Factors Affecting the Distribution of Cytoplasmic Incompatibility in
Drosophila simulans
Ary A.
Hoffmann,” Michael Turellit and LawrenceG.
Harshman**Department of Genetics and Human Variation, La Trobe University, Bundoora, 1083 Australia, tDepartment of Genetics and ‘Department of Entomology, University of California, Davis, California 95616
Manuscript received February 23, 1990 Accepted for publication August 10, 1990
ABSTRACT
In Drosophila simulans a Wolbachia-like microorganism is responsible for reduced egg-hatch when infected males mate with uninfected females. Both incompatibility types have previously been found in North America, Europe and Africa. Some California populations have remained polymorphic for over two years, and the infection is apparently spreading in central California. Egg hatch proportions for wild-caught females from polymorphic populations show that the incompatibility system acts in nature, but egg mortality rates are apparently lower than observed in laboratory populations. Although infected females maintained under various laboratory conditions never produce uninfected offspring, some wild-caught infected females produce both infected and uninfected progeny. This helps explain the persistence of a low frequency of uninfected flies in predominantly infected populations and may also explain the other polymorphisms observed. Fitness comparisons of infected and uninfected stocks, including both larval and adult fitness components, indicate that fecundity may be the component most affected. Infected females suffer a fecundity reduction of IO-20% in the laboratory, but the reduction seems to be smaller in nature. A theoretical analysis provides some insight into the population biology of the infection.
C
YTOPLASMIC incompatibility is widespread in insects, and was first described in Drosophila simulans by HOFFMANN, TURELLI and SIMMONS (1 986) in crosses between Californian populations. T h e lines previously described fell into two types, termed “R” and “W” (HOFFMANN and TURELLI 1988). W females are incompatible with R males, in the sense that a significant fraction of the eggs produced by such matings fail to hatch. In contrast, the reciprocal cross and crosses within compatibility types all produce comparable numbers of progeny. Laboratory studies indicate that type R mothers produce only type R offspring, whereas matings between R males and W females produce 1-2% R offspring (HOFFMANN andTURELLI 1988). Type
R
strains can be converted totype W by culturing them on medium containing tetracycline, suggesting that incompatibility is caused by a microbial infection in the R strains. In support of this hypothesis, LOUIS and NICRO (1 989) and BIN- NINGTON and HOFFMANN (1989) found that a Wolba- chia-like microorganism is present in the gonadal tis- sue of R strains but absent from W strains.
Both incompatibility types appear to be widespread throughout the world, and have been recorded from Africa and Europe as well as the east and west coasts of North America (HOFFMANN and TURELLI 1988; our unpublished data). In contrast, extensive sampling has not uncovered R strains in Australia. Both incom- patibility types can also be found in collections from
<;cne[ic.s 126: 933-948 (December, 1990)
the same site, at least in California (HOFFMANN and TURELLI 1988) and France (J. R. DAVID, personal communication). Polymorphism is not predicted by simple models for the population dynamics of incom- patibility factors. If cytoplasmic incompatibility is un- idirectional, such that uninfected females are incom- patible with infected males, uninfected females will produce fewer adult offspring than infected females, and their disadvantage increases as the infection be- comes more common. Thus, the infection is expected to spread from its center of origin and polymorphisms for the two types should not persist (CASPARI and WATSON 1959).
Our population data suggests that other factors must contribute to the frequency of incompatibility types. Some possibilities are summarized in Table 1 and are addressed by the experiments presented be- low. Perhaps the simplest explanation for persistent polymorphism is that incompatibility is expressed in the laboratory but not under field conditions. We have tested this possibility by scoring egg hatch for wild-caught R and W females from polymorphic pop- ulations.
934 A. A. Hoffmann, M. Turelli and L. G. Harshman
TABLE 1
Factors that may help to explain the patchy geographical distribution and polymorphism of incompatibility t y p e s in D.
simulans
1 .
2.
3.
4.
5.
6.
7 .
Incompatibility between R and W types is not expressed under
Occasional W progeny from R mothers
a. Curing of infection by naturally occurring antibiotics b. Curing of infection by exposure to extreme temperatures Deleterious viability or fecundity effects of the incompatibility-
Nonrandom mating of incompatibility types
Effects of incompatibility type on female remating speed or
Migration between populations nearly monomorphic for R or
Interactions between nuclear eenes and the infection field conditions
causing infection
sperm competition
w
that we have not tested may cause segregation in the field. In particular, exposure to high temperatures may be involved because high temperatures suppress incompatibility (HOFFMANN, TURELLI and SIMMONS
1986); and age may be involved because old R males tend to have fewer Wolbachia-like organisms and are more compatible with W females (HOFFMANN, TUR- ELLI and SIMMONS 1986; BINNINCTON and HOFFMANN 1989). Segregation may also occur when larvae en- counter naturally occurring antibiotics, as demon- strated in Tribolium (STEVENS 1989). We have tested for segregation in the laboratory and with field-col- lected females.
