With 12 text-figures Printed in Great Britain
LOCUST WIND RECEPTORS
II. INTERNEURONES IN THE CERVICAL CONNECTIVE*!
BY JEFFREY M. CAMHIJ Biological Laboratories, Harvard University
{Received 17 June 1968)
INTRODUCTION
The first paper in this series (Camhi, 1969 a) showed that a locust's cephalic wind receptors respond to airflow by producing a train of impulses whose frequency depends upon flow rate and direction. Locusts probably use this sensory information be-haviourally in maintaining flapping flight, in stabilizing yaw (Weis-Fogh, 1949, 1950) and, as the succeeding paper will show (Camhi, 19696), in regulating lift. However, the sensory axons, terminating in the suboesophageal ganglion (Camhi, 1969a), are at least one interneurone removed from the thoracic flight motorneurones, whose activity directly determines wing movements.
This paper describes responses obtained from single interneurones in the cervical connective, just postganglionic to the sensory axon terminations. Mechanisms of integrating sensory information and probable roles of certain interneurones in specific flight reflexes are discussed.
MATERIAL AND METHODS
As in the previous paper, all experiments were performed on Schistocerca gregaria Forsk., phasis gregaria, between 2 and 6 weeks after final moult. The culture methods and wind-tunnel construction were as described earlier. Drawing fine teased out bundles of the connective into a pipette electrode allowed recording of single units, as verified by the impulses' regular amplitude and interval. Nerve responses were amplified by a differential input, capacitance-coupled amplifier (Grass P4), displayed on a dual beam oscilloscope (Tektronix 502A) and recorded on moving film with a Grass C4 kymograph camera.
In order to assure exposure of the sensory setae to all possibly meaningful wind parameters, I performed, for each interneurone recorded, a standardized set of airflow manipulations. First, to determine the effect of wind angle on an interneurone response, I increased the wind from rest to some final maintained speed at successively different angles. Next, to study the effect of wind speed, I twice repeated the previous series of experiments using two different final speeds. In these manoeuvres the acceleration from rest to final speed was fixed. Therefore, to determine any effect of varying acceleration, I increased flow rate over increasing time intervals, from rest to some final speed. Finally, for observing any dynamic angular effects, I pivoted the wind jet
• A preliminary report of this work has already appeared (Camhi, 1967).
t This work was supported in part by a predoctoral fellowship from the National Institutes of Health.
% Present address: Section of Neurobiology and Behavior, Division of Biological Sciences, Cornell
35° JEFFREY M. CAMHI
about the head in 2° to 200 steps in both directions at different angular velocities. The ranges of all parameters used included any values which might be meaningful to a locust in the field.
After establishing the preferred wind conditions for any interneurone, tiny jets of air from a fine pipette were useful in determining which setae provided input to that interneurone. Such puffs usually deflected at least three setae, and so the technique did not allow precise localization of sensory inputs. As the results will show, however, stimulation of individual setae was insufficient to evoke an interneurone response. The puff technique was adequate to construct a fairly detailed map of the sensory inputs.
RESULTS
Each of 28 intemeurones from seventeen different animals gave fairly complete sets of data. Deflecting an individual seta shaft was always an insufficient stimulus for evoking an interneurone response. Simultaneous bending of three or four shafts contributing input to an interneurone generally produced at most a few interneurone spikes. All sensory inputs to all intemeurones studied appeared to be excitatory, since simultaneous puffing with two tiny, non-interacting wind jets on separate groups of setae always evoked the same or greater response than one jet alone. Interneurone spike frequencies were always lower than those of the sensory cells under the same stimulus conditions (Camhi, 1969 a).
Four distinct categories of interneurone response appeared. Two of these cell types were slowly adapting and two very rapidly adapting. In four chance simultaneous recordings of a slow-adapting and a fast-adapting interneurone, the latter always dis-played a greater amplitude, suggesting that the fast-adapting cells may have axons of greater diameter, and presumably higher conduction rates. Each of the four types differs in responding maximally to different parameters of wind stimulation and in having dissimilar spatial distributions of input sensory setae.
