CRAYFISH SKELETAL MUSCLE REQUIRES BOTH INFLUX OF
EXTERNAL Ca
2+AND Ca
2+RELEASE FROM INTERNAL
STORES FOR CONTRACTION
HIDEKI USHIO, SHUGO WATABE*
Laboratory of Marine Biochemistry, Faculty of Agriculture, University of Tokyo, Tokyo 113, Japan
and MASAMITSU IINO
Department of Pharmacology, Faculty of Medicine, University of Tokyo, Tokyo 113, Japan
Accepted 8 April 1993
Summary
The isometric tension and membrane potential of single skeletal muscle fibres from the flexor muscle of the carpopodite in the meropodite of crayfish Procambarus clarkii (Girard) were studied to determine whether crayfish muscle contraction requires Ca2+
release from the sarcoplasmic reticulum. Contraction elicited by brief extracellular electrical stimulation was reduced by the removal of Ca2+ or by the addition of
25mmol l21nicardipine in crayfish Ringer’s solution. Addition of 30mmol l21ryanodine
with 1mmol l21caffeine induced a transient contracture, the peak tension of which was
10–30% of that of the high-K+ induced contracture and which declined to the
pretreatment level in 20–60min. After ryanodine–caffeine treatment, 30mmol l21
caffeine failed to induce contraction, suggesting that intracellular Ca2+stores had been
exhausted by the treatment. Extracellular electrical stimulation also failed to induce contraction after ryanodine–caffeine treatment, although the resting potential was not changed. These results suggest that Ca2+ release from the sarcoplasmic reticulum,
together with Ca2+ influx via nicardipine-sensitive Ca2+ channels, is essential to the
contraction of crayfish leg muscle fibres after a brief membrane depolarization.
Introduction
Ca2+release from the sarcoplasmic reticulum (SR) is important in the regulation of
vertebrate skeletal and cardiac muscle contraction (Endo, 1977). There is a regular array of structures known as feet at the junction between the terminal cisternae of the SR and the transverse tubules (Block et al. 1988). These correspond to the Ca2+release channel
of the SR (Inui et al. 1987a,b; Lai et al. 1988). Ryanodine, a plant alkaloid, binds to this channel and at low concentrations fixes it in an open state (Fleischer et al. 1985; Rousseau
*To whom correspondence should be addressed.
Key words: crayfish, Procambarus clarkii , skeletal muscle, sarcoplasmic reticulum, ryanodine, Ca2+
et al. 1987; Nagasaki and Fleischer, 1988). Using ryanodine as a marker, the Ca2+release
channel has been purified (Hymel et al. 1988; Imagawa et al. 1987; Inui et al. 1987a,b) and sequenced (Takeshima et al. 1989). The Ca2+release channel is believed to face onto
dihydropyridine-binding proteins in the transverse tubule membranes (Block et al. 1988). The two important proteins associated with the foot are also present in crayfish skeletal muscle. Formelova et al. (1990) isolated ryanodine-binding proteins from crayfish abdominal muscle and showed that they exhibited a Ca2+ conductance in planar lipid
bilayers. Recently, Krizanova et al. (1990) have isolated dihydropyridine-binding proteins from crayfish abdominal muscle and found that the properties of these proteins were similar to those of their mammalian counterparts. Furthermore, feet structures are also observed in crayfish muscle (Loesser et al. 1992).
In crustacean skeletal muscle fibres, electrical stimulation triggers local, non-propagatable depolarization (Fatt and Katz, 1953a,b), which subsequently induces an inward Ca2+current (Hagiwara and Naka, 1964; Hagiwara et al. 1964). The Ca2+influx
has been reported to be essential (Gainer, 1968; Matsumura, 1978) or sufficient to (Atwater et al. 1974) give rise to contraction of crustacean muscles. It has been shown that Ca2+ release from the SR is necessary for contraction of barnacle muscle fibres
(Ashley and Lea, 1978). Mounier and Goblet (1987) investigated the effects of caffeine and procaine on the contraction of crab leg muscle fibres and proposed that Ca2+release
from internal stores is required for contraction of these muscle fibres. However, procaine affects both K+ and Ca2+ currents in crustacean muscle membranes (Hagiwara et al.