Third, the dynamics of the infection may be influ- enced by deleterious effects on fitness. Theoretical analyses show that such effects produce an unstable equilibrium frequency, below which the infection is lost, but above which it spreads to fixation (CASPARI and WATSON 1959; FINE 1978). We have previously found suggestive evidence for deleterious fitness ef- fects by comparing the number of progeny produced by R strains to the number produced by derivative strains treated with tetracycline to cure the infection (HOFFMANN and TURELLI 1988). These results are only suggestive because tetracycline may affect fitness- related factors other than the incompatibility system. Also, we did not previously examine fitness compo- nents other than productivity. We have therefore carried out tests on a wider range of fitness compo- nents using comparisons other than those involving tetracycline-treated strains, and we have also obtained fecundity data from co-occurring R and W flies from the field.
A fourth possibility is that incompatibility dynamics are influenced by mating behavior or sperm compe- tition. Mating between R and W flies may be non- random; for example, W females may avoid mating with infected males. We have presented evidence against this in virgin matings (HOFFMANN and TUR-
ELL1 1988); here we describe experiments with previ- ously inseminated females, because such females mate less readily and may therefore be more discriminating. Incompatibility dynamics may also be influenced by other aspects of mating that affect fitness. W females may remate more quickly than R females after mating with R males. Previous experiments with Drosophila melanogaster have found that the tendency of females to remate depends on properties of both sexes (PYLE and GROMKO 1981). Remating speed is particularly dependent on the amount of sperm stored by D. melanogaster females (GROMKO, NEWPORT and KOR- TIER 1984). Thus, if the Wolbachia-like infection af- fects sperm production or transfer, it may influence remating in D. simulans. Incompatibility dynamics could also be altered by differential sperm competi- tion if the incompatibility mechanism involves factors that act prior to syngamy. We have therefore consid- ered the remating dynamics of R and W females and the effects of prior mating with an R or W male on the progeny produced after a second mating.
Two other factors not considered in our experi- ments may also influence the distribution of incom- patibility types. These are migration and nuclear- cytoplasmic interactions. Migration may account for persistent polymorphisms of incompatibility types if there is an unstable equilibrium frequency within populations and populations nearly fixed for the al- ternative types exchange relatively few migrants
(4.
KARLIN and MCGRECOR 1972; NICRO and PROUT1990). Nuclear factors may reduce the level of incom- patibility in crosses between W females and R males, although we have so far failed to find evidence for nuclear-cytoplasmic interactions. Both of these factors are considered in the DIxUSSION.
Before describing experiments concerning these factors, we briefly consider some recent California population surveys that supplement the data on the distribution of R and W in HOFFMANN and TURELLI (1 988). We also present data on the frequency dynam- ics of the incompatibility types in laboratory popula- tions set up with lines from a persistently polymorphic field population.
MATERIALS AND METHODS
Stocks and field collections: Several collections at differ- ent times were made from citrus groves near Piru in a valley separated by the Santa Susana mountains from the Los
TABLE 2
Designation of stocks used in the laboratory experiments
Code Origin Description
R stocks
RH Highgrove 1985 Massbred from 18 isofemale lines RP Piru 1988 Massbred from 20 type R isofemale lines
RPM Piru/Melbourne RP backcrossed to WM males for 5 generations RR Riverside 1984 Massbred from 30 isofemale lines
R R W Riverside 1984 White eye stock derived from a wild-caught white eyed female
Rscgs) Sanger 1985 Isofemale line
W stocks
W H ( J 3 ) Highgrove 1985 Isofemale line
WHT Highgrove 1985 Derived from R H by tetracycline treatment
WM Melbourne 1985 Massbred from 25 isofemale lines
WP Piru 1988 Massbred from 20 type W isofemale lines
W P M Piru/Melbourne W p backcrossed to WM males for 5 generations W R T Riverside 1984 Derived from RR by tetracycline treatment
W R T ( w ) Riverside 1984 Derived from R R ~ ~ ) by tetracycline treatment
WS Sanger 1985 Massbred from 18 isofemale lines
ww
Watsonville 1984 Massbred from 30 isofemale linesquency distribution of the incompatibility types.
Several different stocks were used in the laboratory ex- periments, and these are listed in Table 2 together with their designations. We established two independent R stocks (RIB) and two independent W stocks (Wp) from Piru. Each was initiated with the progeny of 20 isofemale strains from the November 1988 collection. We also used several of the massbred stocks described in HOFFMANN and TURELLI (1988). These include the R stocks Riverside (RR) and High- grove (RH), and the W stocks Melbourne (WM) and Sanger (Ws). We also used an R isofemale strain from Sanger (RS(LS)) and a W isofemale strain from Highgrove (WH(SS)), referred to as S13 and Hg33, respectively, in HOFFMANN and TUR-
ELLI (1988). Finally, we performed remating experiments with a white-eye mutant stock, RR+), isolated from a River- side field collection in 1984. Other stocks were derived from these by treating R stocks with tetracycline to cure the infection and by backcrossing to WH males. All flies were cultured on standard Drosophila medium at 20-23" unless otherwise stated.
