IDENTIFICATION OF AN R-LOCUS REGION THAT CONTROLS
THE
TISSUE SPECIFICITY O F ANTHOCYANIN FORMATION I N MAIZE1
HUGO K. DOONER
Laboratory of Geneiics, University of Wisconsin, Madison, Wisconsin 53706
Manuscript received July 10,1979
ABSTRACT
The R locus in maize controls the tissue specificity of anthocyanin forma- tion. Recombination between two different R-locus alleles with unique pig- mentation domains has resulted in the generation of a nonparental allelic type that displays a third kind of tissue specificity. These novel changes in tissue specificity occurred in very low frequency (1
x IOW).
In light of this finding and of recent results concerning the regulation by R of the enzyme UDPG:flavonoid 3-O-glucosyltransferase, specified by the Bz locus, a model dealing with the genetic control of tissue-specific functions in multicellular organisms is proposed.
manifestation of differentiation in higher organisms is the expression of
A
tissue-specific functions. Thus, the synthesis of chlorophyll takes place in shoots, but not in roots; that of hemoglobin occurs in erythrocytes, but not in leucocytes; and that of melanin is restricted to specialized epithelial cells. These are but a few examples of familiar tissue- or cell-specific functions in higher organ- isms. The many genes necessary for the biosynthesis of any one of the above pig- ments are present in all cells of the individual, yet the product of their concerted action accumulates only in certain cell types. The simplest interpretation of this observation, consistent with other experimental facts, is that the pertinent genes are activated only in specific tissues. This integrated tissue-specific gene activation must itself be under genetic control.Regulatory elements that activate batteries of genes were postulated by BRITTEN and
DAVIDSON
(1969) in their model of genetic regulation for higher organisms. However, as they pointed out, mutations in such regulatory loci characteristically would have pleiotropic effects on the activity of a number of other genes, and hence, would often turn out to be lethal. For example, mutations changing the tissue distribution of essential enzymes, such as RUdP carboxylase in photosynthetic plants, would not be expected to accumulate in natural popu- lations. On the other hand, allelic variants that affect the tissue localization ofnonessential secondary metabolites, such as the anthocyanin pigments, have been collected from diverse races of maize and have been extensively described (STADLER and FOGEL 1943,1945; STYLES, CESKA and SEAH 1973).
Many different genes affect anthocyanin synthesis i n maize (see STYLES and
1 The research was supported by the College of Ab?icultural and Life Sciences, University of Wisconsin-hIadison.
704 H U G O K. DOONER
CESKA 1977 for a recent review). Two of these, R(IOL) and
B
(ZL),
which are functionally duplicate genes, determine the specific plant or seed parts where anthocyanin is produced, i.e., R and B are unique factors that control the tissue distribution of anthocyanin formation (STYLES, CESKA and SEAH 1973).Allelic variability at the
R
locus is extensive (STADLER and FOGEL 1943,1945; STADLER 1948; VAN DER WALT and BRINK 1969; STYLES, CESKA andSEAH
1973). R alleles determine whether anthocyanin is synthesized or not, and those that are functional in some part of the plant vary i n tissue localization, distribution pattern within the tissue and concentration of the pigment. I n short, R variants differ as to whether, where, when and how much anthocyanin is synthesized. We are concerned here mainly with the R-locus function that determines i n what tissues anthocyanin formation occurs.Some R alleles have been found to be structurally complex. Thus, R-r: standard
is a tandem duplication consisting of a proximal member that carries a n anther (plant) pigmenting factor ( P ) and a distal member that carries a seed pig- menting factor (S); the distance between the ( P ) and (S) contrasting sites is 0.16 map units (STADLER and NUFFER 1953; STADLER and EMMERLING 1954; DOONER and KERMICLE 1971, 1974). An R-r allele of Ecuadorian origin (R-r: Ecuador 11 72) has associated with it a separable leaf-color factor (designated