(1) Wind-indicator cells
Among the slow-adapting intemeurones recorded, 10 fall into a category I shall call 'wind-indicator cells'. Figure 1 shows several responses typical of such a cell. The response gives a slight burst, which quickly levels off to a plateau frequency. The plateau level decreases further by only 20% over a 10 min. interval. The threshold wind speed was less than o-1 m./sec. For 34-5 m./sec. wind, the initial plateau frequency at optimal direction was 85 spikes/sec., or about 65% of the sensory frequency under the same stimulus conditions (Camhi, 1969a).
Minute air jets showed that the wind-indicator cells receive sensory inputs from at least 20 setae, located bilaterally and showing an apparently random spatial distribu-tion. Their angles of curvature ( and therefore their sensory cells' optimal directions— Camhi, 1969 a) range from about 300 left to 300 right of centre. Since the sensory axons do not decussate in the brain (Camhi, 1969 a), this finding indicates that some sensory cells or interneurones must decussate within the suboesophageal ganglion, as Guthrie (1964) has suggested on the basis of histological evidence.
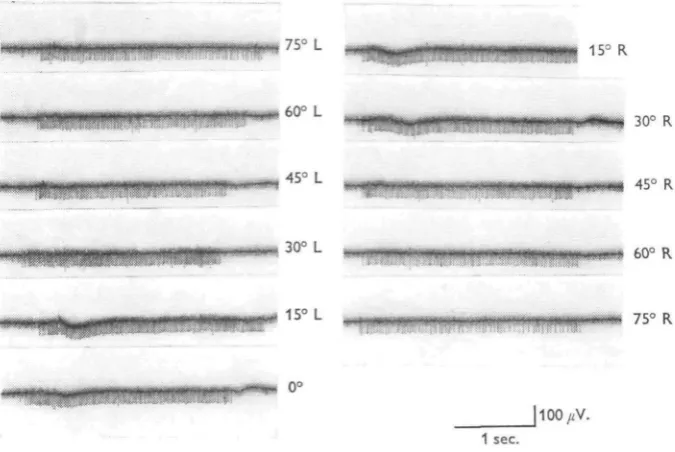
,75° L
160° L
30° R
, 45° L 45° R
30° L 60° R
75° R
[image:3.451.54.392.163.389.2]- " • - - • •' 1 s e c .
Fig. 1. Wind-indicator interneurone. Response to 4-5 m./sec. wind from different directions. Recorded in anterior prothoracic connective. Records read left to right. Note that the response frequencies differ only slightly for wind from widely different angles.
(2) Wind-direction cells
This second group of slow-adapting cells is represented by recordings from six interneurones. Wind flow of 4-5 m./sec. in a cell's optimal direction produced an initial impulse frequency of about 60/sec., adapting within 3 min. to only 1-5/sec. As Fig. 3 indicates, the response is not linear with wind speed, lower speeds evoking unexpectedly few impulses.
Figure 3 also reveals the response's most notable feature—its highly directional nature. The half-peak angles, 270 ± io° for the six cells were consistently more acute than those of any sensory cells recorded (530 + n°). Two of the wind-direction cells observed displayed optimal responses for wind angles as much as 150 from the head-on direction.
By the use of fine air jets it was shown that all setae connecting with any wind-direction intemeurone are clustered on some small region of the cuticle. The shafts of all such setae exhibit very similar angles of curvature (+ 10°), and therefore the sensory cells would show very similar optimal directions (Camhi, 1969 a). Both the side of the head bearing the input setae and the side on which the optimal wind direction appeared were independent of the side of the connective from which the recording was made. This again suggests decussation of some sensory or intemeurone processes in the suboesophageal ganglion.
—' 100
^ ^
'-^ 75
/ / / '
/ A is
\r
/
90° left
V
X.