Frequency of incompatibility types: We sampled the population at Piru on four occasions over 3 years, while most other sites were sampled only once o r twice. Females from the field were set up individually in vials. T h e incom- patibility type of the resulting isofemale line was determined by pair mating 3-5 male progeny to virgin type W females (W, or Ww) and 3-5 virgin female progeny to type R males (RR). Pairs of flies 0-3 days old were placed in vials and left to oviposit for 2-3 days. For vials with at least ten eggs, we scored the proportion of eggs that hatched 25-48 hr after the adults were removed. Crosses were considered incom- patible when <25% of the eggs hatched, and compatible when >75% hatched. In hundreds of crosses, we have found that more than 95% of our control crosses with R and W stocks maintained in the laboratory produce unambiguous results with this criterion. Exceptions almost always involve compatible crosses that produce egg-hatch rates below 75%. Results from newly established isofemale lines are generally consistent for the two types of crosses, L e . , egg hatch is consistently low either when the strain females are mated to R males or when the strain males are mated to W females. We have found no strains in which either both or neither of these crosses produced consistently low egg hatch. How- ever, we have found some newly established isofemale lines
that yield inconsistent results, i.e., the replicate crosses pro- duce inconsistent results or produce intermediate (25-75%) proportions of hatched eggs. These are not counted in our estimates of R and W frequencies but are discussed sepa- rately.
Incompatibility levels in the field T h e first experiment was carried out with females collected at Piru and Lake Cachuma in July 1988. Females and males were taken to the laboratory after having been stored in vials for 1-2 days following the field collection. Females were set up individ- ually on spoons containing grape juice concentrate and Drosophila medium, and covered with a live yeast suspen- sion. Females were transferred to fresh spoons after 24 hr. After another 24 hr of egg laying, females were transferred to standard food vials to establish isofemale lines. T h e number of hatched and unhatched eggs on each spoon was scored >24 hr after the female was removed. Each isofemale line was characterized for incompatibility type as described above.
In the second and third experiments (November 1988 and July 1989), flies were only collected from Piru. T h e sexes were separated in the field immediately after collection to prevent mating in the storage vials. Remating in storage vials could have biased the estimate of incompatibility in the first experiment, because compatibility is increased as R males get older.
In each of these experiments, we protected the flies from heat stress during transport to the laboratory by shielding them from the sun and using air conditioning in the summer. During our summer collections, the temperature at the collection sites approached 37'. Our laboratory tests were performed at room temperature, 20-23 O .
936 A. A. Hoffmann, M. Turelli and L. G. Harshman
virgin females from each population were crossed to RK males, and incompatibility type was determined from the proportion of the eggs that hatched. We also included crosses of Rp males to Rp females to determine the percent- age of vials that may have appeared incompatible because of poor hatchability not associated with incompatibility.
Effects of male age: Previous experiments had indicated that the age of R males influenced levels of incompatibility, while the age of W females had no effect (HOFFMANN, TURELLI and SIMMONS 1986). We examined the effect of male age and temperature in more detail by crossing R p males that were 1-24 days old to Wp females that were 2- 6 days old. In the first experiment, males were collected every day for 24 days and aged at 20" before crossing. In the second experiment, males were aged at 28" and only males aged for 1, 3, 6, 9, 12, 15, 18 and 21 days were used. T o determine incompatibility, pairs of males and females were placed in vials with a spoon as described above for the characterization of field incompatibility levels, except that the grape-juice medium was replaced by a black currant medium. Twenty replicate vials were set up for each male age class. Control crosses (W, X Wp and Rp X Rl,) were also set up in a separate experiment with males that were 1 , 12 and 24 days old (20") or males that were 1, 12 and 21 days old (28"). Males were crossed to females that were 2-6 days old and ten replicates were set up for each cross.
Segregation experiments: Three different experiments were performed to determine conditions under which R females may produce W offspring.
High temperature: RK flies were cultured at 28", a temper- ature that causes a temporary suppression of incompatibility (HOFFMANN, TURELLI and SIMMONS 1986). Female progeny were collected as virgins and crossed to WM males. These flies were placed at 28" again, and F1 progeny were back- crossed to WM males. This procedure was repeated for another three backcross generations. Any type W female progeny that emerge should persist in the population be- cause they are compatible with WM males. T o test for segregation at the end of the experiment, females were set up individually in vials and progeny were tested for incom- patibility type at room temperature as described above.
Aged type R females: At room temperature, compatibility is restored to a large extent when R males older than 2 weeks are crossed to type W females (HOFFMANN, TURELLI and SIMMONS 1986), suggesting that the level of infection may be reduced. This could also occur in old females and may lead to segregation. Using R R flies, we therefore set up 198 isofemale lines from the progeny of 30 females 3 weeks old, as well as 196 isofemale lines from females 2-4 days old. Progeny were crossed to WM males in establishing the isofemale lines to ensure that any type W segregants would produce offspring.
Field segregation: In the third Piru collection, progeny from field-collected females that had been identified as type R were tested for incompatibility type by crossing females to RK males. We tested 10 progeny from 44 females and established an isofemale line for each of the progeny to verify their incompatibility type. We also tested lines derived from a female (designated 34) that could not be assigned to an incompatibility type because the level of incompatibility in crosses with R K males was intermediate.