1969), and further evidence is clearly necessary to confirm the proposal of Mounier and Goblet (1987).
The purpose of this study was to examine the effects of pharmacological agents known to affect excitation–contraction coupling in vertebrate skeletal muscle, such as ryanodine, caffeine, nicardipine and diltiazem, on the contraction of single crayfish muscle fibres in search of the source of Ca2+for their contraction.
Materials and methods
Single skeletal muscle fibres were prepared from the proximal flexor muscle of the carpopodite in the meropodite segment of the crayfish Procambarus clarkii (Girard) leg. These fibres were 5–8mm long and were shaped like elliptical cylinders, 300–500mm along the major axis and 50–150mm along the minor axis. Excised muscle fibres were put in a chamber at 10˚C filled with a crayfish Ringer’s solution (see below). The fibres were also observed in a transmission electron microscope by conventional methods as described in Brandt et al. (1965).
Solutions
The crayfish Ringer’s solution contained 205mmol l21 NaCl, 5.4mmol l21 KCl,
13.5mmol l21CaCl2, 5.6mmol l21MgCl2and 2.4mmol l21 Hepes (pH7.2). The
high-K+solution contained 132.4mmol l21sodium propionate, 8mmol l21NaCl, 70mmol l21
potassium propionate, 13.5mmol l21 calcium propionate and 5.6mmol l21 MgCl2 to
containing tetraethylammonium (TEA+), i.e. 107mmol l21TEACl, 105mmol l21NaCl,
5.4mmol l21 KCl, 13.5mmol l21 CaCl
2, 5.6mmol l21 MgCl2and 2.4mmol l21Hepes
(pH7.2), was used. In the Ca2+-free solution, CaCl2in the crayfish Ringer’s solution was
replaced by the same concentration of MgCl2.
All chemicals were of reagent grade from Sigma (St Louis), except that ryanodine and nicardipine were obtained from Wako Chemicals (Tokyo) and Yamanouchi Pharmaceutical (Tokyo), respectively.
Measurement of tension and membrane potential
Single fibres were stretched to 1.25 times their resting length. For extracellular stimulation, rectangular current pulses of 20ms duration were passed through platinum electrodes placed parallel to the fibre. Isometric tension was measured with a strain gauge transducer and displayed using a pen recorder. To measure membrane potential, a microelectrode filled with 3mol l–1KCl (resistance approximately 10 MV) was inserted
near one end of the fibre. In the experiments using intracellular stimulation, the second microelectrode was inserted about 30mm from the recording electrode and currents of 1.2mA were passed through an electric isolator. Changes in the membrane potential were monitored on an oscilloscope. Both tension and membrane potential signals were digitized using an analog-to-digital converter in a personal computer (PC-9801 VM, NEC, Tokyo) and stored on floppy disks for later analyses.
Results
Effect of Ca2+removal
Single fibres excised from the proximal area of the carpopodite flexor muscle had a sarcomere length of about 8mm (range 7–10mm) under the electron microscope.
The peak tension elicited by extracellular electrical stimulation of the single fibres was about 4–9mNmm22 with a mean ± S.E.M. of 6.2±0.6mNmm22 (N=9). The high-K+
solution (70mmol l21K+) induced a transient contracture, which attained a peak tension
of about 135mNmm22in about 10s ( N=4).
When a fibre was immersed in Ca2+-free solution, the electrically evoked contraction
faded rapidly (Fig. 1A). The contraction was promptly restored by washing with normal crayfish Ringer’s solution. The membrane potential of the fibres in the Ca2+-free solution
was the same as that before such treatment, i.e. it lay within the range 275 to 280mV (N=24).
Effects of Ca2+antagonists
Calcium antagonists such as nicardipine and diltiazem bind to L-type Ca2+channels
and block the Ca2+current (Bean, 1984; Kanaya et al. 1983; Sanguinetti and Kass, 1984).
