatory region in bacterio- phage X that has overlapping signals for the initiation of tran- scription and translation. Proc. Natl. Acad. Sci. USA 81: 555- 559.
ROSENBERG, M. and D. COURT, 1979
SHIMATAKE, H . and M. ROSENBERG, 1981
WULFF, D. L., 1976
![FIGURE ].-DNA are indicated. The SCHMEISNER the recognized by (1984), and the strands 1980)](https://thumb-us.123doks.com/thumbv2/123dok_us/1750367.1224431/2.609.138.473.71.185/figure-dna-indicated-schmeisner-recognized-strands.webp)