Fitness experiments: T o examine the effects of incom- patibility type on fitness and mating, we attempted to com- pare R and W stocks with similar nuclear backgrounds. In earlier experiments (HOFFMANN and TURELLI 1988), we compared stocks that had been cultured for a generation on medium containing tetracycline (0.03%) with parental R stocks that were not exposed to tetracycline. T h e antibiotic
treatment converts R stocks to W stocks (HOFFMANN, TUR- ELLI and SIMMONS 1986), but may alter cellular components other than the incompatibility factor. In an alternative ap- proach, R and W stocks were crossed reciprocally, and fitness comparisons were made between the Fls. The incom- patibility factor is largely inherited via the maternal parent (HOFFMANN and TURELLI 1988), so that Fls will have the incompatibility type of their maternal parent. The nuclear background of the FI females will be identical, although males will have an X chromosome and the mitochondrial from the maternal stock. Our third approach involved com- parisons of the Piru stocks Rp and Wp. Replicate R and W stocks from Piru had been set up from different sets of isofemale lines. These stocks should have similar nuclear backgrounds; but they differ in their mitochondrial ge- nomes because of an association between mtDNA variation and incompatibility type in this population (HALE and HOFF- MANN 1990). In a fourth approach, we made the nuclear background of R and W lines similar by backcrossing lines to the same laboratory stock for several generations.
Egg-to-adultjtness components: These experiments tested the effects of incompatibility type on egg-to-adult viability, development time and biomass of the emerging adults. Comparisons were made between tetracycline-treated and untreated lines from the Riverside (RR) and Highgrove (RI,) stocks, and between the mass-bred RP and Wp lines from Piru. T h e tetracycline-treated stocks were obtained by cul- turing flies for one generation on medium with tetracycline as described in HOFFMANN and TURELLI (1988), followed by two or three generations on medium without tetracycline. The treated stocks will be referred to as WKI. (Riverside) and W k I . , (Highgrove). Two WK-r lines and two WH,I. lines were set up after tetracycline treatment, and the fitness of these lines was compared to two RK or two REI lines set up at the same time. Twenty eggs from a line were transferred to vials containing 10 ml of laboratory medium. Twenty vials were set up per line. Adult progeny were collected daily and held in vials until all progeny had emerged, and flies were then weighed on a microbalance.
Fecundity: The effect of incompatibility type on fecundity was examined by comparing tetracycline-treated and un- treated lines from Riverside used in the egg-to-adult fitness experiment. We also tested R and W stocks from Piru that had been backcrossed to Melbourne males for five genera- tions, these are referred to as R p M and WPM, respectively. Males and females were collected as virgins within a 12-hr period. These flies had been cultured at a low density by placing groups of 20 eggs in vials with 10 ml of medium. R and W males and females were crossed in all possible com- binations so that any fecundity effects could be assigned to male and female incompatibility types. Because two replicate R and W stocks were tested in each comparison, there were 16 combinations. Twenty replicate pairs of males and fe- males were crossed for most combinations, although some vials were discarded because of mortality or escape during transfer.
FIGURE 1 .-Map of California indicating collection sites for sam- ples in Table 3.
combinations randomized within the 20 blocks.
Mating experiments. Two different experimental re- gimes were used to assay different aspects of mating per- formance.
Muting success in large cages: Experiments were carried out in Perspex cubic cages 400 mm on a side. Two sides of the cages were open and covered with gauze. Eight plastic cups with laboratory medium were placed at the bottom of each cage. Rand W lines were tested pairwise. Two hundred males and 200 females were released into each cage. These flies had been aged together for 3 days at a density of 20-
30 flies per vial. The two lines to be tested were marked with a different color (green or pink) of micronized fluores- cent dust prior to release. Flies were introduced into cages in the evening and left to settle for 12 hr. Mating pairs were removed at 1-hr intervals for a 12-hr period and were identified under UV light by their dust colors. We tested four pairs of R and W stocks. We compared the Fls from reciprocal crosses between Rand W lines from Sanger (Rs(ls) and Ws) and Highgrove (RH and WH(33)) as well as Fls from crosses between RR and WM. We also compared the mating success of Rp and Wp.
Remating success: T o examine the effect of male incom- patibility type on remating behavior, approximately 200 W females were first mated to R or W males when flies were
2-3 days old. Females were individually mated in vials so
that males could be removed within 30 min after copulation was completed. Females that mated were held in vials with laboratory medium and live yeast for 2 days. This medium stimulates oviposition which increases the receptivity of fe- males for remating. Females were paired with a second R
or W male in fresh vials and observed for 12-14 hr, at which stage about half had remated. The effect of male incompatibility type was examined by comparing the num- ber of females that had or had not remated for each com- bination of first and second males. This experiment was carried out with the R and W Fls from reciprocal crosses between R and W stocks established from single populations: the Sanger stocks (W, and RXls)), and the Highgrove stocks (RH and WH(s3)). It was also carried out with the mass-bred
TABLE 3
Distribution of R and W lines in 1987-1989 collections from California
Date Location #R #W freq(R) 95% CI“
6/1987 Highgrove 26 1 0.96 (0.81, 0.999)
Ventura 10 2 0.83 (0.52, 0.98)
Lake Cachuma 7 12 0.37 (0.16, 0.62)
Piru 9 8 0.53 (0.28, 0.77)
Arvin 0 20 0.00 0.14
Sanger 0 7 0.00 0.35
7/1988 Lake Cachuma 47 57 0.45 (0.35.0.55)
Piru 38 14 0.73 (0.59, 0.84)
11/1988 Highgrove 9 0 1.00 0.72
Piru 76 97 0.44 (0.36, 0.52)
Davis 0 12 0.00 0.22
7/1989 San Diego 38 2 0.95 (0.83, 0.99)
Highgrove 245 15 0.94 (0.91, 0.97)
Piru 44 31 0.59 (0.47, 0.70)
9/1989 Sanger 28 77 0.27 (0.19, 0.36)
11/1989 Ventura 13 1 0.93 (0.66, 0.998)
Arvin 19 46 0.29 (0.19, 0.42)
‘ “Exact” confidence intervals with probability 0.025 in each tail were computed directly from the binomial. When freq(R) equals
0.0 (1.0). the upper (lower) 5% value is given.