Since dihydropyridine-binding proteins or L-type Ca2+channels have been isolated from
crayfish skeletal muscle, we examined the effects of these Ca2+ antagonists on the
contraction of crayfish leg muscle fibres. Following the application of 25mmol l21
contraction was abolished after 15–20min (Fig. 1B). The resting membrane potential after this treatment was in the range 277 to 281mV (N=4), which was not significantly different from the level in normal crayfish Ringer’s solution. Diltiazem had no effect on the contraction elicited by extracellular electrical stimulation. It has been reported that diltiazem binds to Ca2+ channels when they are in an inactivated state (Kanaya et al.
1983). In high-K+solution, a contracture was induced in crayfish fibres in the presence of
diltiazem to see whether diltiazem is effective when Ca2+ channels are inactivated.
However, even at concentrations up to 100mmol l21, diltiazem had no effect on the
subsequent contraction (data not shown).
Effects of caffeine and ryanodine
Contractions elicited by extracellular electrical stimulation were enhanced by the addition of 1mmol l21caffeine; the peak tension was about 10 times greater than that in
normal crayfish Ringer’s solution (Fig. 2A). Irregular contractures were sometimes observed in the presence of caffeine (Fig. 2A). They were due to local contractures, visible under a binocular microscope, propagating from one end of the fibre to the other with gradual attenuation. Caffeine at a concentration of 30mmol l21 induced a strong
contracture so that the connection between the fibre and the tendon broke (data not shown). Removal of 1mmol l21caffeine restored contractions induced by extracellular
electrical stimulation to pretreatment levels (Fig. 2A).
Addition of 30mmol l21 ryanodine with 1mmol l21 caffeine initially enhanced the
contraction induced by extracellular stimulation and then induced a contracture, the tension of which was 20–30% of that of the high-K+-induced contracture (Fig. 2B). This
contracture was transient and subsided to the pretreatment level in about 20–30min. Following the ryanodine–caffeine treatment, no contraction was observed in response to extracellular stimulation (Fig. 2A). The application of 30mmol l21caffeine also failed to
A
B
25mmoll−1nicardipine 2min 0mmoll−1 Ca2+
[image:4.612.118.403.67.254.2]0.1mN
Fig. 1. Effects of the removal of Ca2+(A) and the addition of 25mmol l21nicardipine (B) on
induce a contracture after ryanodine–caffeine treatment (Fig. 2A). However, a high-K+
-induced contracture was observed even after the treatment (Fig. 2C), although the peak tension was about half and the decay of tension was slower than those of the control high-K+-induced contracture (Fig. 2B). The high-K+-induced contracture rose to 50% of the
peak tension in 2.50±0.41s (N=4) in the control condition. The corresponding duration after ryanodine–caffeine treatment was 13.53±1.89s (N=4), and the difference between these two values was significant (P<0.01, t-test). The resting membrane potential after the ryanodine–caffeine treatment was in the range 274 to 279mV (N=9), which is not significantly different from that in normal Ringer’s solution.
Application of 30mmol l21 ryanodine without caffeine also induced a transient
contracture, the tension of which was about half of that during ryanodine–caffeine treatment and which decayed more slowly. After ryanodine treatment, extracellular electrical stimulation and 30mmol l21caffeine also failed to induce contraction (data not
shown).
Action-potential-induced contractions in the presence of TEA+
Neither extracellular nor intracellular stimulation induced action potentials in the crayfish leg muscle fibres. In the presence of TEA+, however, the fibres produced action
potentials accompanied by strong contractions. The action potential had a plateau phase lasting 300–600ms and a peak tension of about 100mNmm22, which was attained about
300ms after the stimulus (Fig. 3A,B, top traces).
