INVESTIGATION
A Distorted Circadian Clock Causes Early Flowering
and Temperature-Dependent Variation in Spike
Development in the
Eps-3A
m
Mutant of
Einkorn Wheat
Piotr Gawron´ski,* Ruvini Ariyadasa,* Axel Himmelbach,* Naser Poursarebani,* Benjamin Kilian,* Nils Stein,* Burkhard Steuernagel,*,1Goetz Hensel,* Jochen Kumlehn,* Sunish Kumar Sehgal,‡ Bikram S. Gill,‡Peter Gould,†Anthony Hall,†and Thorsten Schnurbusch*,2 *Leibniz Institute of Plant Genetics and Crop Plant Research, OT Gatersleben, D-06466 Stadt Seeland, Germany,†School of Biological Science, University of Liverpool, Liverpool L69 3BX, United Kingdom, and‡Department of Plant Pathology, Wheat Genetics Resource Center, Kansas State University, Throckmorton Plant Sciences Center, Manhattan, Kansas 66506-5502
ABSTRACT Viable circadian clocks help organisms to synchronize their development with daily and seasonal changes, thereby providing both evolutionaryfitness and advantage from an agricultural perspective. A high-resolution mapping approach combined with mutant analysis revealed a cereal ortholog ofArabidopsis thaliana LUX ARRHYTHMO/PHYTOCLOCK 1(LUX/PCL1) as a promising candidate for theearliness per se 3(Eps-3Am) locus in einkorn wheat (Triticum monococcumL.). Using delayedfluorescence measure-ments it was shown that Eps-3Amcontaining einkorn wheat accession KT3-5 had a distorted circadian clock. The hypothesis was subsequently confirmed by performing a time course study on central and output circadian clock genes, which showed arrhythmic transcript patterns in KT3-5 under constant ambient conditions,i.e., constant light and temperature. It was also demonstrated that variation in spikelet number between wild-type and mutants is sensitive to temperature, becoming negligible at 25°. These observa-tions lead us to propose that the distorted clock is causative for both earlyflowering and variation in spike size and spikelet number, and that having a dysfunctionalLUXcould have neutral, or even positive, effects in warmer climates. To test the latter hypothesis we ascertained sequence variation ofLUXin a range of wheat germplasm. We observed a higher variation in the LUX sequence among accessions coming from the warmer climate and a unique in-frame mutation in early-flowering ChineseT. turgidumcultivar‘Tsing Hua no. 559.’Our results emphasize the importance of the circadian clock in temperate cereals as a promising target for adaptation to new environments.
T
HE circadian clock is an intrinsic regulator of biological processes oscillating within an24-hr period (Pittendrigh 1993). It is considered to be the main mechanism by which plants recognize the optimal photoperiod for seasonalflowering (Imaizumi 2009). Transcriptional regulation of the circadian clock has been well described in Arabidopsis (Pokhilkoet al.2012) with the latest model emphasizing the importance of the Evening Complex (EC) composed of EARLY FLOWERING 3, EARLY FLOWERING 4, and LUX AR-RHYTHMO/PHYTOCLOCK 1 (ELF3, ELF4, and LUX/PCL1) proteins (Onai and Ishiura 2005; Nusinow et al. 2011; Pokhilkoet al.2012). The EC directly represses the function ofPSEUDO RESPONSE REGULATOR 9(PRR9) (Helferet al. 2011) and acts antagonistically to the elements expressed in the morning, including LATE ELONGATED HYPOCOTYL (LHY) and CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) (Pokhilkoet al.2012).LHY,CCA1, andPRR9form the so-called morning loop, which becomes arrhythmic when the EC is impaired (Hazen et al. 2005; Dixon et al. 2011; Nusinow Copyright © 2014 by the Genetics Society of America
doi: 10.1534/genetics.113.158444
Manuscript received November 10, 2013; accepted for publication January 9, 2014; published Early Online January 17, 2014.
Supporting information is available online athttp://www.genetics.org/lookup/suppl/ doi:10.1534/genetics.113.158444/-/DC1.
New nucleotide sequences were deposited in the GenBank under accession nos. KF769443–KF769445.
1Present address: Sainsbury Laboratory, Norwich Research Park, Norwich NR 7UH, United Kingdom.
2Corresponding author: Leibniz Institute of Plant Genetics and Crop Plant Research, Corrensstrasse 3, OT Gatersleben, D-06466 Stadt Seeland, Germany.
E-mail: [email protected]
et al. 2011). The EC also down-regulates transcription of evening genes such as GIGANTEA (GI) and TOC1 (Hazen et al.2005; Dixonet al.2011; Pokhilkoet al.2012).
Genetic studies have shown that the recently cloned ma-turity-a, (mat-a; syn. early maturity 8, eam8) locus is an ortholog of AtELF3 in barley (Hordeum vulgare L.). eam8 appears to be epistatic toeam10 (syn. easp), which in turn
seems to be a possible ortholog ofLUX/PCL1(Gallagheret al. 1991; Zakhrabekova et al.2012; Campoli et al.2013). This would be consistent withEam10andEam8in barley forming a complex similar to AtLUX and AtELF3 (and AtELF4) in Arabidopsis (Nusinowet al.2011). Interestingly, the genetic location ofeam10in barley andearliness per se 3Am(Eps-3Am)
in einkorn wheat (Triticum monococcum L.) appears to be syntenic (Gallagheret al.1991; Börneret al.2002; Gawron´ski and Schnurbusch 2012). Both mutants also display similar phenotypic features and flower early under both long and short day conditions, thus resembling mutants in the Arabi-dopsis EC (Hickset al. 2001; Shindo and Sasakuma 2001; Doyle et al. 2002; Hazenet al. 2005; Zakhrabekova et al. 2012). Moreover, recent analysis of eam8 (HvELF3) and eam10 (HvLUX1) mutants showed misexpression of circa-dian clock gene-related transcripts (Faureet al.2012; Cam-poliet al.2013), which was in line with previousfindings in Arabidopsis(Hickset al.2001). So far, cerealPCL1/LUXwas proposed as a candidate conferring earlyflowering ineam10 and KT3-5 mutants (Mizuno et al. 2012; Campoli et al. 2013). Authors have concluded that the mechanism of the PCL1/LUX protein action is similar in both einkorn wheat and barley; it represses expression of Vrn3/FTthrough the negative regulation of Ppd-1/Ppd-H1, thus delayingfl ower-ing. When PCL1/LUX is knocked-out, it causes the early heading phenotype, especially when the mutant harbors the wild-type (wt) allele ofPpd-H1;i.e., photoperiod sensi-tivity (Mizunoet al.2012; Campoliet al.2013).
Recently, delayed fluorescence (DF) has been proposed as a method for quick but high-resolution analysis of the circadian clock in virtually any species (Gould et al. 2009). The DF phenomenon, discovered by Strehler and Arnold (1951) is a luminescence produced by the photosynthetic apparatus after excitation with ambient light as a result of charge recombina-tion in the photosystem II (PSII) and subsequent emission of a photon (Rutherford et al.1984). DF allows measuring the intrinsic oscillation in chlorophyll fluorescence (PSII) that remains under circadian control (Gouldet al.2009). The value of DF measurements in cereal mutant research has not, to our knowledge, been reported in the literature. However, it appears to have considerable potential for high-throughput analysis in circadian clock research.
Here we present more detailed genetic and physiological analyses of theEps-3Ammutant in diploid wheat KT3-5. Fine
mapping and comparative genetic analyses with barley and bread wheat provide more confirmation that the TmLUX/ PCL1 is the most sensible candidate conferring the early flowering phenotype. Moreover, we present physiological evidence that KT3-5 possesses a distorted circadian clock
and has a decreased phenotypic plasticity, in agreement with the view that variation in the clock is an effective adaptive mechanism (Dodd et al. 2005). We also mark the impor-tance of the circadian clock for adaptation to temperature, in addition to photoperiod perception as discussed else-where (Faureet al.2012; Zakhrabekovaet al.2012).
Materials and Methods
High-resolution mapping
Seeds of the F2population consisting of 658 individuals, as
well as parental lines recombinant inbred line (RIL) RILWA25 and RILWA71, were germinated on Petri dishes. To synchro-nize germination, soaked seeds were kept at 4°for 2 days. Seedlings of selected recombinant individuals plus 38 ran-dom plants were transplanted and placed in a cool room (15°, photoperiod of 11 hr light (L)/13 hr dark (D)) for 2 weeks, followed by cold treatment (4–8°) for 3 weeks and a photoperiod of 10 hr L/14 hr D. For acclimation, plants were moved back to cool temperature (15°, photoperiod of 11 hr L/13 hr D) for 2 weeks and were transplanted to 1.1-liter pots. After transplanting, the LD treatment (16 hr/8 hr) started at 18°/15°until harvest. Each pot was manually ran-domized every 3–4 days to avoid positional bias. Heading time was scored when the visible part of awns on the main culm was about 1 cm long. A total of 658 individuals were screened withflanking markers INDEL_271_272 and INDEL_201_202 giving size polymorphism (Supporting Information,Table S1). Plants with a recombination event betweenflanking markers were further genotyped with markers listed inTable S1and Table S2. Marker development was performed according to the procedure described elsewhere (Gawron´ski and Schnurbusch 2012).