^ - 50
/ x
1 °
pu
l
V
~^\
v4
\ \
r\F A \
\ \ 190° rightJ
— •
J
/
J
180°
Fig. 2. Wind-indicator intemeurone. Wind direction against spike frequency. Wind speed: O, 4'5 m./sec.; • , 3-0 m./sec.; x , 15 m./sec. The response is not highly directional. Fre-quency is approximately linearly related to wind speed.
(3) Recentre cells
Of the fast-adapting interneurones recorded, seven were of a type which I shall call 'recentre cells'. Figure 5 shows the polar curve of a typical recentre cell. The half-peak angles of the seven cells were fairly broad, 85° + 120. The cells responded almost identically to vastly dissimilar accelerations from rest to a given terminal velocity (Fig. 10, dashed curves).
353
[image:5.451.72.358.54.390.2]180°
Fig. 3. Wind-direction interneurone. Wind direction against spike frequency. O, 4'5 m./sec.; • , 3-0 m./sec.; x , 1-5 m./sec. The response is very highly directional. Frequency is a non-linear function of wind speed, lower speeds evoking unexpectedly few impulses.
15'
100/iV.
-» 1 in I T 1111 inni if 200 msec.
[image:5.451.50.378.451.576.2]354
180°
Fig. 5. Recentre intemeurone. Wind direction against total number of spikes. 3 0 m./sec. The response is not highly directional.
I c r c / c / c
I c I c I c
a vigorous burst of spikes. Recentring from the opposite side evoked no response. The preferred side was not necessarily the same as the side of the connective from which the recording was made.
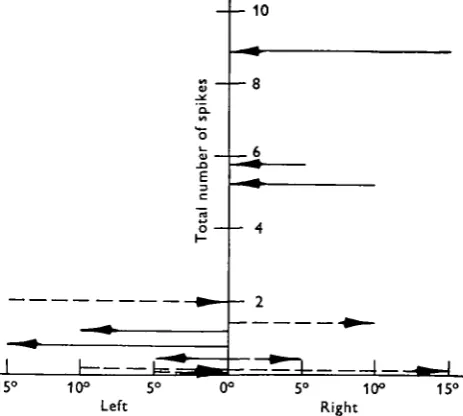
Figure 6 demonstrates the pivoting responses. Figure 7 summarizes all the turning responses of a typical recentre cell. It is of particular interest that all pivots of the wind jet began and ended at angles from which a near maximal response would attend the onset of wind.
10
15° 10° 5
Left
[image:7.451.110.342.160.368.2]15°
Fig. 7. Recentre interneurone. Direction and extent of wind pivot against total number of spikes evoked. The length and direction of each arrow indicates the extent and direction of the turn. Dashed arrows represent turns to the right, solid arrows turns to the left. An arrow's level on the vertical axis indicates the number of spikes evoked by that turn, averaged over five re-sponses. This cell's preference was for wind recentring from the right.
Rapid wind recentring gives a greater response (shorter interspike intervals) than slow recentring. As plotted in Fig. 8, the response of a typical recentre cell was greatest for pivots lasting less than 100-150 msec. Thus recentre interneurones appear to be primarily sensitive to rapid changes to wind angle from a specific side to the head-on direction.
No clearly denned spatial distribution of sensory seta inputs has appeared for this group of cells. Usually inputs are bilateral, but may occur either in clusters or in widely separated positions.
(4) Acceleration cells
This final group, represented by five recorded interneurones, also displayed fast-adapting responses. The onset of wind from optimal direction evoked a burst of im-pulses, provided the wind reached a speed of at least 1 -0-1-5 m./sec. For velocities below about i-o m./sec. these cells remained silent. Wind pivots as great as 200 evoked no response. As the graph of Fig. 9 shows, half-peak angles for acceleration cells were very broad, 1050 ± 300.