Lc), which maps about 1.5 units distal to R (BRAY 1964). Lc evidently is a dis- placed R factor since it is borne on a chromosomal segment homologous with the R-r: standard duplicated segment (DOONER and KERMICLE 1976). Though some common tissues are pigmented by the action of ( P ) and
Lc
( e . g . , roots, coleoptile) coloration in the anthers uniquely indicates ( P ) action, whereas leaf color is a specific Lc effect. On the other hand, (S) action is totally nonoverlapping with that of ( P ) or Lc, being limited to the aleurone layer of the seed. Thus, ( P ),
(S) andLc
have unique pigmentation domains.It is shown i n this report that new (P)-type derivatives may be obtained by recombination between (S) and Lc, i.e., that changes in the R-locus-controlled tissue specificity of anthocyanin formation can arise by interallelic recombi- nation. In contrast, an attempt to detect new tissue specificities by mutation has yielded negative results (COE 1978, 1979). I n light of both the above finding and recent results (DOONER and NELSON 1977,1979) concerning the regulation by R of the enzyme UDPG: flavonoid 3-O-glucosyltransferase (UFGT)
,
which is specified by the Bz locus (9S), a model dealing with the genetic control of tissue-specific functions in multicellular organisms is proposed.MATERIALS A N D METHODS
R aUe2es: Because of the multiplicity of R-controlled pigment parameters, the designation of R alleles in the literature has not been entirely consistent. The R us. r disjunctive refers to t h e presence us. absence of pigment in the aleurone layer of the endosperm. The hyphenated second part of the symbol (earlier used in the form of a superscript) may designate presence
or absence of anther color ( - r = red anthers; -g = green anthers), anthocyanin distribution
in the aleurone (-nj, -st, -mb) or pericarp pigmentation (-ch) .
R D E T E R M I N E R S
O F TISSUE SPECIFICITY IN MAIZE 705used i n this study can be found in DOONER and KERMICLE (1974, 1976). Properties of R alleles that are pertinent to this presentation are outlined below.
R-mtandard: Anthocyanin pigment in the anthers (plus other plant parts) and in the aleurone layer of the seed. This allele is associated with a tandem duplication bearing the plant- pigmenting factor (P) in its proximal member and the seed-pigmenting factor (S) in its distal member. It can give rise, by unequal crossing over, to derivatives lacking one member of the duplication. Such derivatives are designated r-r if they retain (P) and lose (S), or R-g if, con-
versely, they retain (S) while losing ( P ) .
R-g:l: Colored aleurone, green plant parts, an (S) derivative from R-mtandard that has lost one member of the duplication by unequal crossing over. The number 1 following the gene symbol simply refers to the first of 15 R-g derivatives obtained at the University of Wisconsin from R-rrstandurd.
Lc: Color in several plant parts (e.g., coleoptile, roots, nodes and pericarp) but most con- spicuously in the leaf blade (BRAY 1964). This factor, extracted from a n Ecuador strain, maps 1.5 units distal to R and is borne on a displaced duplicate R segment. No evidence was obtained for fractionation of the compound phenotyFe conditioned by Lc (DOONER and KERMICLE 1976).
L c does not elicit aleurone pigmentation and is therefore functionally equivalent to a n r allele.
It closely resembles the r-ch allele of R, except that, unlike r-ch, Lc does not elicit anther color.
r-g: No anthocyanin pigment in any seed or plant parts. This null allele represents the basal member of the R allelic series.
Markers: Linked markers used in this study were g (golden), which maps about 20 units proximal to R, and Mst, the modifier of R-st (R-stippled), which maps 6 units distal to R. The unlinked factor, wx (waxy endosperm), was used as a male parent marker.
Crosses: Colorless seed exceptions were selected from among the colored kernel progeny of the cross g R-g:i Lc
+
/ G R-g:l Lc Mstx
g r-g IC f; wx. These selections were grown out,scored for anther color, Lc traits and the proximal marker golden, then crossed reciprocally with r-g; wx for transmission tests, and also used as the pollen source on R-st -I- plants to determine Mst constitution.