Analysis in einkorn wheat mutants
Growing conditions (greenhouse), phenotypic analyses, DNA extraction and PCR genotyping were conducted as previously reported (Gawron´ski and Schnurbusch 2012). Mutants and wild-types analyzed are listed inTable S3.
Delayedfluorescence measurement
Leaf samples were taken from 4-week-old plants, cut into 2-cm pieces andfloated on sterile distilled water containing 18 mg/liter fungicide Dithane poured into a 25-mm com-partmental Petri dish. At dusk (10:00 PM, ZT = 12, where
ZT means “Zeitgeber time”) dishes were put into a Sanyo MIR-553 cooled incubator (Sanyo Gallenkamp) and the DF was imaged using an ORCA-II-BT 1024 16-bit camera (Hamamatsu Photonics) cooled to280°. However, the“zero” time point was set at dawn next day (10:00 AM, ZT = 0).
Leaf pieces were kept in darkness intermitted every hour with a pulse of red/blue light (80 mmol/m2s) lasting for
1 min. Immediately after the pulsing, the picture was cap-tured allowing for assaying the circadian clock output with 1-hr resolution. The whole process was automated by using
Wasabi software (Hamamatsu Photonics) for controlling the camera and light source (light-emitting diodes). The temper-ature during the experiments was kept constant at 22°or 17°. The pictures were analyzed in Metamorph 6.0 (Universal Imaging) and the numerical light intensities were extracted. Obtained data were normalized and detrended using Excel (Microsoft). Regression equation of the polynomial trend line in the order of six was used to subtract Yvalues (delayed fluorescence) thus removing the trend. The data showed a pattern of up to six peaks of DF. Thefirst peak of DF needed to be discarded as it was out of phase. Data collected between hours 36 and 132 (normalized but not detrended) were used to calculate periods corresponding to the four middle peaks of the delayed fluorescence. The periods at 95% confidence interval and relative amplitude errors (RAEs) were calculated in BRASS ( http://millar.bio.ed.ac.uk/PEBrown/BRASS/Brass-Page.htm) by running fast Fourier transformed nonlinear least-square analysis (Plautzet al.1997).
Time-course RT-qPCR
Seeds of DH_BG284E11-PP1, BG353/1E15 (File S1), T. monococcumKT3-1 and KT3-5,T. turgidum‘Tsing Hua no. 559’ and‘Fo Shou Mai,’and azygous segregants of BG284E11-38 were soaked in short day (SD) water and kept in the cold (4°–8°) to synchronize germination. After 2 days, young seedlings were moved to room temperature for 2 days and planted in soil (150-ml wells). Plants were grown in the greenhouse at 20°/17°day/night under a photoperiod of 16 hr for 4 weeks. In the actual phase of the experiment, plants were moved to the incubator Heraeus Vötsch, type HPS 1500/S. For the following 2 days conditions were changed to 12 hr/12 hr at 18°/22°to synchronize the cir-cadian clocks of the plants. In the last stage of the experi-ment, constant light and temperature (22°) were set for 3 days. Sampling was performed on 31-day-old plants starting at 6:00 AM of the second day under constant light and
temperature. Leaves were harvested every 3 hr (LL con-ditions) from at least three plants per time point per geno-type (biological replicates) and immediately frozen in liquid nitrogen. RNA extraction was performed by using PureLinkRNA mini kit (Invitrogen) in combination with TRI-zol reagent (Invitrogen) as described by the manufacturer. QuantiTect Reverse Transcription kit (Qiagen) was applied to synthesize cDNA using 1mg of total purified RNA. qPCR reactions were performed on the ABI 7900HT Fast Real-Time PCR system (Applied Biosystems) employing Quanti-Tect SYBR Green PCR kit (Qiagen) at the following thermal profile: 94°for 15 min, five cycles of 94°for 30 s, 65°for 30 s decreasing by 1°per cycle, and 72°for 30 s, and then 40 cycles of 94°for 30 s, 60° for 30 s, 72° for 30 s, and 72° for 1 min. Primers used to amplify clock genes are listed in the Table S1. Gene nomenclature and sequences used for primer development were as published elsewhere (Campoli et al. 2012). TtLUX-A and TtLUX-B, A and B ge-nome specific primers, respectively, were developed initially by using the LUX sequences from following databases:
http://www.wheatgenome.org/ and http://www.cshl.edu/ genome/wheat. The genome specificity was confirmed by amplification of the TtLUX from flow-sorted chromosome arms of cv.‘Chinese Spring.’
Detailed methods for physical mapping, DNA gel blot analysis, growing conditions of KT3-1 and KT3-5, targeting induced local lesions in genomes (TILLING), development and analysis of knock-down lines, andLUXresequencing are included in File S1(Materials and Methods).
Results
Phenotypic characterization of KT3-5
KT3-5 is an Eps-3Amearlyflowering einkorn wheat mutant
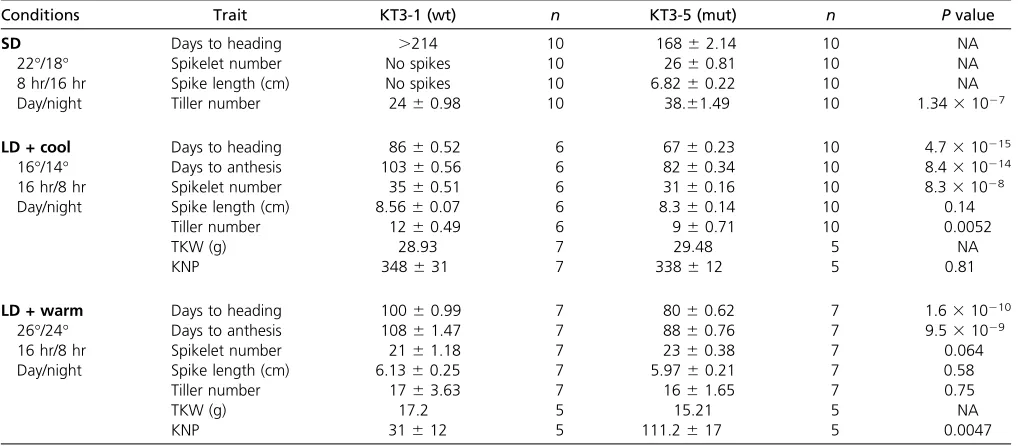
derived from a donor line called KT3-1. We phenotypically analyzed KT3-5 and KT3-1 in different environments and showed that KT3-5flowered early in our experimental con-ditions, irrespective of photoperiod (Table 1 and File S2), consistent with observations made in a previous study (Shindo and Sasakuma 2001). We found that differences between wild-type and mutant plants were exaggerated when the plants were exposed to SD treatment (8-hr pho-toperiod, Table 1). Immature spike investigations under long-day (LD, 16 hr/8 hr day/night) and cool (16°/14°) growing conditions revealed that KT3-5 transitioned to the generative phase (double-ridge stage, P= 0.028) 10 days earlier than KT3-1 and to the terminal spikelet stage 7 days in advance (terminal spikelet stage,P= 0.0085).
It has previously been postulated thatepsloci can be neg-atively associated with spike characters including spike length and spikelet number depending on temperature (Lewiset al. 2008; Gawron´ski and Schnurbusch 2012). At LD (16°/14°) we observed that KT3-1 produced on average four spikelets more than KT3-5 (P= 8.331028) but this did not signifi
-cantly affect the difference for the kernel number per spike (KNP,P= 0.81) (Figure 1A, Table 1). At higher temperatures during early development, fewer spikelets developed on both KT3-1 and KT3-5 inflorescences. When plants were grown at LD and 26°/24°, both KT3-1 and KT3-5 produced a similar number of tillers, similar number of spikelets per spike, and spike sizes (Table 1, File S2, and Figure 1A). However, the KT3-5 mutant carryingEps-3Amheaded 20 days earlier and
produced significantly more grains per spike (KNP). Thus, Eps-3Amdoes not appear to negatively influence yield-related
spike characters when plants are exposed to high tempera-tures during early spike development and there was a signif-icant interaction between genotype and temperature during spike development (Table 1 and Figure 1A).
Genetic analysis of Eps-3Am
Initial phenotyping and high-density mapping of Eps-3Am
was performed in a recombinant inbred line (RIL) popula-tion derived from a cross between KT3-5 (spring type) and wild-type line KT1-1 of T. boeoticum Boiss. (winter type) (Gawron´ski and Schnurbusch 2012). Two selected
type lines from this population, RILWA25 and RILWA71, were then crossed to create an F2population for
high-reso-lution mapping. The population consisted of 658 individuals and was screened with two markers flankingEps-3Am(File
S3). Thirty-eight recombinants were identified in this initial mapping and these were subsequently used to delimit Eps-3Amto a sub-cM level. As foream10in barley (Campoliet al.