Rapid wind accelerations from rest produced a more vigorous burst (shorter inter-spike intervals) than did slower acceleration to the same final velocity. Figure 10 shows for a typical acceleration cell, the inter-spike interval as a function of acceleration rate for increases from rest to 4-5 m./sec. Included for comparison is a plot of the same data for a typical recentre interneurone. The graph suggests that the sensitivity to very rapid wind onset is a unique property of the acceleration interneurones.
300
2S0
200
.9 150
.•a
« 100
50
50 100 150 200 250 300
Duration of wind pivot (msec.)
350 -W0 450
Fig. 8. Recentre interneurone. Duration of 10° wind recentring against inter-spike interval for the first three intervals. O, i st interval; • , second interval; x , third interval. TTie cell responds maximally to turns lasting less than about 150 m./sec.
All sensory setae contributing input to acceleration interneurones are situated bilaterally along the head's anterior and lateral margins. Thus some sensory axons or interneurones appear again to decussate in the suboesophageal ganglion. The shafts of input setae to any one interneurone curve in widely dissimilar directions, ranging from about 200 left to 200 right of centre.
DISCUSSION
Table 1 summarizes the salient characteristics of the four types of interneurone response recorded. Every interneurone recorded thus far has conformed to the de-scriptive grouping presented.
(1) Integration of sensory inputs
357
[image:9.451.74.366.71.381.2]180°
Fig. 9. Acceleration intemeurone. Wind direction against total number of spikes. 3-0 m./sec. The response is not highly directional.
200
-I
1S0s 100
50
100 200 300 -400 500 600 Time interval of wind acceleration (msec.)
700 800
Fig. 10. Interval of wind acceleration against inspike interval. Acceleration from rest to ter-minal velocity of 4-5 m./sec. Acceleration intemeurone: O , first spike interval; • , second spike interval. Recentre intemeurone: x , first spike interval; A , second spike interval. T h e accelera-tion cell* are much more discriminating of acceleraaccelera-tion rate.
[image:9.451.47.392.421.601.2]both located in the suboesophageal ganglion (Camhi, 1969a), these are not known to be first-order interneurones. Latency determinations of wind-stimulated interneurones do not adequately resolve the question of whether other neurones are connected in series within the ganglion, because conduction rates of different sensory axons differ by a factor of two (Camhi, 1969a). Moreover, the sensory axons change diameter as they enter the suboesophageal ganglion (Guthrie, 1964), presumably affecting con-duction rates. Nevertheless, by treating the ganglion as a ' black box' having accessible inputs and outputs, several deductions and suggestions can be offered regarding the integrative physiology of the ganglion.
Table 1 Interneurone type Wind-indicator cell Wind-direction cell Recent recell Acceleration cell
N o .
recorded
I O
6
7
5
Fast or slow adapting Slow Fairly slow Fast Fast Half-peak angle Broad (I40°±20°) Narrow (27°±IO°) Fairly broad (85° ±12°) Broad (iO5°±3°°) Wind Pronounced pivot acceleration response response None None
+ for None pivots into
optimal direction from either side
+ for None pivots from only one side onto centre None + Input setae distribution Bilateral; all areas;diverse curvature angles Unilateral; restricted area; similar curvature angles ? Bilateral; anterior and lateral margins; diverse curvature angles
First, since all sensory axons recorded give slow-adapting responses (Camhi, 1969 a) the suboesophageal ganglion must be the site of modification into the more rapidly adapting responses of wind-direction, recentre and acceleration cells.
Secondly, as stated earlier, each type of interneurone responds only to simultaneous excitation of several sensory cells. Since all sensory inputs appear excitatory, an intemeurone's activity presumably represents the summation of all impinging sensory impulses. The following several paragraphs suggest how the different interneurones' observed direction responses might result from summation of input excitatory impulses.