RESULTS A N D DISCUSSION
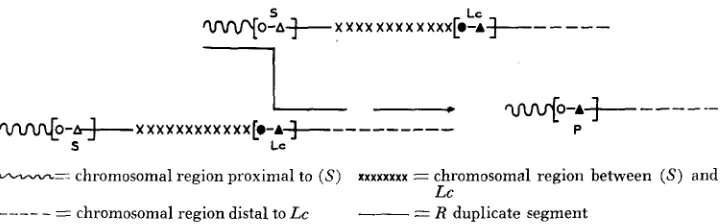
An R-g:l Lc chromosome carries in the distal one-fifth of the long a r m a direct displaced duplication, not unlike in size to the R-r: standard direct tandem dupli- cation, whose noncontiguous proximal and distal members are marked respec- tively by S and Lc, as diagrammed below.
*o*/*;
S LC
-0J5-e -1.5
--
0.15-
I n R-g:l Lc homozygotes, following oblique synapsis of the (S) and
Lc
seg- ments, recombination distal to the respective R markers in the displaced paired segments yields (S) derivatives that are deficient for Lc and for the 1.5-unit-long region between ( S ) and Lc. Utilizing a n efficient seedling screening procedure,DOONER
andKERMICLE
(1976) obtained R-g Zc crossover derivatives, which retain seed color and lose Lc traits, with a frequency of approximately 5x
IO4.I n addition to R-g Zc types, colorless seed ( r ) exceptions are generated from the cross g R-g:Z Lc+
/
G R-g:Z Lc Mst X g r-gIC
+;
wx. Table 1 reports the characterization of these colorless seed selections. R-g: 1 is known to mutate to r-g With a frequency of about 1 in 10,000 gametes (BRAY and BRINK 1966; DOONER706 H U G O K. DOONER
tion of R-g: 1 to r-g; their frequency (1.1 X IO-.) is very close to that anticipated, and they do not appear to arise in relation to outside marker exchange (ten recombinants were observed, whereas 8.5 should have occurred assuming that
R-g + r-g mutation and recombination in the g to Mst region are independent
events).
The second class of T derivatives from R - g : l is unexpected: r-r Zc derivatives
lack both seed and leaf color, but have pigmented anthers. Although pigment intensities have not been compared, the tissue distribution of anthocyanin in the new r-r variants appears to be very similar to that of other r-r alleles. Pigmented tissues include roots, coleoptile, seedling leaf-tip and anthers. That is, the r-r
IC
derivatives have lost both (8) and Lc function, while gaining at the same time a (P)-type function. They occur in very low frequency (1 X and their origin is associated with outside marker exchange.Evidence that recombination occurs within the obliquely synapsed (S) and
Lc
displaced duplicate segments derives from the following consideration. Like the crossover R-g IC derivatives described above, these r-r
IC
derivatives should lack the 1.5-unit-long-region between (S) and Lc.DOONER
andKERMICLE
(1976) reported that chromosomes deficient for this region show reduced male trans- mission relative to a normal chromosome. This same transmission behavior was displayed by the r-rIC
selections from R - g : lLc.