2013), a putative ortholog of AtLUX ARRHYTHMO (Hazen et al.2005; Onai and Ishiura 2005) (i.e.,TmLUX) cosegre-gated with theflowering phenotype. Phenotypic distribution of Eps-3Am(8 early:30 late) was not significantly different
from 1:3, thus being in agreement with a single-gene-model inheritance (P= 0.57,x2= 0.316).
We grew all recombinant plants plus 38 randomly chosen F2’s and showed that the distribution of flowering time
agreed completely with the genotypic status at theTmLUX gene (Figure 1B). PCR-based screens of barley BACs using molecular markers flankingEps-3Am identified barley
con-tig_95 on the recently released physical map (Mayer et al. 2012) (Table S2,File S4, and Figure 2A). Additional marker analyses then enabled us to narrow the region containing Eps-3Amto a syntenic region in barley that contained only
two putative genes, HvPUMILIO (GenBank accession no. KF769445) and HvLUX (Figure 2A). F2 individuals having
the closest recombination events (seven proximal, one dis-tal), were advanced to the F3 generation (File S3). This
allowed us to confirm the association between heading time (Eps-3Am) and presence/absence ofTmLUX(Figure 1C). We
were able to discard HvPUMILIO as Eps-3Am by TILLING.
Screening for mutations in HvPUMILIO identified a single mutant that carried a premature stop codon (Figure S1,File
S5). Importantly this mutant did not exhibit an earlyfl ower-ing phenotype (Figure S1). Finally, to address the possibility that other nonsyntenic genes might have been present at Eps-3Am in einkorn wheat and may be responsible for the
phenotype, we built a physical map from bread wheat chro-mosome 3A (T. aestivumL. cv.‘Chinese Spring,’(Figure 2A, File S6, and File S7) and identified two contigs that were syntenic toEps-3Am. In terms of sequence conservation, we
observed that the Eps-3Am locus in einkorn wheat had
higher resemblance to the syntenic barley sequence than to that of bread wheat (Figure 2A andTable S2).
During our genetic characterization ofEps-3Am, we noted
that two adjacent marker loci, TmLUX(PAV_261_262) and TmPUMILIO(PAV_295_296), were scored as null alleles in KT3-5 (Figure 2A), suggesting a putative deletion of both genes in agreement with previous findings (Mizuno et al. 2012). DNA gel blot analysis confirmed this result (Figure S2). We then identified 15 additional X-ray early mutants of einkorn wheat with null alleles at both loci (Table S3). Hap-lotype analysis revealed the presence of four groups, with null allele at markerflankingEps-3Amin KT3-10, suggesting
a minimum of two independent deletion events being pres-ent in the deletion panel (Table S4). All 15 mutantsfl ow-ered significantly earlier than KT3-1, with the differences ranging from 25 up to 60 days (Table S4).
Circadian clock distortion in KT3-5
DF measurements have been shown to mirror CAB2:LUC reporter analysis in circadian clock mutants ofArabidopsis. Importantly, DF has no requirement for transgenesis, which was an important advantage while working with einkorn Table 1 Phenotypic analysis of einkorn wheat lines
Conditions Trait KT3-1 (wt) n KT3-5 (mut) n Pvalue
SD Days to heading .214 10 16862.14 10 NA
22°/18° Spikelet number No spikes 10 2660.81 10 NA
8 hr/16 hr Spike length (cm) No spikes 10 6.8260.22 10 NA
Day/night Tiller number 2460.98 10 38.61.49 10 1.3431027
LD + cool Days to heading 8660.52 6 6760.23 10 4.7310215
16°/14° Days to anthesis 10360.56 6 8260.34 10 8.4310214
16 hr/8 hr Spikelet number 3560.51 6 3160.16 10 8.331028
Day/night Spike length (cm) 8.5660.07 6 8.360.14 10 0.14
Tiller number 1260.49 6 960.71 10 0.0052
TKW (g) 28.93 7 29.48 5 NA
KNP 348631 7 338612 5 0.81
LD + warm Days to heading 10060.99 7 8060.62 7 1.6310210
26°/24° Days to anthesis 10861.47 7 8860.76 7 9.531029
16 hr/8 hr Spikelet number 2161.18 7 2360.38 7 0.064
Day/night Spike length (cm) 6.1360.25 7 5.9760.21 7 0.58
Tiller number 1763.63 7 1661.65 7 0.75
TKW (g) 17.2 5 15.21 5 NA
KNP 31612 5 111.2617 5 0.0047
Conditions were applied from sowing until harvest. NA, not applicable; TKW, thousand kernel weight, measured with 0.001-g precision; KNP, kernel number per plant. KT3-5 mutant was grown along with the donor line KT3-1 under various conditions. Although the KT3-KT3-5 mutant alwaysflowered earlier than KT3-1, difference in the spikelet number depended on the ambient temperature; KT3-1 developed more spikelets than KT3-5 at lower temperatures but the situation was different at higher temperatures. All values represent mean6SEM (standard error of the mean).Pvalues were calculated by using paired Student’st-test.
wheat (Gouldet al.2009). We hypothesized that ifTmLUX was functionally orthologous toAtLUX (Hazenet al.2005; Onai and Ishiura 2005), then a complete knock-out of the TmLUXshould cause circadian clock distortion. To test this hypothesis, we measured DF in 5 and its wild-type KT3-1 (File S8). At 22°DF oscillation generally ceased in the KT3-5 after 2 days (Figure 3, A and B). Samples that remained rhyth-mic displayed significant amplitude lengthening to 28 hr compared to 23 hr in the wild-type (Figure 3, B and C,Table S5). At 17°the differences were less pronounced (Figure 3D). Nevertheless we conclude that DF is appropriate for measuring circadian clock distortion in einkorn wheat, easily revealing the perturbed clock phenotype in the KT3-5 einkorn wheat mutant.
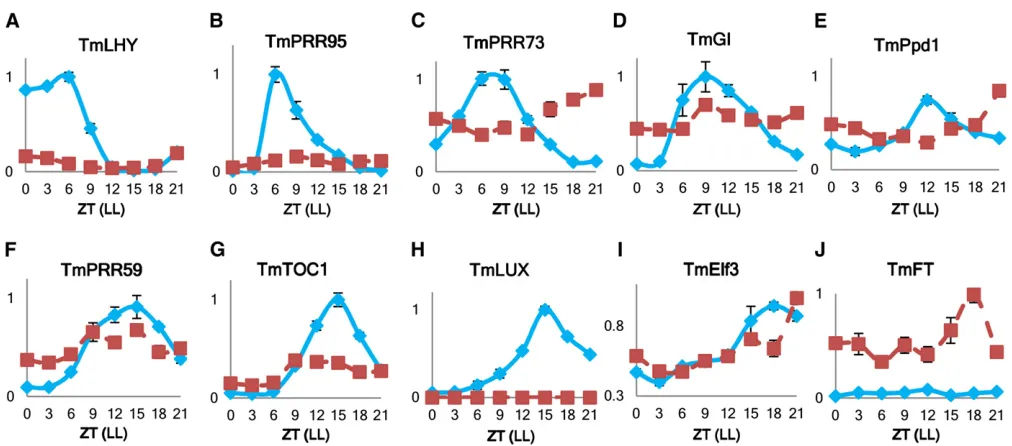
It had already been shown that KT3-5 had altered levels of circadian clock-related transcripts (Mizunoet al.2012); how-ever, the arrhythmia could not be detected because of the alternating sampling conditions in the previous study (Mizuno et al. 2012). To enable a proper comparison between KT3-5 andLUXmutants inArabidopsis(Hazenet al.2005; Onai and Ishiura 2005), the einkorn lines must be kept under constant ambient conditions,i.e., constant light and temperature. This is a basic prerequisite to analyze the circadian clock, as, according to its definition, it has to be maintained under constant envi-ronments (de Montaigu et al. 2010). In our study, einkorn wheat mutant KT3-5 showed a severely dampened amplitude and/or arrhythmia in transcript patterns of both morning (TmCCA1,TmPRR95,TmPRR73(Figure 4, A–C,File S9) and evening (TmGI, TmPpd1, TmPRR59, TmTOC1, TmLUX, and TmElf3) elements (Figure 4, D–I,File S9). As KT3-5 is a null mutation, it did not expressTmLUXat all (Figure S2and Figure 4H). However, consistent with an earlyflowering phenotype, KT3-5 displayed elevated levels ofTmFT—on average 10 times higher in KT3-5 than wt KT3-1 (Figure 4I).
To gain a complementary proof of function of TmLUX, we had already generated transgenicHvLUX-RNAi (RNA interfer-ence) plants in barley cv.‘Golden Promise.’We therefore took the opportunity to analyze clock gene expression in two of these transformants, DH_BG284E11-PP1 and BG353/1E15, which showed clear down-regulation ofHvLUXtranscripts com-pared to their azygous segregant BG284E11-38 (Figure S3D, File S9). In contrast to the KT3-5 mutant, the transcript abun-dances of HvCCA1, HvPRR95,HvGI, and HvLUX were not
arrhythmic (Figure S3). Transcript levels ofHvCCA1were slightly up-regulated in DH_BG284E11-PP1 and BG353/1E15, while those ofHvPRR95andHvGIwere quite strongly up-regulated (Figure S3,A–D). Furthermore, neither transformants exhibited an earlyflowering phenotype (Table S6,File S10).