359
50060°
Left Right
Fig. 11 a, 6. Suggested scheme for integration of sensory direction information. Graphs plot wind direction against spike frequency. The many identical low curves in both diagrams repre-sent the direction/responses of several sensory cells. The highest curve in each diagram (dashed line) is the direction/response curve of the total number of impulses in all sensory cells shown. The lowest curve of each diagram (thick, solid line) represents the direction/response curve of an intemeurone having input from the sensory cells shown. See text for further explanation.
Impulses
60° 40°
Left 20° 20° Right 40° 60°
40°
Left 20° 0° 20° 40°Right
360 JEFFREY M. CAMHI
indicated by the dashed curves, which represent simply the sums of all underlying sensory curves. If, as in Fig. 11 a, the sensory inputs display widely divergent optimal directions, the summed curve will show far less acute half-peak angle (be much broader) than any one sensory curve. If, however, the input-direction optima cluster about a particular direction, as in Fig. \ib, the half-peak angle of the summed curve will be about the same as that of any one sensory curve.
Since all interneurones recorded responded with impulse frequencies lower than those of the sensory cells, the summed (dashed) curves of Fig. 11 a and b must be greatly reduced in height, if they are to describe actual intemeurone responses. Wind-indicator interneurones have inputs resembling those illustrated in Fig. 11 a. These cells were found to exhibit impulse frequencies approximately linear with wind speed. Since sensory responses are also linearly related to wind speed (Camhi, 1969a), this observation implies that the response of the wind-indicator cell is a linear function of the total number of impinging sensory impulses. Accordingly, reducing by a constant factor the height of each point along the dashed curve of Fig. ixa results in a new plot, repre-sented by the heavy solid line. Figure 12a compares this hypothetical curve with several experimentally determined curves for wind-indicator cells. The hypothetical and experimental curves are closely similar.
Wind-direction cells have sensory inputs resembling those plotted in Fig. 116. These cells were shown to exhibit impulse frequencies which were non-linearly related to air speed, lower speeds evoking unexpectedly few spikes. However, as mentioned pre-viously, the input sensory frequency appears linear with air speed. Re-shaping the dashed curve of Fig. 11 b in accordance with this non-linearity requires that one reduce
non-uniformly the heights of various points along the dashed curve, such that each
impulse at higher frequencies is reduced less than those at lower frequencies. The heavy solid curve of Fig. 116 shows the result of such a non-linear reduction. This curve was constructed by multiplying each point of the dashed curve by a constant fraction times the ratio
impulse frequency of the point maximum impulse frequency
Experimentally observed values are plotted together with the hypothetical curve in Fig. 12b. The two are closely similar in all important respects.
(2) Behavioural correlations
Weis-Fogh (1949) showed that in tethered locusts wind stimulation of the facial sensory setae initiates flight, the animal continuing to fly as long as the wind persists. Locusts may also use the setae in yaw-stabilization manoeuvres (Weis-Fogh, 1949, 1950). The next paper in this series (Camhi, 19696) will describe a further function which they mediate, bilaterally symmetrical control of forewing twist, resulting in lift regulation. All these manoeuvres involve flight muscles whose motorneurone cell bodies he in the thoracic ganglia. Therefore, at least some of the cervical connective interneurones reported here must represent a part of the controlling circuitry for these flight reflexes. It is possible at this point to suggest specific roles for certain of the interneurone categories in some of these reflexes.
Weis-Fogh's observation that tethered flight continues as long as wind flows, usually ceasing upon wind interruption, implies either a maintained (tonic) or an on-off neural correlate. For the latter type of signal, the 'on' response would close a switch, setting the thoracic flight motor in operation (Wilson, 1961), an 'off' response later re-opening the switch to arrest the motor. One finding which argues against 'on-off' coding is that repetitive electrical stimulation of the ventral cord, which presumably would stimulate any interneurones normally responding to wind cessation (Wilson & Wyman, 1965), maintains normal flight activity. Moreover, no interneurones recorded in the present study displayed 'off' responses. Flight maintenance probably results then from the activity of a slow-adapting neural circuit.