Rp and Wp stocks from Piru. Thus flies from predominantly W, predominantly R, and highly polymorphic populations were analyzed. The W Fls were obtained by crossing old R males to W females ($. HOFFMANN and TURELLI 1988).
Sperm competition: We used a Riverside stock homozy- gous for a recessive white eye gene (RR(J to investigate the effects of incompatibility type on sperm competition. White eye flies that were type W (WRT,,)) were obtained by tetra- cycline treatment. White eye virgin females that were type W were mated to wild type males (2-3 days old) that were either type W (WRT) or R (RR). The WRT males were from the Riverside stock that had been treated with tetracycline for one generation. Males and females were set up in pairs
so that mating could be monitored. Males were removed within 30 min after we observed mating. Females were then held individually in vials with live yeast for 4 days before they were exposed to a white eye male that was type R (RR(,,,)) or W (WRT(,)). Most of the females (>60%) were observed to remate within 12 hr. Males were removed within 30 min after we observed remating and females were transferred to fresh yeasted vials. Females were subse- quently transferred to new vials every 3 days until no more progeny were produced. The number of white eye and wild type progeny that emerged from eggs laid after the second mating were scored.
RESULTS
938 A. A. Hoffmann, M . Turelli and L. G. Harshman
tions. However, a contingency test indicates signifi- cant heterogeneity in the four Piru collections ( G =
15.48, d.f. = 3, P
<
0.002); and the difference be- tween the two 1988 collections was also significant ( G= 14.02, d.f. = 1) even though they were separated by only 4 months.
We failed to collect any R strains in the Central Valley in 1987, but we collected several R strains in large samples from Sanger and Arvin in 1989. T h e confidence intervals indicate that the frequency dis- tribution of R and W was not significantly different in the 1987 and 1989 Sanger collections, but the 1987 sample is quite small. However, the 1989 Sanger
collection does differ significantly from the combined 1987 Central Valley collections from Arvin and San-
ger ( G = 14.64, d.f. = 1, P
<
0.001). The 1987 and1989 Arvin collections do differ significantly (G =
11.78, d.f. = 1 , P
<
0.001), but the 1989 collections from Arvin and Sanger do not.In agreement with earlier results (HOFFMANN and TURELLI 1988), we found a low frequency of W strains in the predominantly R populations at Highgrove and Ventura. A 1989 sample from San Diego gave similar results. There are no statistically significant differ- ences among the six samples from Highgrove, Ven- tura and San Diego. Pooling them, we find that the frequency of R in these southern California popula- tions is 0.94, with a 95% confidence interval of 0.91- 0.96.
In the 1989 Arvin sample, 83 isofemale lines were successfully tested using the F1 from field-collected females. For each line, Si, we set up three replicates of each test cross, Si? X RRd and WW? X Sid. From these, 65 lines could be assigned to an incompatibility type using the criteria that all replicates of each cross were consistent, and only one of the two types of crosses was compatible. The remaining 18 lines could not be assigned to an incompatibility class because of inconsistency among the replicates. Isofemale sublines have been set up from these ambiguous lines for further analysis.
Incompatibility in the field: In the first and fourth field collections, most females laid only a few eggs in the first 24-hr period and counts were pooled over the 2 days. In the second and third field collections, most fenlales laid more than ten eggs in the first 24- hr period, and egg counts were therefore not made in the second laying period. One-tailed t-tests were used to test for a deleterious effect of the infection on fecundity. The proportion of eggs that hatched showed a skewed distribution because all eggs hatched in many cases. Non-parametric Kruskal-Wallis tests were therefore used to test for a decrease in the proportion of hatched eggs in vials with W females. Proportions were calculated only for females that laid ten or more eggs.
There was no statistically significant effect of incom- patibility type on the proportion of eggs that hatched in the Lake Cachuma collection (Table 4). In contrast, W females produced significantly higher proportions of unhatched eggs in all three Piru collections, with differences in the proportion unhatched ranging from
13% to 28%. These results provide clear evidence that cytoplasmic incompatibility is expressed under field conditions at Piru.