Resequencing of LUX ortholog in 96 wheat accessions
To study sequence variation ofLUXin wheat, we selected 96 wild and cultivated accessions covering the majority of the observed phenotypic variation in flowering time and rese-quenced the putative LUXortholog (Figure S4A,File S11). While focusing on A and B genome copies of the gene (Gen-Bank accession nos. KF769443 and KF769444), which were mostly represented (Table S7), we found a higher number of haplotypes in wild wheat accessions—18vs.8 and 5vs.4 for A and B copy, respectively (Table S8,File S11,File S12, andFile S13). Because we found that the KT3-5 mutant was thermosensitive (Table 1), we also grouped the accessions according to the climatic conditions at the site where they had been collected (Figure 1A, Table 1). This grouping revealed more sequence variation being present in warmer climates within the A copy of LUX—16vs.12 (warm/cool) haplotypes and one haplotype more in the B copy, 7 vs. 6 (Table S9). This trend might suggest increased wheat LUX haplotype diversity in warmer climates. However, most hap-lotypes of the cultivated accessions came from the cooler climate (7vs.4), whereas most haplotypes of the wild acces-sions were collected from the warmer climate (14 vs. 9) (Table S10). Thus, higher sequence variation among the wild wheat accessions grown in warmer conditions, at least in part, might explain this observation (Table S8).
Another finding was the discovery of a TtLUX-A (A-genome homoeolog ofTtLUX) allele containing a seven-amino acid in-frame deletion within the MYB domain (Figure 2B, File S12). Importantly, a tetraploid wheat cultivar harboring this mutation,‘Tsing Hua no. 559,’headed among the early lines (May 31), while the range for the whole collection was from May 23 to June 30 (Figure S4A, File S11). The mu-tated TtLUX-Awas considered to be nonfunctional and the circadian clock of the cultivar was examined in a time-course RT-qPCR (Figure S4, B–E,File S9). Transcript levels of both TtLUX-AandTtLUX-B(B-genome homoeolog ofTtLUX) were up-regulated in‘Tsing Hua no. 559’compared to‘Fou Shou Figure 1 Phenotypic analysis of theEps-3Am locus. (A) Spikelet numbers per spike of wild-type KT3-1 (Triticum monococcumL.,) and mutant KT3-5 were plotted to show interactions with temperature. Presence of the interaction was confirmed with two-way factorial ANOVA at P , 0.0001. At 15°, KT3-1 developed significantly more spi-kelets than KT3-5 (P , 0.0000001, paired Student’s t-test). Seven plants per genotype and temperature treat-ment were analyzed. Error bars indicate SEM. (B) Frequency distribution of heading time among 38 F2 recombinants
and 38 randomly selected individuals grouped according to the presence/absence of theTmLUXgene. (C) Days to heading of F3families with closest recombination events were similarly determined by the genotypic status at theEps-3Amlocus; (a + b) plants with
mutant allele (solid) headed earlier than those carrying the wild-type allele (shaded). Error bars represent SEM.
Mai’(Figure S4B). As previous studies had shown that func-tionalAtLUXsuppresses its own expression (Hazenet al.2005; Onai and Ishiura 2005; Helferet al.2011), up-regulated tran-script levels ofTtLUXin‘Tsing Hua no. 559’may be a direct consequence of a nonfunctional TtLUX-A. Also, because the viable TtLUX-B most likely rescued the effect of mutated TtLUX-A, no clock distortion was observed (Figure S4, B–E). This idea is consistent with patterns of oscillation of other clock genes, TtGI, TtPRR95, and TtCCA1, which were very similar between both genotypes (Figure S4, C–E). However, we cannot completely rule out the possibility that a dysfunc-tional TtLUX-A has a partial phenotypic effect on flowering, which would be consistent with a role foreps genes infine tuningflowering in polyploid wheats (Snapeet al.2001).
Discussion
Mutants in LUX ARRHYTHMO have been associated with earlyflowering inArabidopsiswhere genetic and functional
studies allowed the gene to be placed within the evening loop of the circadian clock (Hazen et al. 2005; Onai and Ishiura 2005). LUX participated in forming an EC together with two other proteins, ELF3 and ELF4 (Nusinow et al. 2011). Recently,WPCL1, a gene homologous toArabidopsis PCL1/LUX was proposed as a candidate conferring early heading in einkorn wheat mutant KT3-5 (Mizuno et al. 2012). The prediction was based on high-density QTL map-ping and synteny with barley genome (Mizunoet al.2012). Preliminary results showed that a possible deletion of WPCL1 could be responsible for the observed phenotype in KT3-5 (Mizunoet al.2012). Within the present study, we found thatWPCL1is the most sensible candidate forEps-3Am
containing KT3-5. By usingfine mapping, we also delimited the locus to only two genes when compared to the syntenic region in the barley genome. This included putative TmPCL1/TmLUXandTmPUMILIO(Figure 2A). We performed TILLING of both genes; however, only for TmPUMILIOdid we obtain one mutation potentially affecting gene function Figure 2 Comparative analysis of theEps-3Amlocus. (A) Genetically mappedEps-3Amlocus (3Am) was integrated with physical ctg_95 from barley chromosome 3H and two contigs, ctg_1331 and ctg_1512 from bread wheat chromosome 3A. Wheat contigs did not overlap (gap). Only BAC addresses selected for the minimum tiling path (MTP) are shown. Putative genes (color-coded) annotated from the barley and wheat sequence could be mapped in F2population of einkorn wheat, revealing higher synteny with barley. New recombinations found in the F2/F3population (rectanglesfilled
with number of recombinations) delimited theEps-3Amlocus to only two genes:TmLUX ARRHYTHMO(TmLUX, red) andTmPUMILIO(orange). Both genes had been deleted from the genomes of the early headingT. monococcumKT mutants. Genes mapped proximally to the locus (KIN,HMA, and LEG) did not have putative orthologs in the collinear part of rice chr1 (Os01) and the syntenic relationship was thus broken at this site. RFP, a putative ringfinger protein and ZT, a putative zinc transporterflanked theEps-3Amlocus from the distal site. A marker developed based on the putative transcription factor (BTB/POZ) could not be linked with the locus in einkorn wheat. (B) Amino acid sequence of the MYB domain from LUX was conserved between wheat and barley. Resequencing ofLUXin selected germplasm revealed deletion and substitution mutants at a highly conserved nucleic acid binding motif‘SHxQK’(Hwanget al.2002)‘Tsing Hua no. 559’inT. turgidumL. (A genome copy ofLUX), and‘Super Precoz 2H’(eam10) in barley (Campoliet al.2013), respectively.
(Figure S1). Although as expected, we did not observe an early flowering phenotype in those plants, the obtained re-sult needs to be further verified by introgressing the muta-tion into the nonmutant background to rule out the possible masking effect of other genome aberrations. Such an ap-proach has already been successfully applied in barley mu-tant research (Drukaet al.2011).
Previously discovered differential regulation of clock-related transcripts allowed us to hypothesize that earlyfl ower-ing in KT3-5 was due to a disrupted circadian clock (Mizuno
et al.2012). However, in the previous study, plants were only analyzed under short day conditions, which is insufficient to fully explain the circadian clock distortion in KT3-5 (Mizuno et al.2012). In the present study, we kept the plants in a con-stant environment,i.e., constant light and temperature, and this revealed arrhythmia in transcript patterns of clock-re-lated genes (Figure 4). We thus demonstrated that the KT3-5 mutant is very similar toeam10as both are probably caused by knock-out of the orthologous gene PCL1/LUX (Campoliet al.2013).
HvLUX-RNAi plants generated in our study helped us to confirm the position of HvLUX in the circadian clock net-work. In agreement with expectations, we observed higher transcript levels of evening elementsHvPRR95andHvGIin these plants (Figure S3, B and C). This suggests the repres-sive action of HvLUX by direct binding to theHvPRR9 pro-moter as already well investigated between AtLUX and AtPRR9inArabidopsis(Helferet al.2011). Besides, the ef-fect of HvLUX knock-down on HvLHY transcript level was much smaller, probably due to the missing direct interaction between these elements (Figure S3A) (Pokhilkoet al.2012). However, slight up-regulation ofHvLHYtranscript is against prediction from the most recent clock model, in which PRR9 and PRR7 proteins down-regulate LHY (Pokhilko et al. 2012). On the other hand, the reciprocal positive effect of LHY onPRR9may be responsible for the observed low tran-script levels of TmPRR95 in KT3-5 (Figure 4B), (Pokhilko et al.2012). We hypothesize that under constant conditions and complete absence of TmLUX action, the level of TmPRR95 protein initially rises and leads to severely dampened LHY transcript level through the negative feedback. However, low TmLHY protein levels cannot fulfill the required activating effect on theTmPRR95and high transcript levels ofTmPRR95 cannot be maintained, resulting in becoming low and arrhythmic (Figure 4B).