1mmoll−1 caffeine 30mmoll−1 caffeine
70 mmoll−1K+ 70 mmoll−1K+
2min
2.5 mN
0.5 mN A
B C
1mmoll−1 caffeine + 30mmoll−1ryanodine
Fig. 2. (A) The effects of successive treatment with 1mmol l21caffeine and 30mmol l21
ryanodine plus 1mmol l21 caffeine on the contraction induced by extracellular electrical
stimulation and the aftereffect of the ryanodine treatment on the contraction induced by 30mmol l21caffeine. Contractures induced in 70mmol l21K+before (B) and after (C) the
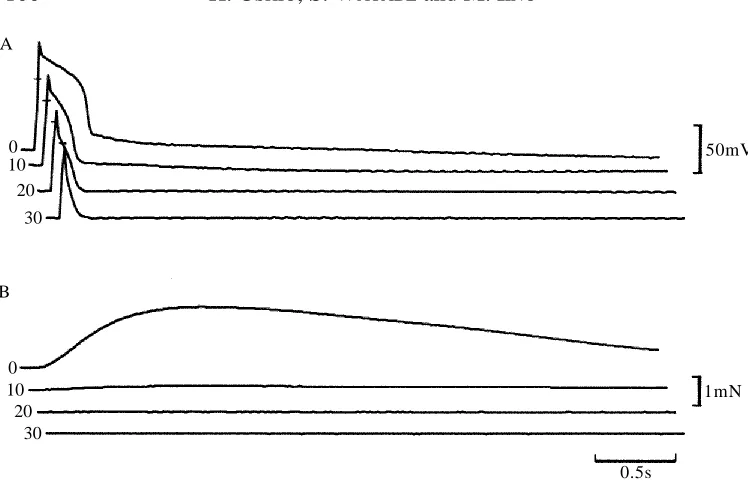
[image:5.612.68.425.73.274.2]When the fibres were immersed in Ca2+-free TEA+-containing solution, both action
potential and contraction were reduced gradually and disappeared in 10–30min (Fig. 3). The depolarization remaining after 30min in Ca2+-free condition was mostly a passive
response to intracellular stimulation. The slow time course of the effect of Ca2+
withdrawal on the action potential duration could be explained by the difficulty of removing Ca2+from the multibranched cleft and transverse tubule systems (Brandt et al.
1965; Hoyle et al. 1973). Addition of 25mmol l21 nicardipine also reduced both the
action potential duration and the contraction (Fig. 4).
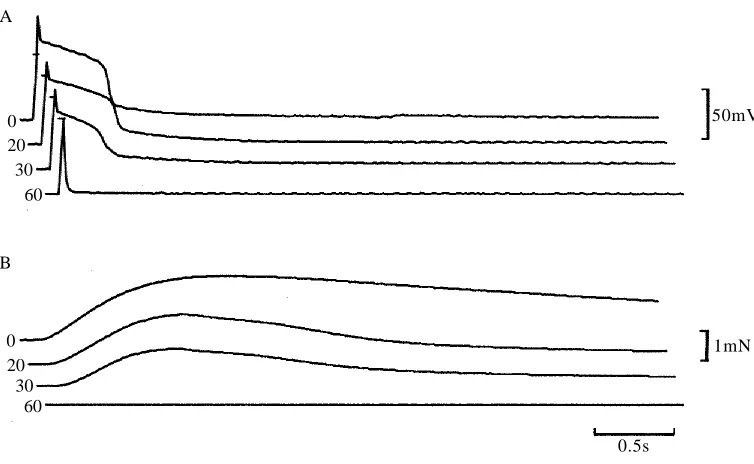
Action potentials in the presence of TEA+had considerably longer time courses after
ryanodine–caffeine treatment (Fig. 5A,B). The duration of this prolonged action potential was not influenced by tetrodotoxin at concentrations up to 500mmol l21(data not shown).
Although contractions elicited by extracellular stimulation were abolished by ryanodine–caffeine treatment in the absence of TEA+ (Fig. 2), a prolonged action
potential during the treatment in the presence of TEA+was followed by a slow rise in
tension (Fig. 5B, lower trace). Peak tension was attained after about 10s. In Ca2+-free
TEA+-containing solution, both the action potential and the rise in tension were almost
abolished (Fig. 5C).
Discussion
Our results clearly show that the electrically evoked contraction of single crayfish muscle fibres is greatly diminished after treatment with ryanodine, with or without
B A
0 10
20
30
0 10
20 30
50mV
0.5s
[image:6.612.60.434.59.300.2]1mN
Fig. 3. Effects of the removal of Ca2+from the crayfish Ringer’s solution on the action
potential induced by intracellular stimulation (using a 1.2mA current) in the presence of TEA+
simultaneous caffeine application. It has been shown in vertebrate skeletal muscle that ryanodine binds to the Ca2+-release channels of the SR when the channels are open and
fixes them in an open state (Fleischer et al. 1985; Rousseau et al. 1987; Nagasaki and Fleischer, 1988), resulting in the inhibition of Ca2+release from the SR. The same effect
was observed in the crayfish muscle fibres: caffeine blocked the release of Ca2+after
ryanodine treatment (Fig. 2). These results indicate that electrically evoked contraction depends critically on Ca2+release from the SR.