We can use the hatch rates for field-collected fe- males to estimate the relative hatch rate produced by incompatible ( i e . , W? X
Rd)
vs. compatible crosses(ie., R X R, W X W, or R? X Wd). Assuming that all
three compatible crosses produce the same average hatch rate, let Hr; denote the average hatch rate from compatible crosses, and let HI denote the average hatch rate from incompatible crosses. If the two in- compatibility types mate at random in the field, the relative hatch rate from incompatible crosses can be estimated as
where
flu.
andRK
denote the average hatch rates from field-collected W and R females andp
denotes the frequency of R in the population. This expression holds irrespective of multiple mating, it simply re- quires that the fraction of eggs fertilized by R sperm isp ,
the fraction of R adults. Table 5 presents two estimates of H for each field experiment as well as approximate 95% confidence intervals based on the bias-corrected percentile method with 1000 bootstrap samples (EFFRON 1982, Sec. 10.7). T h e two estimates of H for each experiment result from computing the averagesRW
andf
l
,
in (1) in different ways. T h e “unweighted averages” are obtained by calculating a hatch rate for each female that produced ten or more eggs, then averaging these hatch rates for R and W females. T h e “weighted averages” are obtained by pooling all the eggs laid by females of the same incom- patibility type (irrespective of the number laid), then calculating the fraction of all of these that hatched. As shown in Table 5, both averages give very similar results.TABLE 4
Incompatibility levels and number of eggs laid (with sample size N) by R and W females collected from the field
Type R T y p e w
Collection rf(S0) N X(SD) N P
Lake Cachuma 7/88
Number of eggs 27.1 (13.8) 47 32.8 (17.3) 57 C0.05
Proportion hatched 0.95 (0.07) 42 0.91 (0.12) 52 NS
Number of eggs 40.6 (1 1.9) 38 41.2 (20.8) 14 NS
Proportion hatched 0.95 (0.06) 38 0.82 (0.20) 14 co.001
Number of eggs 29.0 (9.8) 76 29.9 (8.5) 97 NS
Proportion hatched 0.98 (0.05) 75 0.77 (0.29) 96 co.001
Number of eggs 42.0 (16.8) 44 46.6 (14.4) 31 NS
Proportion hatched 0.97 (0.04) 41 0.69 (0.36) 31 co.001
Piru 7/88
Piru 1 1/88
Piru 8/89
Probabilities are for one-tailed t-tests (number of eggs) or Kruskal-Wallis tests (proportion hatched).
TABLE 5
Inferred relative egg hatch proportions (H) from incompatible matings in the field
Unwei hted H Weighted H
Collection ( 9 5 8 CY) (95% CI)
Lake Cachuma 7/88 0.91 (0.83, 1.00) 0.91 (0.82, 0.99)
Piru 7/88 0.81 (0.63, 0.95) 0.85 (0.72, 0.97)
Piru 11/88 0.52 (0.33, 0.66) 0.53 (0.34, 0.68)
Piru 8/89 0.50 (0.22, 0.71) 0.57 (0.29, 0.77)
Based on the bias-corrected percentile method with 1000 boots- trap samples. If we weight each of these estimates by the inferred number of incompatible m_atings, the average H using weighted (unweighted) estimates of HW and HR equals 0.65 (0.67) with the Lake Cachuma data and 0.56 (0.59) without the Lake Cachuma data.
T h e relative productivities (numbers of eggs laid) for R vs. W females in the four samples range from 0.83 to 0.985 and average 0.92.
Changes of incompatibility-type frequency in lab- oratory populations: These experiments tested
whether type R increased in frequency as expected when populations were started at intermediate fre- quencies similar to those found at Piru, using the mass-bred Rp and WP stocks from this location. When flies were not aged prior to being transferred to fresh bottles, the R incompatibility type increased in all ten populations after five generations, as shown by the increased frequency of compatible matings (Figure 2).
In nine of the ten populations, an additional increase was observed after ten generations. For populations maintained with aged flies, eight of the ten popula- tions showed an increase in the frequency of compat- ible matings after five generations. T h e two popula- tions that did not show an increase were maintained for a further five generations, and the number of compatible matings increased to 92% and 86% after ten generations. We did not expect all crosses to be “compatible” even when R had gone to fixation, be- cause vials with females that had not mated or that
”Ol
0.91
0.44
0 5 10
generation
FIGURE 2.-Changes in the frequency of compatible crosses in laboratory populations started with 50% R and 50% W flies from mass-bred Piru stocks, R p and Wp. The solid (open) squares indicate populations in which 0-1-day-old (8-9-day-old) flies were used to set up each generation. The initial frequency of compatible matings is estimated from the controls.
940 A. A. Hoffmann, M. Turelli and L. G . Harshman
-.-
0 5 IO 25 20 15
0 5 10 25 20 15
male age (days)
FIGURE 3.-Fraction of eggs that hatch from Wp? X R& crosses set up with males of different ages. The solid squares indicate the means for these incompatible crosses. The open circles (squares) indicate the means for the Rp X Rp (Wp X WP) controls. The error bars are experimental standard deviations, not standard errors of the means.