It has been suggested that the early heading of KT3-5 was driven by elevatedPpd1expression and subsequent accumu-lation of WFT transcript (Mizuno et al. 2012). Evidence found by Campoli et al.(2013) also supports that HvLUX1 (barley ortholog of PCL1/LUX) acts onHvFT through Ppd-H1, as it shows a clear interaction with this gene,e.g.,eam10 mutant lines harboring the wild-typePpd-H1alleleflowered earlier than those with the recessive ppd-H1 mutant allele (Campoli et al. 2013). In our study, we also observed ele-vated transcript level of TmPpd-1in KT3-5 during the sub-jective night/dawn period (Figure 4E) and in turn higher TmFT/TmVrn3 transcript levels (Figure 4J). In addition, our transgenic HvLUX-RNAi plants were generated in ‘Golden Promise,’which possesses theppd-H1mutant allele, i.e., photoperiod insensitivity (Faure et al. 2012), and this may be another decisive factor for the missing earlyfl ower-ing phenotype in our transgenic plants (Table S6). Hence, we conclude that HvLUX knock-down is not sufficient to abolish the circadian rhythm under constant environment and to induce an earlyflowering phenotype in photoperiod insensitive barleycv.‘Golden Promise.’
Figure 3 Delayedfluorescence (DF) measurements were performed to compare wild-type and mutant circadian clock outputs to the chloroplast. Oscillation of the DF detected in the wild-type KT3-1 (A) (T. monococcum L.) lasted longer than that of the mutant KT3-5 (B). Leaf samples were kept under 22°in the darkness interrupted every hour with a light pulse lasting for 1 min prior to capturing the picture. Light intensities were extracted and normalized and trends for the DF curves were removed. Error bars indicate standard deviations for 16 replicates. (C and D) Relative amplitude errors (RAEs) of the DF oscillation plotted against the periods generated from the BRASS software. Periods were calculated for the DF from 36 to 132 hr. Each data point represents the time course measure-ment at a single region on the leaf from wild-type KT3-1 or mutant KT3-5 plants. Experiments were performed in two different temperature regimes: 22°(C) and 17°(D). In both cases, wild-type samples were more tightly clustered around the expected value of the period equaling 24 hr. (C) At 22°, mutant KT3-5 showed greater period lengthening than KT3-1, despite the higher RAE values found in both lines. The temperature equaling 22°was thus an appropriate condition to recognize a mutant clock phenotype.
Disruptive mutations in cerealLUXseem to be favorable for spring cultivars that are dedicated for late sowing. In Japan, mutant KT3-5 performed better than KT3-1 as it could escape the humid and hot summer season and reach full maturity due to earlierflowering (Utsugiet al.2006). In the case of winter cultivars, a functional LUX seems to be required to provide full yield potential, a requirement con-sistent with low temperature treatment during vegetative growth, which significantly increased yield of wild-type plants. We speculate that the low photoperiodic require-ment of cereal LUX mutants could also be advantageous for a better adaptation to shorter growing seasons, shorter photoperiods, and warmer climates. This makes it very sim-ilar to HvELF3mutations (Faureet al.2012; Zakhrabekova et al.2012).
It is thought that theepstrait is connected with adverse effects on spike size and decreased plant yield (Lewiset al. 2008). We observed a similar relationship in the KT3-5 mu-tant, but only in the cooler temperature regime (15°). We found that spike development also depends on temperature, with the most temperature-sensitive period being the early phase of development. We observed much greater pheno-typic variation in spikelet number per spike in KT3-1 (wt) than in KT3-5 when grown in different environmental regimes, indicating that KT3-5 has a decreased phenotypic plasticity.
Consistent with thisfinding that at warmer temperatures KT3-5 mutant performed better than the donor wild-type KT3-1, we hypothesizedfinding higher sequence variation in LUXin warmer climates. We in fact observed this association in wheat within the sequence of the A-genome copy ofLUX.
Besides, we found one putative nonfunctional mutation in the Chinese cultivar ‘Tsing Hua no. 559.’This cultivar was initially classified as being from a cooler climate, although the average temperature value was just below the chosen threshold (17.5°). Apart from this allele, we did notfind any yet known functional variation in wheat homoeologs ofLUX, which generally is very similar to the situation among barley accessions (Campoliet al.2013). Future studies need to be carried out to further test the relationship of an adaptive advantage between temperature and circadian clock in tem-perate cereals.
In this study onLUX ARRHYTHMOin einkorn wheat, we have provided further justification for circadian clock re-search in modern crops and a clear link to environmental adaptation and yield optimization. We have also shown that the most efficient route to follow can be screening for clock mutants by using the DF measurements and subsequently confirming putative mutations with the time course RT-qPCR. Further investigations are required, including those at the protein level, to bring more direct evidence for the existence and molecular function of an EC in cereal crops.
Acknowledgments
We appreciate help from Hana Simkova, Institute of Ex-perimental Botany (Olomouc, Czech Republic), for sharing the 3AL, 3B, and 3DL DNA samples. We thank Robbie Waugh, James Hutton Institute (Dundee, United Kingdom), for critically reading previous versions of the manuscript. We acknowledge the help of Susanne König, Ravi Koppolu, Marzena Kurowska, Dimitar Douchkov, Patrick Schweizer, Figure 4 Relative transcript levels of circadian clock genes in einkorn wheat. (A)TmCCA1, (B)TmPRR95, (C)TmPRR73, (D) TmGI, (E)TmPpd1(F) TmPRR59, (G)TmTOC1, (H)TmLUX, (I)TmElf3, and (J)TmFTwere measured from a 1-day time-course RT-qPCR study on wild-type KT3-1 (T. mono-coccumL., blue diamonds) and early heading mutant KT3-5 (T. monococcumL., red squares). Sampling was performed on 31-day-old plants starting at 6:00AM(ZT = 0) of the second day under constant light and temperature. Leaves were harvested every 3 hr (LL conditions) from at least three plants per time point per genotype (biological replicates). Error bars indicate SEM.
Andrew J. Millar, Anna A. Filatenko, and excellent techni-cal assistance from Kathrin Gramel-Eikenroth, Enk Geyer, Corinna Trautewig, Jelena Perovic, Anne Kusserow, Ulrike Beier, Carola Bollmann, Sabine Sommerfeld, Andrea Müller, Ingrid Otto, Ute Krajewsky, Heike Harms, Christiane Kehler, Marita Nix, Birgit Dubsky, and Kerstin Wolf. This work was supported by grants from the US Department of Agri-culture National Institute of Food and AgriAgri-culture grant 2008-35300-04588 to BSG; German Research Foundation (Deutsche Forschungsgemeinschaft, DFG) Priority Programme SPP1530 to BK; BARLEX grant no. 0314000 to NS; and DFG grant no. SCHN 768/3-1 and the German Federal Ministry of Education and Research GABI-FUTURE Start Program grant no. 0315071 to TS.
Literature Cited
Börner, A., G. H. Buck-Sorlin, P. M. Hayes, S. Malyshev, and V. Korzun, 2002 Molecular mapping of major genes and quanti-tative trait loci determiningflowering time in response to pho-toperiod in barley. Plant Breed. 121: 129–132.
Campoli, C., M. Shtaya, S. Davis, and M. von Korff, 2012 Expression conservation within the circadian clock of a monocot: natural variation at barleyPpd-H1affects circadian expression of flowering time genes, but not clock orthologs. BMC Plant Biol. 12: 97.
Campoli, C., A. Pankin, B. Drosse, C. M. Casao, S. J. Daviset al., 2013 HvLUX1is a candidate gene underlying theearly matu-rity 10 locus in barley: phylogeny, diversity, and interactions with the circadian clock and photoperiodic pathways. New Phy-tol. 199: 1045–1059.
de Montaigu, A., R. Toth, and G. Coupland, 2010 Plant develop-ment goes like clockwork. Trends Genet. 26: 296–306. Dixon, L. E., K. Knox, L. Kozma-Bognar, M. M. Southern, A. Pokhilko
et al., 2011 Temporal repression of core circadian genes is me-diated through EARLY FLOWERING 3 in Arabidopsis. Curr. Biol. 21: 120–125.
Dodd, A. N., N. Salathia, A. Hall, E. Kevei, R. Toth et al., 2005 Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science 309: 630–633. Doyle, J. J., and J. L. Doyle, 1987 A rapid DNA isolation
proce-dure for small quantities of fresh leaf tissue. Phytochem. Bull. 19: 11–15.
Druka, A., J. Franckowiak, U. Lundqvist, N. Bonar, J. Alexander
et al., 2011 Genetic dissection of barley morphology and de-velopment. Plant Physiol. 155: 617–627.
Faure, S., A. S. Turner, D. Gruszka, V. Christodoulou, S. J. Daviset al., 2012 Mutation at the circadian clock gene EARLY MATURITY 8 adapts domesticated barley (Hordeum vulgare) to short growing seasons. Proc. Natl. Acad. Sci. USA 109: 8328–8333.
Gallagher, L. W., K. M. Soliman, and H. Vivar, 1991 Interactions among loci conferring photoperiod insensitivity for heading time in spring barley. Crop Sci. 31: 256–261.