The average ratio of red to green seedlings from the testcrosses r-gIC
X r-rIC
/
r-g IC was found to be 1:1.2, which represents a deviation from 1:l very similar to that reported byDOONER
andKERMICLE
(1976). In contrast, counterpart testcrosses of the sib r-gLc
selections gave equal numbers of Lc andIC
seedlings. Though the opposite recombinants for flanking markers should occur equally if r-r IC derivatives arise by unequal exchange, only one recombinant type was observed in the present experiment. The 3:O split is not significantly deviant from 1:1, and the absence of the other recombinant type is probably due to small sample size.The preceding account describes a novel phenomenon: change in the tissue specificity controlled by a locus brought about by interallelic recombination at that locus. This event is diagrammed in Figure 1. The R-locus region controlling tissue specificity is designated by brackets as a subregion within the larger dupli- cate segment. Since recombination can occur in this region and the recombinants display a novel type of specificity not evidenced by the R - g : l Lc parent, the region is taken to represent a cis-acting unit that differs in (S) and Lc at a mini- mum of two sites. A recombination event that brings the proximal (S) site into
TABLE 1
Colorless seed (r) derivatives from marked R-g:l Lc females crosspd to g r-g :c
+;
wx malesOutside marker constitution Class of Parental Recombinant
Parent genotype Population derivative G Mst g 4- G
+
R M s t Total G H-g:l Lc Mst 312,200 r-g Lc 10 14 3 7 34g R-g:l Lc
+
R DETERMINERS OF TISSUE SPECIFICITY I N MAIZE 70 7
S Lc
w O - A * X X X X X X X X X X X X [ O - A e -
-
--
-
@-A+
- -
-
-
--W O - A * S X X X X XX X
1-
X X X X X [.-A+ Lc--
- - -
----
PA,---.= chromosomal region proximal to (S) xxxxxxxx = chromosomal region between (S) and
Lc
-__- - - - chromosomal region distal to Lc
--
= R duplicate segmentFIGURE 1.-Origin of r-r IC derivatives from R-g:l Lc homozygotes. The brackets in the duplicate segment designate a cis-acting unit in the R locus that controls tissu2 specificity. T h e determinants of tissue specificity in (S) and Lc differ in at least two sites that are recom- binationally separable. The juxtaposition of the proximal site in (S) with the distal site in LC by recombination generates a novel (P) -type Specificity not evidenced by either parental allele.
cis-association with the distal Lc site generates the (P)-type specificity seen in the crossover r-r IC derivatives. The generation of unpredictable phenotypes by intragenic recombination between co-dominant alleles at a locus has also been discussed by SHEPHERD and MAYO (1 972) with reference to genes conferring disease resistance in certain plants.
Alternately, it might have been argued that when R-g:l was formed by unequal crossing-over in R-r, i.e., ( P ) ( S ) homozygotes, the original ( P ) tissue- specific region was not completely lost and that it is now being reactivated or restored by the new recombinational event. However, similar r-r IC derivatives also occur in R-g:6 Lc homozygotes. R-g:6 has a ( p ) (S) constitution and arose from R-r by mutation of
( P )
to ( p ).
Therefore, R-g: 6 can be presumed to carry the unaltered (S) component of R-r. R-g:6 Lc homozygotes yield a ( p ) (P) type of r-rlc derivative that can be explained by the same sort of mechanism as depicted in Figure 1 for R-g:l Le. Thus, the occurrence of ( P ) derivatives by intragenic recombination between(8)
and Lc is not a function of the R-g allele used in the test.DOONER and KERMICLE (1971, 1974) have mapped a site distinguishing
( P )
708 HUGO K. DOONER
Analysis of unstable (P) alleles [ (r-r(Z,): ASHMAN 1970; R-g:6; DOONER, unpublished; R-g: 9 ; KERMICLE, unpublished) has revealed that mutations in sites distal to the ( P ) determinant of tissue specificity cause loss of (P) function.
In
at least one case (Z-R of R-st), transfer of the mutant site by interallelic recombination to other alleles (R-sc,R-ni,
R-p 3) has resulted in nullification of the function of those alleles (ASHMAN 1970; KERMICLE 1970; GAVAZZI 1977). These recombination events are at least one order of magnitude more frequent than those giving rise to r-rIC
inR-g:l
Lc. Thus, the cis-acting components ofR
appear to include a proximal region defined by tightly linked sites controlling tissue specificity and a distal region defined by sites that, when mutated, cause impairment of R function, and that may be more easily separable from the tissue- specificity sites than the laiter are from each other. Cis-acting regulatory genetic determiners, very closely linked to genes of known function, have been reported in several higher organisms. These determiners may regulate properties such as the level (CHOVNICK, GELBART and MCCARRON 1977; BREEN, LUSIS and PAIGEN1977) and time of synthesis during development (SCHWARTZ 1962) of several en- zymes, as well as the relative degree of expression of two alleles in a heterozygote
( SCHWARTZ 1976).