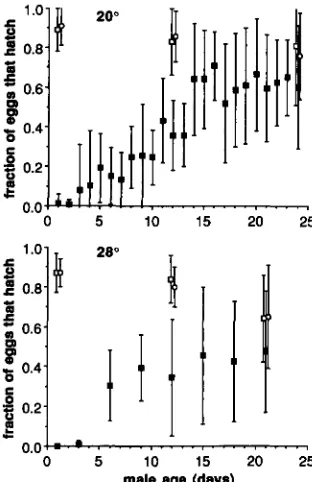
Effects of male age and temperature: T h e propor- tion of eggs that hatched was calculated for all females that laid ten or more eggs. Proportions are plotted against male age in Figure 3. Data points are based on at least 12 females. T h e proportion of eggs that hatched increased with male age as expected. For males stored at 20
"
, hatchability increased gradually and appeared to level out at 50-70% when males were two or more weeks old, although the error bars (which provide standard deviations, not standard errors for the means) indicate that the level of incompatibility varied enormously from vial to vial. For the males stored at 28", the proportion of eggs that hatched increased more quickly but appeared to level out at an earlier age at around 40%.In the Wp X Wp control crosses with males stored at 20", males that were 1, 12 and 24 days old pro- duced mean hatchabilities of 0.89 (SD = 0.1 l), 0.83 (0.17) and 0.81 (0.30), respectively, while the mean hatchabilities for the Rp X Rp control crosses with males that were 1, 12 and 21 days old were 0.9 1 (SD
= 0.10), 0.86 (0.13) and 0.76 (0.22), respectively. Both controls suggest a decrease in hatchability when older males are used, although only the 1 us. 21-day- old comparison at 28" was statistically significant (P
<
0.05). Figure 3 shows that the control hatch rates obtained with males that are 1 or 12 days old fall outside the experimental standard deviations obtained from the incompatible crosses, but those with the 24- day-old males do not. T h e Wp X Wp crosses withmales stored at 28" produced mean hatchabilities of 0.87 (SD = ().lo), 0.84 (0.12) and 0.64 (0.22), respec- tively for males that were 1, 12 and
2
1 days old; the corresponding figures for the Rp X Rp crosses were 0.87 (SD = 0.07), 0.80 (0.10) and 0.65 (0.26), respec- tively. T h e means for 21-day-old males fall within the empirical standard deviations of the means from the incompatible crosses, while means for the younger males do not (Figure 3).These results suggest that egg hatchability in incom- patible crosses approaches control levels when females are crossed to older males, although a low level of incompatibility probably persists even when males are 3 weeks old. Averaging the results for the Rp X Rp and Wp X Wp control crosses, the relative hatch rate for the incompatible crosses is 0.77 for 24-day-old males at 20" and 0.74 for 2 1-day-old males at 28". In contrast, with 1-day-old males, the relative hatch rate from incompatible crosses is 0.014 at 20" and 0.002 at 28", so that incompatibility is virtually complete; and with 12-day-old males, the relative hatch rate is 0.42 at both 20" and 28". Thus, once hatch rates are normalized by the controls, adult temperature seems to have little effect on incompatibility.
Segregation experiments: None of the laboratory experiments provided evidence for segregation of type W flies from the RR stock. In the high tempera- ture experiment, all 200 lines established from flies after five generations at 28" were type R. In the experiment with old females, all of the 240 lines were type R, indicating that old RR females did not produce type W progeny. We have carried out other experi- ments using heat shock (35") and culture under cold temperatures with similar results. T h e incompatibility factor is therefore transmitted with complete fidelity via the maternal lineage under a range of conditions. From our 1989 Piru collection, we identified four females out of 44, initially characterized as type R, that produced both R and W type progeny. These females produced a single W progeny from the 9-10 progeny that we tested. Isofemale lines were estab- lished from all progeny characterized as W, and the incompatibility type of these lines was determined from crosses to RR males and WM females in the next generation. All four lines were found to be W, as expected. However, when retested after another two generations, one of these lines was characterized as R. Further switches in incompatibility type were not de- tected in subsequent generations.
TABLE 6
Egg-to-adult fitness components for incompatibility types
Number of Development Weight per
Comparison progeny time (days) fly (g x IO")
Highgrove (tetracycline) Means (SD)
RH(line 1 ) 16.3 (4.0) 12.74 (0.30) 10.96 (0.67) RH(line 2) 14.6 (4.0) 12.65 (0.28) 11.07 (0.68) WKT(line 1 ) 16.2 (4.5) 12.65 (0.34) 10.95 (0.74) WRT(line 2) 15.3 (4.5) 12.65 (0.35) 10.89 (0.69) ANOVAs (mean squares)
Incompatibility type (d.f. = 1) 2.00 0.24 0.17 Replicate line (d.f. = 2) 17.49 0.13 0.07 Error (d.f. = 70) 17.96 0.10 0.48 Riverside (tetracycline)
Means (SD)
RR(line 1) 14.7 (4.2) 12.79 (0.36) 10.63 (1.31) RR(line 2) 17.6 (2.4) 12.54 (0.23) 10.91 (1.29) WRT(line 1) 16.6 (2.7) 12.82 (0.40) 11.07 (1.74) WRT(line 2) 16.8 (3.3) 12.47 (0.25) 10.51 (1.26) ANOVAs (mean squares)
Incompatibility type (d.f. = 1 ) 5.30 0.01 0.00
Replicate line (d.f. = 2) 42.41 0.92*** 1.94
Error (d.f. = 70) 10.50 0.10 1.99
Piru (mass bred) Means (SD)
Rp(line 1) 15.2 (1.7)
Rp(1ine 2) 16.4 (2.1)
Wp(line 1) 15.7 (1.8)
Wp(line 2) 16.6 (2.1)
ANOVAs (mean squares)
Incompatibility type (d.f. = 1) 3.10 Replicate line (d.f. = 2) 11.13 Error (d.f. = 75) 3.89
10.93 (0.72) 11.02 (0.68) 1 1.67 (0.76) 1 1.29 (0.65)
5.10 0.75 0.47
*** P < 0,001. Means are based on 20 vials.
were assigned as W and three lines were assigned as R on the basis of crosses to RR males and WM females. One of these W lines had switched to R when lines were retested two generations later. N o further switches in incompatibility type were detected in later generations.