Gawron´ski, P., and T. Schnurbusch, 2012 High-density mapping of theearliness per se 3Am(Eps-3Am) locus in diploid einkorn wheat and its relation to the syntenic regions in rice and Bra-chypodium distachyonL. Mol. Breed. 30: 1097–1108.
Gould, P. D., P. Diaz, C. Hogben, J. Kusakina, R. Salem et al., 2009 Delayedfluorescence as a universal tool for the measure-ment of circadian rhythms in higher plants. Plant J. 58: 893–901. Hazen, S. P., T. F. Schultz, J. L. Pruneda-Paz, J. O. Borevitz, J. R. Eckeret al., 2005 LUX ARRHYTHMO encodes a Myb domain
protein essential for circadian rhythms. Proc. Natl. Acad. Sci. USA 102: 10387–10392.
Helfer, A., D. A. Nusinow, B. Y. Chow, A. R. Gehrke, M. L. Bulyk
et al., 2011 LUX ARRHYTHMO encodes a nighttime repressor of circadian gene expression in the Arabidopsis core clock. Curr. Biol. 21: 126–133.
Hicks, K. A., T. M. Albertson, and D. R. Wagner, 2001 EARLY FLOWERING 3 encodes a novel protein that regulates circadian clock function and flowering in Arabidopsis. Plant Cell 13: 1281–1292.
Hwang, D., H. C. Chen, and J. Sheen, 2002 Two-component sig-nal transduction pathways in Arabidopsis. Plant Physiol. 129: 500–515.
Imaizumi, T., 2009 Arabidopsis circadian clock and photoperiod-ism: time to think about location. Curr. Opin. Plant Biol. 13: 83– 89.
Lewis, S., M. E. Faricelli, M. L. Appendino, M. Valarik, and J. Dub-covsky, 2008 The chromosome region including the earliness per se locusEps-Am1affects the duration of early developmental phases and spikelet number in diploid wheat. J. Exp. Bot. 59: 3595–3607.
Mayer, K. F. X., R. Waugh, P. Langridge, T. J. Close, R. P. Wiseet al., 2012 A physical, genetic and functional sequence assembly of the barley genome. Nature 491: 711–716.
Mizuno, N., M. Nitta, K. Sato, and S. Nasuda, 2012 A wheat ho-mologue ofPHYTOCLOCK 1is a candidate gene conferring the early heading phenotype to einkorn wheat. Genes Genet. Syst. 87: 357–367.
Nusinow, D. A., A. Helfer, E. E. Hamilton, J. J. King, T. Imaizumi
et al., 2011 The ELF4–ELF3-LUX complex links the circadian clock to diurnal control of hypocotyl growth. Nature 475: 398– 402.
Onai, K., and M. Ishiura, 2005 PHYTOCLOCK 1 encoding a novel GARP protein essential for the Arabidopsis circadian clock. Genes Cells 10: 963–972.
Pittendrigh, C. S., 1993 Temporal organization: reflections of a Darwinian clock-watcher. Annu. Rev. Physiol. 55: 17–54. Plautz, J. D., M. Straume, R. Stanewsky, C. F. Jamison, C. Brandes
et al., 1997 Quantitative analysis of Drosophila period gene transcription in living animals. J. Biol. Rhythms 12: 204–217. Pokhilko, A., A. P. Fernandez, K. D. Edwards, M. M. Southern, K. J.
Hallidayet al., 2012 The clock gene circuit in Arabidopsis in-cludes a repressilator with additional feedback loops. Mol. Syst. Biol. 8: 574.
Rutherford, A. W., Govindjee, and Y. Inoue, 1984 Charge accu-mulation and photochemistry in leaves studied by thermo-lumi-nescence and delayed light-emission. Proc. Natl. Acad. Sci. USA 81: 1107–1111.
Shindo, C., and T. Sasakuma, 2001 Early heading mutants ofT. monococcum and Ae. squarrosa, A- and D-genome ancestral spe-cies of hexaploid wheat. Breed. Sci. 51: 95–98.
Snape, J. W., R. Sarma, S. A. Quarrie, L. Fish, G. Galiba et al., 2001 Mapping genes for flowering time and frost tolerance in cereals using precise genetic stocks. Euphytica 120: 309–315. Strehler, B. L., and W. Arnold, 1951 Light production by green
plants. J. Gen. Physiol. 34: 809–820.
Utsugi, S., M. Maekawa, and K. Noda, 2006 An efficient transient gene expression system using aleurones of diploid wheat seeds. Plant Biotechnol. 23: 413–417.
Zakhrabekova, S., S. P. Gough, I. Braumann, A. H. Muller, J. Lundqvist et al., 2012 Induced mutations in circadian clock regulator Mat-a facilitated short-season adaptation and range extension in cultivated barley. Proc. Natl. Acad. Sci. USA 109: 4326–4331.
Communicating editor: J. Borevitz
GENETICS
Supporting Information
http://www.genetics.org/lookup/suppl/doi:10.1534/genetics.113.158444/-/DC1
A Distorted Circadian Clock Causes Early Flowering
and Temperature-Dependent Variation in Spike
Development in the
Eps-3A
mMutant of
Einkorn Wheat
Piotr Gawron´ski, Ruvini Ariyadasa, Axel Himmelbach, Naser Poursarebani, Benjamin Kilian, Nils Stein, Burkhard Steuernagel, Goetz Hensel, Jochen Kumlehn, Sunish Kumar Sehgal, Bikram S. Gill, Peter Gould, Anthony Hall, and Thorsten Schnurbusch
2 SI P. Gawroński et al.
P. Gawroński et al. 3 SI
4 SI P. Gawroński et al.
Figure S3 Relative transcript levels of circadian clock genes in transgenic barley. (A) HvCCA1, (B) HvPRR95, (C) HvGI and (D) HvLUX, were measured from a one‐day time‐course RT‐qPCR study on azygous BG284E11‐38 (‘Golden Promise’, H. vulgare L., brown diamonds), HvLUX‐RNAi plants in ‘Golden Promise’ DH BG284E11‐PP1 (orange
P. Gawroński et al. 5 SI
Figure S4 Allele mining in a diverse wheat collection revealed a putative mutant at TtLUX‐A in cultivar ‘Tsing Hua no. 559’. (A) ‘Tsing Hua no. 559’ (T. turgidum L.) was a relatively early heading (red arrow) genotype which flowered one week earlier than another Chinese cultivar ‘Fo Shou Mai’ (T. turgidum L., blue arrow), two plants per accession were evaluated in the field at IPK in Gatersleben in 2011. Deletion involving seven AA from the MYB domain of TtLUX‐A could potentially impair function of the protein, Figure 2B. (B) Genome‐specific primers were used to amplify both copies of TtLUX. In ‘Tsing Hua no. 559’, transcripts of TtLUX‐A (A‐genome copy of LUX, red squares) and TtLUX‐B (B‐genome copy of LUX, black triangles) were up‐regulated when compared to ‘Fo Shou Mai’ (TtLUX‐A,