The primary function of the
R
locus is not known. Recently, however, DOONER and NELSON (1977,1979) have found that the enzyme specified by the Bz locus(9s)
,
UDPG: flavonoid 3-O-glucosyltransferase (UFGT),
which is known to catalyze a late step in anthocyanin biosynthesis, fails to be induced in r (color- less) endosperms during development. Therefore,R
can be considered as a regulatory locus determining the tissue-specific synthesis of UFGT (and, con- ceivably, of other anthocyanin biosynthetic enzymes.) A possible mechanism by which factors, such as R, may control the tissue specificity of a given function is proposed below.Two common types of elements postulated in models of genetic regulation are specific signals for gene activation and specific receptors for such signals, often in a hierarchical type of organization (e.g., BRITTEN and DAVIDSON 1969,1979). Such integrated hierarchies of signals and receptors readily come to mind when possible mechanisms for the tissue-specific activation of a gene are considered. Thus, it stands to reason that there exists at the Bz locus a receptor for a n R-
dependent signal that mediates activation of Bz. Such activation could occur via
a n R-coded regulatory protein or RNA (cf., FRENSTER 1979) or, indirectly, via
a flavonoid intermediate resulting from an R-specified enzymatic function. The
R
signal would be common to all R-locus alleles that are active in one or another tissue, and would be under the control of the distal segment of the gene.The control of tissue specificity resides in a cis-acting, proximally located R
region, variable among R locus alleles, that is not involved in the determination of the Bz-activating signal, but that, instead, determines in what tissues the signal is produced. This region would function as a specific receptor for signals that are unique to particular tissues (though their origin may trace ultimately to
R
DETERMINERS O F TISSUE SPECIFICITY IN MAIZE 709allelic diversity, stable quantitative variation among
R
alleles with respect to anthocyanin pigmentation in a particular tissue can be accounted for on the basis of the relative affinity of the R tissue-specificity region for the regulatory sig- nal(s) characteristic of that tissue. Likewise, if the concentration of such a signal varies during development of the tissue, observed variations in time of onset or termination of activity (STYLES, CESKA andSEAH
1973) can also be explained.For reasons of cellular economy, it would be of advantage to regulate the cell- specific synthesis of a compound at a stage in its biosynthetic pathway immedi- ately following the branchpoint that separates this from other compounds that, though related, may have an altogether different cellular distribution. Thus, the enzymes needed to process a common precursor into a cell-specific product would be synthesized only in those cells where the product accumulates. I n the bio- synthesis of phenolics, p-coumaroyl CO-A constitutes a critical branchpoint, beyond which the first enzyme unique to flavonoid biosynthesis is flavanone synthase (HAHLBROCK and GRISEBACH 1975). This enzyme has recently been found to be deficient in r developing endosperms (DOONER, unpublished). Con- ceivably, R may determine in what tissues anthocyanin is formed by controlling the tissue-specific production of flavanone synthase and other enzymes, such as UFGT, acting later in the anthocyanin biosynthetic pathway. The model pre- sented here may be of general applicability to the biosynthesis in higher organ- isms of other compounds that are restricted to specific cell or tissue types. How- ever, events such as those described in this text would almost invariably escape detection since they involve infrequent recombination between alleles of an unusual type of locus controlling, as R does, the tissue-specific formation of a particular compound. Only if this compound is nonessential, as is anthocyanin in maize, can one expect allelic variation at such a locus to be maintained in nature.
I am grateful to R. A. BRINK, J. L. KERMICLE and KO SHIMAMOTO for helpful criticisms of the manuscript and to J. L. KERMICLE for sharing unpublished data. Part of this work was iunded through DOE grant EY76S-02-1300.
LITERATURE CITED
ASHMAN, R. B., 1970
BRAY, R. A., 1964
38: 134.
The compound structure of the R-st allele in maize. Genetics 64: 239-
A plant color factor linked to the R locus. Maize Genet. Coop. News Letter
Mutation and paramutation at the R locus in maize. Genetics
Linkage of genetic determinants for mouse &galactosidase electrophoresis and activity. Genetics 85 : 73-84.