To determine whether ryanodine treatment had any effect on the membrane potential and/or on the voltage-dependent Ca2 + channel, we studied the effect of ryanodine on
action-potential-induced contractions. Crayfish muscle fibres do not usually generate action potentials, but these can be induced when K+currents are suppressed by TEA+
(Fatt and Ginsborg, 1958; Hagiwara et al. 1964; Hencek and Zachar, 1977). Such action potentials are resistant to tetrodotoxin but are sensitive to nicardipine (Fig. 4 ) , suggesting that the inward currents are mainly carried by Ca2 +. After ryanodine
treatment, the resting membrane potential was unchanged and the action potential duration was prolonged, causing enhanced Ca2 +i n flux. However, the development of
tension was slower and smaller (Fig. 5). Similarly, high-K+-induced depolarization
produced slower and smaller forces after ryanodine treatment (Fig. 2). These results c o n firm that Ca2 +i n flux is essential (Fatt and Katz, 1953a,b), but is not sufficient, for
contraction following brief depolarizations. Under such circumstances, Ca2 + r e l e a s e
from the SR is important for the initiation of rapid tension development. However, when
B A
50mV
0.5s 0
20 30
60
0 20
30 60
[image:7.612.53.430.72.304.2]1mN
Fig. 4. Effects of addition of 25mmol l21 nicardipine on the action potential induced by
intracellular stimulation in the presence of TEA+(A) and the accompanying development of
there is a prolonged depolarization, Ca2 +i n flux alone may induce contraction, albeit
very slowly (Fig. 5 B ) .
The prolongation of the action potential in the ryanodine-treated fibres could be explained by a reduction in the release of Ca2+. The inactivation of inward Ca2+current is
modulated by cytosolic Ca2+ concentration in vertebrate cardiac (Lee et al. 1985),
vertebrate smooth (Jmari et al. 1986) and insect skeletal (Ashcroft and Stanfield, 1981) muscles and in other excitable cells (Eckert and Chad, 1984). If this is also the case in crayfish muscle fibres, the inhibition of Ca2+release from the SR by ryanodine–caffeine
treatment will delay inactivation of Ca2+ channels, thus prolonging the duration of
membrane depolarization. In addition, part of the Ca2+-activated outward K+current is
resistant to TEA+(Mounier and Vassort, 1975a,b). This K+current must contribute to
repolarization, and the reduced rise in cytoplasmic Ca2+ concentration due to
ryanodine–caffeine treatment is expected to delay repolarization. Confirmation of our hypothesis requires further studies on the kinetics of both Ca2+and K+currents after
ryanodine treatment using a voltage-clamp method.
The ryanodine-binding protein has been isolated from crayfish muscle and shown to have Ca2+ channel activity (Formelova et al. 1990). Our results demonstrate that
0.5s
2mN 50mV B
A
[image:8.612.68.417.57.335.2]C
Fig. 5. Effects of ryanodine–caffeine treatment on action potential and tension development in the presence of TEA+. The action potentials (top traces) and the development of tension
(bottom traces) were recorded before ryanodine–caffeine treatment (A), after ryanodine–caffeine treatment in the presence of Ca2+ (B) and after ryanodine–caffeine
treatment in the absence of Ca2+(C). The bars in the upper traces represent the 0mV potential
ryanodine has a profound effect on the Ca2+ release mechanism. Goblet and Mounier
(1986) showed that a low concentration of Ca2+(about 1027mol l–1) induced Ca2+release
from the SR of crab leg muscles. Mounier and Goblet (1987) demonstrated, using the same type of muscle, that the tension development induced by voltage-clamp was inhibited by procaine, an inhibitor of Ca2+-induced Ca2+release from the SR (Endo,
1985). Similarly, the Ca2+-induced Ca2+ release blocker induced a large decrease in
intracellular Ca2+ transients following step voltage changes in crayfish muscle fibres
(Gyorke and Palade, 1992). These results indicate the presence of a Ca2+-induced Ca2+
release mechanism in the crayfish muscle and its involvement in excitation–contraction coupling. Since Ca2+influx is essential for the initiation of contraction, it is possible that
the inward Ca2+current induces a secondary Ca2+release from the SR through a Ca2+
-induced Ca2+release mechanism.