6 SI P. Gawroński et al.
Table S1 List of primers
Name Comment or enzyme or amplicon size [nt] Forward primer (5'‐3') Reverse primer (5'‐3')
INDEL_271_272 a size polymorphism CAAGGCTCTGCAGTACTTGACAGAG GACTTCATCAAGCGTAAGCACATGTC
CAPS_321_322 a Hpy188I TTGCCGCTTAAGAATAAGCATCCTC GTTTCCATCATAGGGGAAGTTTGTG
CAPS_325_326 a Hpy188I TTTCATTGTGCACCACCAACC CAGCCGTGGTGCTCTTCTTCT
CAPS_331_332 a BamHI GCTACTCTGACTCCGCCAAGG AGCCCGACCTCCTTCACCA
PAV_261_262 a presence/absence CTCTACGTCAAGCGGATGCAG GTGGTAGTGCGGGTAGGAGGAG
PAV_295_296 a presence/absence CATACTGGTCTGTAGCAAGCAAGCA AGCACGGCTCAGATAAAGGAGTTG
CAPS_341_342 a EcoNI CGAGAGCATGTCCGACTTC ATATTCCCCAGTGTGTGATGC
CAPS_zt3_4 a BstUI GAGGCATGGAGGAACCTATGGAC AACTTGAAGATGGGCTTGTCGAA
CAPS_335_336 a MspI CAGTGATGCAGGCGTGGAG ATGGCCATGTCGATGTACGG
CAPS_311_312 a MvaI TCGTAATAAGATGTGCGGAGAGATG CAATGAACCAACTGTTGCATTTTCA
TP_13_14 a external primers CAAGCCTGACCCAGTACTCAGG CACAGCCTCCTCCCACAGTTC
TP_13_14 a internal primers ATGCGACTTCGGTTGATGCTC ATTCACCACATTAGCAACGCA
AS_PCR_17_18 a additional mismatch ACATCTGAAGAGCCAAGTTGATCC GCTATCACATACCGCAAGAGAATCTC
INDEL_201_202 a size polymorphism GACAAATGTAGTGTTCATGGGGATG TTCCAGAGGATATGCCTTTGCACTTGG
PAV_365_366 a presence/absence TCCAGATGGGAATGTGTCGTCT GCCATAGCTTTGTTGTGGATGC
CAPS_359_360 a NotI AGTTTGCAAGAAAAACAGCACAGG AGAAACAGGCCCGAAAACAAGTC
TmCCA1, HvCCA1, TtCCA1 b 150 CAAGGTCTTCTCCCTCTTTTTGCTC GTTGACCTTGCTCCTGAGCTACTTG
P. Gawroński et al. 7 SI
Name Comment or enzyme or amplicon size [nt] Forward primer (5'‐3') Reverse primer (5'‐3')
TmUBC, HvUBC, TtUBC b 151 AAGCAGCCAGAATGTACAGCGAGAAC GGTACAGACCAGCAAAGCCAGAAATG
HvLUX b 136 GGTGACCGAGTGGGAGACG GCGGTGCACGTCCAGAAG
TmGI, HvGI, TtGI b 135 TCCATGACAAAGTAGGTGGCTGA CATGGTCCTGATGTTGAGTGGAG
TmPRR95, HvPRR95, TtPRR95 b 106 GTCTGGGTTCCTCCTACTCTCCAC GCATTTGAAAACCATGCTAACTGC
TmPpd1 b 129 AATGGTGCTGCTGACTTCTTGG CTTCTGCGTCTGAATGGCACTT
TmFT b 153 GCAGGAGGTGATGTGCTACGAG AGGTTGTAGAGCTCGGCGAAGT
TmPRR73 b 169 AAGGGTCTTGCTGGTGGAGAAC AGGCATGGCGACCTCAGTTAAT
TmPRR59 b 145 AAGAAAATCAGCGGTTGCATCA GTTTCACCGCTCACAGTCCATC
TmTOC1 b 111 AGGAGACGGATGCTTGGTTTG GTCCGAAAGGAGTGTGGTGCT
TmElf3 b 100 GAGTGGGCCTACGGTGTCCAA AGAGTCCCTTGATCTCGTGTCCG
TtLUX‐A b 199 GAGTTAGCCCGGCAGGTAACAAC CCCTCCCCCGAAGTCAAAAC
TtLUX‐B b 131 GATTGGTGTTGCGAGGTTCG CGTCTCCCACTCCGTCACC
TmPUMILIO b 174 AGCGACTTCTCCACACTCCTCAG AATATTTAACCACACGCCGCAAC
TILLINGHvLUX c 1177 GGTGACCGAGTGGGAGACG ATCCTAATTCCCTTGTTGGGCTTC
TILLINGHvPUM c 1593 CAGGCAATCAAAACAATGAGTCG CGTACATTTGAAGGCTGAGTGCTAA
TILLINGHvPUM c ‐ ACCCACTCTATGCTCAATTTCTTCG GCTTCTGTGACTGAACCAATGGA
LUX‐A d Variable GAGTTAGCCCGGCAGGTAACAAC ACAGAGCACACACTCTGCAACTCTC
LUX‐B d Variable TCCAATCCGTCCAATCCAATC TTGACTGATCGACACAAACACAC
8 SI P. Gawroński et al.
a Markers mapped in F2/F3 population. Order presented reflects the real genetic order of mapped loci. Markers PAV_365_366 and CAPS_359_360 could not be genetically linked to the locus b Primer sequences used for RT‐qPCR experiments
c Primer sequences used for TILLING
P. Gawroński et al. 9 SI
Table S2 Gene content in the physical contig_95 from barley and contigs 1331 and 1512 from bread wheat syntenic to the Eps‐3Am locus
MTP BAC Putative genes/putative rice ortholog
Putative function (BlastX) Abbreviation corresponding to the Figure 2
Genetic marker
contig_95
HVVMRXALLmA0519N21 Os03g0819400 heavy metal associated protein HMA CAPS_325_326
HVVMRXALLmA0519N21 Os01g0976200 legumin like LEG CAPS_331_332
HVVMRXALLmA0519N21 Os02g0472700 ser/treo kinase KIN CAPS_321_322
HVVMRXALLmA0519N21 Os03g0819400 truncated heavy metal associated protein 2 HMA_trunc no
HVVMRXALLeA0205E03 Os03g0819400 heavy metal associated protein HMA CAPS_325_326
HVVMRXALLeA0205E03 Os01g0976200 legumin like LEG CAPS_331_332
HVVMRXALLeA0205E03 Os01g0971800 similar to AtLUX ARRHYTHMO LUX PAV_261_262
HVVMRXALLeA0205E03 Os03g0819400 truncated heavy metal associated protein 2 HMA_trunc no