The only interneurone responses I have recorded which adapt sufficiently slowly to maintain flight over an interval of several hours are those of the wind-indicator cells. This suggests that wind-indicator interneurones, or neurones with which these make synaptic contact, converge upon and excite neurones of the pterothoracic flight motor (Wilson, 1961). The next paper in this series (Camhi, 19696) will present evidence suggesting that wind-indicator cells also connect with motorneurones controlling forewing twist.
The broad direction response of the wind-indicator interneurones is of particular interest in relation to this function. The broad response would enable the locust to maintain flight even while experiencing a severe yaw.
The wind-direction cells are the only highly directional interneurones recorded in these experiments. Since detecting the direction of the relative wind is a prerequisite for an aerodynamic yaw-stabilizing reflex, the responses of these cells suggest a func-tion of signalling the direcfunc-tion of yaw. This suggesfunc-tion implies that wind-direcfunc-tion interneurones influence, perhaps through other interneurones, the activity of the motorneurones of forewing twist muscles (Gettrup & Wilson, 1964; Dugard, 1967; Waldron, 1968).
SUMMARY
1. The cervical connective of the desert locust contains interneurones giving responses of four different types to wind stimulation of the facial wind receptor setae. The four categories are:
(a) 'Wind-indicator cells', giving a slowly adapting response to wind from any of a wide range of directions.
(£) 'Wind-direction cells', displaying fairly slowly adapting responses, and sen-sitive to wind only within a very narrow range of directions. Pivoting wind into a cell's optimal direction evokes a full response.
(c) 'Recentre cells', showing rapidly adapting responses to wind onset from a fairly wide directional range. Rapidly shifting the wind direction from only one pre-ferred side into the centre evokes a vigorous burst.
(d) 'Acceleration cells', giving a rapidly adapting response which is especially
pronounced for rapid wind accelerations.
2. Interneurones connect with several sensory setae, whose spatial distribution and angles of curvature are unique to each interneurone type.
3. A suggested integrative mechanism relates the angles of curvature of the input setae to the direction response of each interneurone type.
4. The responses suggest roles for certain interneurone categories in flight maintenance and yaw-correcting reflexes.
I wish to thank Professors Ian Cooke and Kenneth Roeder whose critical reading of this manuscript was most helpful.
REFERENCES
CAMHI, J. (1967). Locust aerodynamic setae: sensory and interneuron responses. Am. Zoologist. 7 (4)
127.
CAMHI, J. (1969a). Locust wind receptors. I. Transducer mechanics and sensory response. J. exp. Biol. 5°> 33S-348.
CAMHI, J. (19696). Locust wind receptors. III. Contribution to flight initiation and lift control. J. exp.
Biol. 50, 363-373.
DUGARD, J. J. (1967). Directional changes in flying locusts. J. Insect. Phytiol. 13, 1055-63.
GUTHRIE, D. M. (1964). Observations on the nervous system of the flight apparatus in the locust
Schistocerca gregaria. Q. Jl microic. Set. 105, (2), 183-201.
GETTRUP, E. & WILSON, D. M. (1964). The lift-control reaction of flying locusts. J. exp. Biol. 41,183-90. WALDRON, I. (1968). Neural mechanisms by which controlling inputs influence motor output in the
flying locust. J. exp. Biol. 47, 213-28.
WEIS-FOGH, T. (1949). An aerodynamic sense organ stimulating and regulating flight in locusts. Nature,
Lond. 163, 873-4.
WEIS-FOGH, T. (1950). An aerodynamic sense organ in locusts. 8th Int. Congress Entomol. (Stockholm, 1948), pp. 584-8.
WILSON, D. M. (1961). The central nervous control of flight in a locust. J. exp. Biol. 38, 471-90. WILSON, D. M. & WEIS-FOGH, T. (1962). Patterned activity of co-ordinated motor units, studied in
flying locusts. J. exp. Biol. 39, 643-67.