Crustacean muscle fibres can be classified into two broad groups: the fast phasic muscle fibres with short sarcomeres and the slow tonic ones with long sarcomeres. Both the shape and the disposition of intracellular membrane systems are different in the two groups (Abbott and Parnas, 1965; Fahrenbach, 1967; Jahromi and Atwood, 1967; Eastwood et al. 1982; Franzini-Armstrong et al. 1986). In lobster fast phasic muscle, Crowe and Baskin (1981) showed that the transverse tubules have a surface area about 50% greater than that of the SR and suggested that the large quantities of Ca2+that enter
the sarcoplasm from the extracellular space through the sarcolemma and the tubules of this muscle are sufficient to activate the contractile machinery. In contrast, fibres of the slow tonic type have less well developed tubular systems (Brandt et al. 1965). The crayfish fibres used in the present study belong to the latter type according to Govind
et al. (1981) on the basis of their sarcomere lengths (about 8mm). Therefore, the role of the SR in excitation–contraction coupling can vary according to the type of muscle fibre considered.
The different effects of nicardipine and diltiazem on electrically evoked contraction were unexpected. Both drugs have a profound effect on Ca2+ currents in vertebrate
cardiac cells (Lee and Tsien, 1983; Sanguinetti and Kass, 1984). It will be interesting to determine whether crayfish Ca2+channels lack binding sites for diltiazem.
The expenses of the present study were supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science and Culture of Japan.
References
ABBOTT, B. C. ANDPARNAS, I. (1965). Electrical and mechanical responses in deep abdominal extensor muscles of crayfish and lobster. J. gen. Physiol. 48, 919–931.
ASHCROFT, F. M. AND STANFIELD, P. R. (1981). Calcium dependence of the inactivation of calcium currents in skeletal muscle fibers of an insect. Science 213, 224–226.
ASHLEY, C. C. ANDLEA, T. J. (1978). Calcium fluxes in single muscle fibres measured with a glass scintillator probe. J. Physiol., Lond. 282, 307–331.
ATWATER, I., ROJAS, E. ANDVERGARA, J.(1974). Calcium influxes and tension development in perfused single barnacle muscle fibres under membrane potential control. J. Physiol., Lond. 243, 523–551. BEAN, B. P. (1984). Nitrendipine block of cardiac calcium channels: high-affinity binding to the
BLOCK, B. A., IMAGAWA, T., CAMPBELL, K. P. ANDFRANZINI-ARMSTRONG, C.(1988). Structural evidence
for direct interaction between the molecular components of the transverse tubule/sarcoplasmic reticulum junction in skeletal muscle. J. Cell Biol. 107, 2587–2600.
BRANDT, W. P., REUBEN, J. P., GIRARDIER, L. ANDGRUNDFEST, H.(1965). Correlated morphological and
physiological studies on isolated single muscle fibers I. Fine structure of the crayfish muscle fiber.
J. Cell Biol. 25, 233–260.
CROWE, L. M. ANDBASKIN, R. J. (1981). Activation of the contractile system in crustacean muscle: ultrastructural evidence for the role of the T system. Tissue & Cell 13, 153–164.
EASTWOOD, A. B., FRANZINI-ARMSTRONG, C. ANDPERACCHIA, C. (1982). Structure of membranes in crayfish muscle: comparison of phasic and tonic fibres. J. Muscle Res. Cell Motil. 3, 273–294. ECKERT, R. AND CHAD, J. E. (1984). Inactivation of Ca channels. Prog. Biophys. molec. Biol. 44,
215–267.
ENDO, M.(1977). Calcium release from the sarcoplasmic reticulum. Physiol. Rev. 57, 71–108.
ENDO, M. (1985). Calcium release from sarcoplasmic reticulum. Curr. Topics Membr. Transport 25, 181–230.