HVVMRXALLhA0201J07 Os01g0971800 similar to AtLUX ARRHYTHMO LUX PAV_261_262
HVVMRXALLrA0045I17 Os01g0971900 similar to AtPUMILIO 1 PUM PAV_295_296
HVVMRXALLeA0121L04 Os01g0971900 similar to AtPUMILIO 1 PUM PAV_295_296
HVVMRXALLeA0121L04 Os01g0972200 zinc transporter ZT CAPS_zt3_4
HVVMRXALLeA0121L04 Os05g0110000 ring finger protein RFP CAPS_341_342
HVVMRXALLmA0021I14 Os01g0972200 zinc transporter ZT CAPS_zt3_4
HVVMRXALLmA0021I14 Os05g0110000 ring finger protein RFP CAPS_341_342
10 SI P. Gawroński et al.
MTP BAC Putative genes/putative rice ortholog
Putative function (BlastX) Abbreviation corresponding to the Figure 2
Genetic marker
HVVMRXALLeA0274H04 no
HVVMRXALLrA0172M23 Os01g0972300 rotundifolia like/transposon RTF no
HVVMRXALLmA0521P17 Os01g0972300 rotundifolia like/transposon RTF no
HVVMRXALLmA0477A13 Os01g0972800 WRKY1‐like WRKY CAPS_335_336
HVVMRXALLmA0074K19 Os01g0972900 WD repeat cell cycle switch WDR no
HVVMRXALLmA0074K19 Os01g0972800 WRKY1‐like WRKY CAPS_335_336
HVVMRXALLeA0320E22 Os01g0972800 WRKY1‐like WRKY CAPS_335_336
HVVMRXALLeA0320E22 Os01g0972900 WD repeat cell cycle switch WDR no
HVVMRXALLmA0295O08 Os09g0330700 ankyrin‐like ANK no
HVVMRXALLmA0295O08 Os12g0512800 cytochrome P450 CYT_A no
HVVMRXALLeA0208F01 Os09g0330700 ankyrin‐like ANK no
HVVMRXALLeA0208F01 Os12g0512800 cytochrome P450 CYT_A no
HVVMRXALLeA0208F01 Os12g0512800 cytochrome P450 CYT_B no
HVVMRXALLmA0133D05 Os08g0340900 PPR‐repeat containing PPR no
P. Gawroński et al. 11 SI
MTP BAC Putative genes/putative rice ortholog
Putative function (BlastX) Abbreviation corresponding to the Figure 2
Genetic marker
HVVMRXALLmA0133D05 Os12g0512800 cytochrome P450 CYT_A no
HVVMRXALLmA0133D05 Os12g0512800 cytochrome P450 CYT_B no
HVVMRXALLmA0133D05 Os12g0512800 cytochrome P450 CYT_C no
HVVMRXALLmA0133D05 Os12g0512800 cytochrome P450 CYT_D no
HVVMRXALLmA0133D05 Os03g0274300 telomere binding prot TBP TBP CAPS_311_312
HVVMRXALLrA0088H17 Os01g0973000 hydrolase HYD TP_13_14
HVVMRXALLrA0088H17 Os12g0512800 cytochrome P450 CYT_D no
HVVMRXALLrA0088H17 Os03g0274300 telomere binding prot TBP TBP CAPS_311_312
HVVMRXALLrA0088H17 Os08g0340900 PPR‐repeat containing PPR no
contig_1331
3ALhA_0186G01 Os02g0472700 ser/treo kinase KIN CAPS_321_322
3ALhA_0173M19 Os02g0472700 ser/treo kinase KIN CAPS_321_322
3ALhA_0173M19 Os03g0819400 heavy metal associated protein HMA CAPS_325_326
3ALhA_0101H11 Os03g0819400 truncated heavy metal associated protein 2 no no
3ALhA_0101H11 Os03g0819400 heavy metal associated protein HMA CAPS_325_326
3ALhA_0101H11 Os01g0976200 legumin like LEG CAPS_331_332
3ALhA_0101H11 Os01g0976200 truncated legumin like 2 no no
3ALhA_0101H11 Os09g0538750 truncated bZIP, proline rich no no
3ALhA_0159B08 Os09g0538750 truncated bZIP, proline rich no no
12 SI P. Gawroński et al.
MTP BAC Putative genes/putative rice ortholog
Putative function (BlastX) Abbreviation corresponding to the Figure 2
Genetic marker
3ALhA_0159B08 Os01g0971800 similar to AtLUX ARRHYTHMO LUX PAV_261_262
3ALhA_0046O18 Os01g0971800 similar to AtLUX ARRHYTHMO LUX PAV_261_262
3ALhA_0123J12 no no no no
3ALhA_0194A07 no no no no
3ALhA_0150I20 Os01g0971900 similar to AtPUMILIO 1 PUM PAV_295_296
3ALhA_0150I20 no TAA1
3ALhA_0150I20 no RRM
3ALhA_0150I20 no TAA2
3ALhA_0072H09 no ATP‐ase no no
3ALhA_0072H09 no TAA1 no no
3ALhA_0072H09 no RRM no PAV_365_366, not
linked
3ALhA_0072H09 no TAA2 no no
3ALhA_0208L01 no RRM no PAV_365_366, not
linked
3ALhA_0208L01 no TAA2
3ALhA_0086P11 Os11g0622600 BTB/POZ containing TF BTB/POZ CAPS_359_360, not
linked
P. Gawroński et al. 13 SI
MTP BAC Putative genes/putative rice ortholog
Putative function (BlastX) Abbreviation corresponding to the Figure 2
Genetic marker
contig_1512
3ALhA_0089I03 Os01g0972200 zinc transporter ZT CAPS_zt3_4
Os05g0110000 RING‐H2 finger protein RFP CAPS_341_342
no AP2 domain contaning TF no no
3ALhA_0140J06 Os01g0972200 zinc transporter ZT CAPS_zt3_4
Os05g0110000 RING‐H2 finger protein RFP CAPS_341_342
no AP2 domain contaning TF no no
3AlhA_0156I13 No no no no
14 SI P. Gawroński et al.
Table S3 KT lines of einkorn wheat analyzed
No. Accession No. Description based on KIBR b
1 KT001‐002 T. boeoticum ssp. boeoticum
2 KT001‐003 T. boeoticum ssp. boeoticum
3 KT002‐001 T. urartu variety nigrum 4 KT003‐002 T. monococcum variety vulgare 5 KT003‐003 T. monococcum variety flavescens 6 KT003‐004 T. monococcum variety hornemanni 7 KT003‐038 T. monococcum strain KUS 68
8 KT003‐039 T. monococcum strain KUS 82
9 KT003‐006 T. monococcum strain vulgare spiral mutant
10 KT003‐007 T. monococcum strain vulgare old rose (hetero) mutant 11 KT003‐008 T. monococcum strain vulgare light green mutant 12 KT003‐009 T. monococcum strain vulgare orange mutant 13 KT003‐010 a T. monococcum strain vulgare ej., non–glossy mutant
14 KT003‐011 a T. monococcum strain vulgare early, spiral mutant
15 KT003‐012 a T. monococcum strain vulgare early, green mutant
16 KT003‐013 a T. monococcum strain vulgare ej., glossy (stripe) mutant
17 KT003‐014 a T. monococcum strain vulgare albino mutant
18 KT003‐015 a T. monococcum strain vulgare pigmy mutant
19 KT003‐016 a T. monococcum strain vulgare pigmy, narrow leaf mutant
20 KT003‐017 a T. monococcum strain vulgare purple red mutant
21 KT003‐019 T. monococcum strain vulgare nh. mutant 22 KT003‐020 T. monococcum strain vulgare sg. mutant 23 KT003‐021 T. monococcum strain vulgare moegi mutant 24 KT003‐022 T. monococcum strain vulgare dwarf mutant 25 KT003‐023 T. monococcum strain vulgare black glume mutant 26 KT003‐024 T. monococcum strain vulgare branched spike mutant 27 KT003‐025 T. monococcum strain vulgare soft spike mutant 28 KT003‐026 T. monococcum strain vulgare wrapped glume mutant 29 KT003‐027 a T. monococcum strain vulgare rolled leaf mutant
30 KT003‐028 a T. monococcum strain vulgare rolled leaf, dwarf mutant
31 KT003‐029 a T. monococcum strain vulgare early, snith mutant
32 KT003‐030 a T. monococcum strain early Translocation aT1
33 KT003‐031 a T. monococcum strain early, male sg. Translocation aT1
34 KT003‐032 T. monococcum strain Translocation aT2 35 KT003‐033 a T. monococcum strain Translocation aT5
36 KT003‐034 a T. monococcum strain Translocation mT6
37 KT003‐035 a T. monococcum strain Translocation aT7
38 KT003‐036 T. monococcum strain Translocation mT9 39 KT003‐037 T. monococcum strain haploid inducer
a Sixteen lines marked in italics share a deletion at the Eps‐3Am locus and were selected for further analysis,
however KT003‐017 did not recover after vernalization and phenotypic scores are not presented
b KIBR= Kihara Institute of Biological Research, more information can be obtained at
P. Gawroński et al. 15 SI
Table S4 Haplotype marker analysis performed on 15 KT mutants at the Eps‐3Am locus
Marker
haplotype eps mutant CAPS_23_24 CAPS_37_38 CAPS_131_132 CAPS_341_342
PAV_261_262
PAV_295_296 CAPS_331_332
Heading date difference according
to KT3‐1;
P value for heading date
difference
1 KT003‐013_mut A A A + ‐ + ‐45.17 5.11*10‐6
1 KT003‐014_mut A A A + ‐ + ‐36 0.014
1 KT003‐015_mut A A A + ‐ + ‐32.61 1.05*10‐5
1 KT003‐016_mut A A A + ‐ + ‐32.17 2.7*10‐7
1 KT003‐027_mut A A A + ‐ + ‐33 5.87*10‐6
1 KT003‐028_mut A A A + ‐ + ‐30.42 0.0015
1 KT003‐029_mut A A A + ‐ + ‐39 5.28*10‐5
1 KT003‐030_mut A A A + ‐ + ‐32 2.68*10‐6
1 KT003‐031_mut A A A + ‐ + ‐31.72 4.03*10‐7
1 KT003‐033_mut A A A + ‐ + ‐25.5 0.11
1 KT003‐034_mut A A A + ‐ + ‐31.17 2.81*10‐7
1 KT003‐035_mut A A A + ‐ + ‐54.11 1.6*10‐7
2 KT003‐010_mut B A A ‐ ‐ + ‐60.25 3.89*10‐6
3 KT003‐011_mut B A A + ‐ + ‐52.5 4.5*10‐5
4 KT003‐012_mut A A C + ‐ + ‐31.83 1.47*10‐6
16 SI P. Gawroński et al.
Table S5 Selected results from the delayed fluorescence (DF) measurements performed on T. monococcum wild type KT3‐1 and mutant KT3‐5
DF experiment 17° 22°
Genotype KT3‐1, n=69 KT3‐5, n=69 KT3‐1, n=69 KT3‐5, n=69
% of rhythmic regions 71 58 67 49
Period a 25.85+0.36 27.03+0.68 23.44+0.51 28.19+1
RAE a 0.32+0.04 0.42+0.02 0.38+0.02 0.45+0.03
a Values represent partially weighted means + partially weighted SEM
P. Gawroński et al. 17 SI
Table S6 Phenotypic data of transgenic HvLUX knock‐down (RNAi) plants as compared to cv. ‘Golden Promise’
Conditions Genotype Days to heading SEM n
LD, BG284E10 71.32 0.82 31
17°/15°, BG284E10 azygous 69.27 1.02 11
16h/8h, BG284E11 65.10 0.82 31
day/night BG284E11 azygous 67.20 1.33 10
Golden Promise 62.63 0.96 8
LD, DH_BG284E11‐PP1 b 121.94 a 1.02 16
14°/12°, DH_BG284E11‐PP7 120.93 a 1.29 14 12h/12h, BG353/1E15 b 109.7 2.17 9 day/night BG353/1E15 azygous 106 0 1
BG353/1E01 120.54 1.82 16
BG353/1E01 azygous 118.33 5.24 3
BG353/2E06 119.71 1.34 14
BG353/2E06 azygous 132.5 3.5 4
Golden Promise 112.4 2.54 5
a Only DH_BG284E11‐PP1 and DH_BG284E11‐PP7 headed significantly different to the wild type ‘Golden Promise’ at
P=0.01 as analyzed by paired Student’s t‐test. However, they were about one week later which was against the expectations.
b These plants were also analyzed in the time‐course RT‐qPCR experiment
18 SI P. Gawroński et al.
Table S7 Results from re‐sequencing of the wheat LUX in the collection of 96 accessions
Wheat genome Number of haplotypes found Number of accessions analysed
A 21 69
B 8 40
D 5 8
S 5 6
P. Gawroński et al. 19 SI
Table S8 Selected wheat accessions grouped according to the cultivation status (wild or domesticated) to assess variation in the LUX sequence, expressed as a number of haplotypes
Number of accessions Genome Status Number of haplotypes
47 A wild a 18
34 A domesticated b 8
10 B wild 5
30 B domesticated 4
a Wild = belonging to Triticum boeoticum, T. dicoccoides or T. araraticum species
b Domesticated = belonging to Triticum monococcum, T. dicoccon, T. durum,T. aethiopicum, T. carthlicum, T.