Gene regulation for higher cells: a theory. Science 165: 349-357. -, 1979 Regulation of gene expression: possible role of repetitive sequences. Science 204: 1052-1059.
245.
BRAY, R. A. and R. A. BRINK, 1966
BREEN, G. A., A. J. LUSIS and K. PAIGEN, 1977
710 H U G O K. DOONER
CHOVNICK, A., W. GELRART and M. MCCARRON, 1977
COE, E. H., 1978
Organization of the rosy locus in Dro- sophila melanogaster. Cell l l : 1-10.
Determinants of allelic variation in tissue specificity in maize. (Abstr.) Proc. XIV Internat. Cong. Genetics: 574. --, 1979 Specification of the anthocyanin bio- synthetic function by B and R i n maize. Maydica 24: 49-58.
Structure of the R-r tandem duplication in maize. Genetics 67: 427-435. -, 1974 Reconstitution of the R-r compound allele in maize. Genetics 78: 691-701.
-
, 1976 Displaced and tandem duplications in the long arm of chromosome 10 in maize. Genetics 82 : 309-322.Genetic control of UDP glucose: flavonol 3-0-glucosyl- transferase i n the endosperm of maize. Biachem. Genet. 15: 509-519. __ , 1979 Inter- action among C , R and V p in the control of the Bz glucosyltransferase during endosperm development in maize. Genetics 91 : 309-315.
Selective gene derepression by derepressor RNA. In: Eukaryote Gene Regulation. Edited by G. M. KOLODNY. Chemical Rubber Co. Press, Cleveland.
The genetic complexity of the R locus i n maize. Stadler Symp. 9 : 37-61. Biosynthesis of flavonoids, pp. 866-915. In: The Flavonoids. Edited by J. B. HARBONE, T. J. MABRY and H. MABRY. Academic Press, New York.
Somatic and meiotic instability of R-stippled, an aleurone spotting factor in maize. Genetics 64: 247-258.
Genetic studies on mutant enzymes in maize. 111. Control of gene action
in the synthesis of pH 7.5 esterase. Genetics 47: 1609-1615.
-
, 1976 Regulation of expression of Adh in maize. Proc. Natl. Acad. Sci. U.S. 73: 582-584.Genes conferring specific plant disease resistance. Science 175: 375-380.
Spontaneous mutation at the R locus in maize. 11. Race differences in
Problems of gene structure. 111. Relation of unequal crossing over to the interdependence of R-r elements (S) and (P) . (Abstr.) Science 119: 585.
Gene variability in maize. I. Some alleles of R-r (R-r series) (Abstr.) Genetics 28: 90-91 -, 1954 Gene variability i n maize. 11. The action of certain R alleles. (Abstr.) Genetics 30: 23-24.
STADLER, L. J. and M. G. NEUFFER, 1953 Problems of gene structure. 11. Separation of R-r ele- ments (S) and (P) by unequal crossing over. (Abstr.) Science 117: 471472.
STYLES, E. D. and 0. CESKA, 1977 The genetic control of flavonoid synthesis in maize. Can. J.
Genet. Cytol. 19: 289-302.
STYLES, E. D., 0. CESKA and K. T. SEAH, 1973 Developmental differences in action of R and B alleles in maize. Can. J. Genet. Cytol. 15: 59-72.
VAN DER WALT, W. J. and R. A. BRINK, 1969 Geographic distribution of paramutable and para-
mutagenic R alleles in maize. Genetics 61 : 677-695.
Corresponding editor: R. L. PHILLIPS
DOONER, H. K. and J. L. KERMICLE, 1971
DOONER, H. K. and 0. E. NELSON, 1977
FRENSTER, J. H., 1979
GAVAZZI, G., 1977
HAHLBROCK, K. and H. GRISERACH, 1975
KERMICLE, J. L., 1970
SCHWARTZ, D., 1962
SHEPHERD, K. W. and G. M. E. MAYO, 1972
STADLER, L. J., 1948
mutation rate. Am. Naturalist 82: 289-314. STADLER, L. J. and M. H. EMMERLING, 1954