FAHRENBACH, W. H. (1967). The fine structure of fast and slow crustacean muscles. J. Cell Biol. 35,
69–79.
FATT, P. ANDGINSBORG, B. L. (1958). The ionic requirements for the production of action potentials in crustacean muscle fibres. J. Physiol., Lond. 142, 516–543.
FATT, P. ANDKATZ, B. (1953a). The electrical properties of crustacean muscle fibres. J. Physiol., Lond. 120, 171–204.
FATT, P. ANDKATZ, B. (1953b). Distributed ‘end-plate potentials’ of crustacean muscle fibres. J. exp. Biol. 29, 433–439.
FLEISCHER, S., OGUNBUNMI, E. M., DIXON, M. C. ANDFLEER, E. A. M. (1985). Localization of Ca2+
release channels with ryanodine in junctional terminal cisternae of sarcoplasmic reticulum of fast skeletal muscle. Proc. natn. Acad. Sci. U.S.A. 82, 7256–7259.
FORMELOVA, J., HURNAK, O., NOVOTOVA, M. ANDZACHAR, J. (1990). Ryanodine receptor purified from
crayfish skeletal muscle. Gen. Physiol. Biophys. 9, 445–453.
FRANZINI-ARMSTRONG, C., EASTWOOD, A. B. ANDPEACHEY, L. D.(1986). Shape and disposition of clefts, tubules and sarcoplasmic reticulum in long and short sarcomere fibers of crab and crayfish. Cell
Tissue Res. 244, 9–19.
GAINER, H. (1968). The role of calcium in excitation–contraction coupling of lobster muscle. J. gen.
Physiol. 52, 88–110.
GOBLET, C. AND MOUNIER, Y. (1986). Calcium-induced calcium release mechanism from the sarcoplasmic reticulum in skinned crab muscle fibres. Cell Calcium 7, 61–72.
GOVIND, C. K., BUDD, T. W. ANDATWOOD, H. L. (1981). Fiber composition and innervation patterns of the limb closer muscle in the lobster Homarus americanus. Biol. Bull. mar. biol. Lab., Woods Hole 160, 69–79.
GYORKE, S. AND PALADE, P. (1992). Calcium-induced calcium release in crayfish skeletal muscle.
J. Physiol., Lond. 457, 195–210.
HAGIWARA, S., CHICHIBU, S. ANDNAKA, K. (1964). The effect of various ions on resting and spike potentials of barnacle muscle fibers. J. gen. Physiol. 48, 163–179.
HAGIWARA, S., HAYASHI, H. AND TAKAHASHI, K. (1969). Calcium and potassium currents of the membrane of a barnacle muscle fibre in relation to the calcium spike. J. Physiol., Lond. 205, 115–129. HAGIWARA, S. ANDNAKA, K.(1964). The initiation of spike potential in barnacle muscle fibers under low
intracellular Ca2+. J. gen. Physiol. 48, 141–162.
HAGIWARA, S., TAKAHASHI, K. ANDJUNGE, D. (1968). Excitation–contraction coupling in a barnacle muscle fiber as examined with voltage clamp technique. J. gen. Physiol. 51, 157–175.
HENCEK, M. ANDZACHAR, J.(1977). Calcium currents and conductances in the muscle membrane of the crayfish. J. Physiol., Lond. 268, 51–71.
HOYLE, G., MCNEILL, P. A. ANDSELVERSTON, A. I.(1973). Ultrastructure of barnacle giant muscle fibers.
J. Cell Biol. 56, 74–91.
HYMEL, L., INUI, M., FLEISCHER, S. ANDSCHINDLER, H. (1988). Purified ryanodine receptor of skeletal muscle sarcoplasmic reticulum forms Ca2+-activated oligomeric Ca2+channels in planar bilayers. Proc. natn. Acad. Sci. U.S.A. 85, 441–445.
from skeletal muscle sarcoplasmic reticulum is the Ca2+-permeable pore of the calcium release
channel. J. biol. Chem. 262, 16636–16643.
INUI, M., SAITO, A. ANDFLEISCHER, S. (1987a). Purification of the ryanodine receptor and identity with
feet structures of junctional terminal cisternae of sarcoplasmic reticulum from fast skeletal muscle.
J. biol. Chem. 262, 1740–1747.
INUI, M., SAITO, A. AND FLEISCHER, S. (1987b). Isolation of the ryanodine receptor from cardiac
sarcoplasmic reticulum and identity with the feet structures. J. biol. Chem. 262, 15637–15642. JAHROMI, S. S. ANDATWOOD, H. L. (1967). Ultrastructural features of crayfish phasic and tonic muscle
fibers. Can. J. Zool. 45, 601–606.
JMARI, K., MIRONNEAU, C. ANDMIRONNEAU, J. (1986). Inactivation of calcium channel current in rat
uterine smooth muscle: evidence for calcium- and voltage-mediated mechanisms. J. Physiol., Lond. 380, 111–126.
KANAYA, S., ARLOCK, P., KATZUNG, B. G. ANDHONDEGHEM, L. M. (1983). Diltiazem and verapamil
preferentially block inactivated cardiac calcium channels. J. molec. cell. Cardiol. 15, 145–148. KRIZANOVA, O., NOVOTOVA, M. ANDZACHAR, J. (1990). Characterization of DHP binding protein in
crayfish striated muscle. FEBS Lett. 267, 311–315.
LAI, F. A., ERICKSON, H. P., ROUSSEAU, E., LIU, Q.-Y. AND MEISSNER, G. (1988). Purification and
reconstitution of the calcium release channel from skeletal muscle. Nature 331, 315–319.
LEE, K. S., MARBAN, E. ANDTSIEN, R. W. (1985). Inactivation of calcium channels in mammalian heart
cells: joint dependence on membrane potential and intracellular calcium. J. Physiol., Lond. 364, 395–411.
LEE, K. S. AND TSIEN, R. W. (1983). Mechanism of calcium channel block by verapamil, D600,
diltiazem and nitrendipine in single dialyzed heart cells. Nature 302, 790–794.
LOESSER, K. E., CASTELLANI, L. ANDFRANZINI-ARMSTRONG, C. (1992). Dispositions of junctional feet in
muscles of invertebrates. J. Muscle Res. Cell Motil. 13, 161–173.
MATSUMURA, M. (1978). The rate of action of calcium on the electrical and mechanical responses of the
crayfish muscle fibers. Jap. J. Physiol. 28, 75–87.
MOUNIER, Y. ANDGOBLET, C.(1987). Role of the different calcium sources in the excitation–contraction
coupling in crab muscle fibers. Can. J. Physiol. Pharmac. 65, 667–671.
MOUNIER, Y. ANDVASSORT, G. (1975a). Initial and delayed membrane currents in crab muscle fibre
under voltage-clamp conditions. J. Physiol., Lond. 251, 589–608.
MOUNIER, Y. AND VASSORT, G. (1975b). Evidence for a transient potassium membrane current
dependent on calcium influx in crab muscle fibre. J. Physiol., Lond. 251, 609–625.
NAGASAKI, K. AND FLEISCHER, S. (1988). Ryanodine sensitivity of the calcium release channel of
sarcoplasmic reticulum. Cell Calcium 9, 1–7.
ROUSSEAU, E., SMITH, J. S. AND MEISSNER, G. (1987). Ryanodine modifies conductance and gating
behavior of single Ca2+release channel. Am. J. Physiol. 253, C364–C368.
SANGUINETTI, M. C. ANDKASS, R. S.(1984). Voltage–dependent block of calcium channel current in the
calf Purkinje fiber by dihydropyridine calcium channel antagonists. Circulation Res. 55, 336–348. STRIESSNIG, J., GLOSSMANN, H. ANDCATTERALL, W. A. (1990). Identification of a phenylalkylamine
binding region within the a1 subunit of skeletal muscle Ca2+channels. Proc. natn. acad. Sci. U.S.A.
87, 9108–9112.
TAKESHIMA, H., NISHIMURA, S., MATSUMOTO, T., ISHIDA, H., KANGAWA, K., MINAMINO, N., MATSUO, H.,
UEDA, M., HANAOKA, M., HIROSE, T. ANDNUMA, S. (1989). Primary structure and expression from