T o test for segregation in these flies under labora- tory conditions, we pooled the R progeny from each of the four females that produced one W progeny to establish four lines. Thirty females were collected from each line and mated individually to WM males. Progeny from the females were tested for incompati- bility type, and all were found to be R. This indicates that R females originating from Piru do not produce W progeny after several generations of laboratory culture, in agreement with earlier results with a Riv- erside stock (HOFFMANN and TURELLI 1988). This was confirmed by testing 100 isofemale lines from one of the mass-bred Rp stocks, all were found to be R.
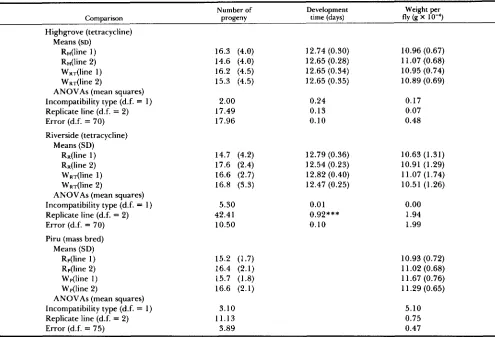
Fitness experiments: Egg-to-adult-jtness: Results from these experiments are summarized in Table 6. ANOVAs indicate that there was no statistically sig- nificant effect of incompatibility type on any of these
fitness components, regardless of whether W lines from a tetracycline treatment or Wp lines were used in the comparison. Using a t-test, we find that these experiments had the power to detect a viability differ- ence of about 10% (15%) with probability 0.5 (0.8)
once data from the replicate lines are pooled. It should be noted, however, that our experimental regime, with 20 eggs per vial, provides minimal competition.
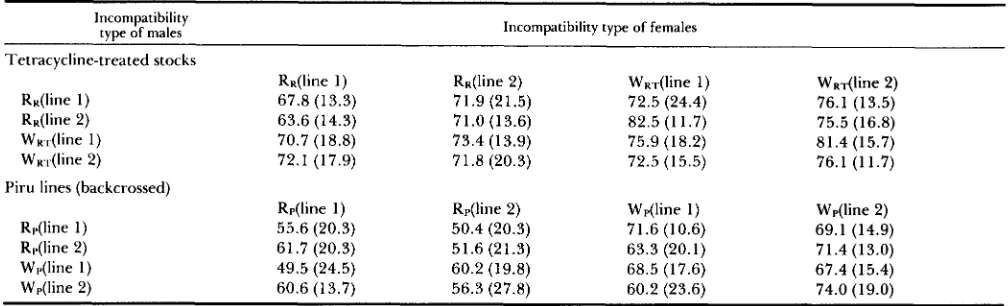
Fecundity: Mean egg counts for each combination
of R and W lines (Table
7)
suggest that R females produced fewer eggs than W females in comparisons with tetracycline-treated lines from from Riverside and with Piru lines backcrossed to WM males, regard- less of the incompatibility type of the male. This was tested with mixed-model ANOVAs (Table 8) using the modely . . y k l
-
-
a+
bi+
cj+
d h ( i )+
et(j) + f j+
gm+
h z ( i j h l m ) ,where a is the grand mean, bi the term due to female incompatibility type, c, the male incompatibility term,
drc,) and el(,) the nested replicate line terms, f j the
interaction between male and female incompatibility types, g , the block term and Itn(+,) the error term.
942 A. A. Hoffmann, M. Turelli and L. G . Harshman
TABLE 7
Mean fecundities (and standard deviations) from reciprocal crosses between R and W lines
Incompatibility
type of males Incompatibility type of females
Tetracycline-treated stocks
RK(line 1) RK(line 2) WK.,(line 1)
WKr(line 2)
RK(line 1) RR(line 2) WRT(line 1) WRT(line 2)
67.8 (13.3) 71.9 (21.5) 72.5 (24.4) 76.1 (13.5)
63.6 (14.3) 7 1 .O (1 3.6) 82.5 (1 1.7) 75.5 (16.8)
70.7 (18.8) 73.4 (1 3.9) 75.9 (18.2) 81.4 (15.7)
72.1 (17.9) 7 1.8 (20.3) 72.5 (15.5) 76.1 (11.7)
Piru lines (backcrossed)
Rp(line 1) Rp(line 2) Wp(line 1) Wp(line 2)
R,.(line 1) 55.6 (20.3) 50.4 (20.3) 71.6 (10.6) 69.1 (14.9)
R,.(line 2) 61.7 (20.3) 51.6 (21.3) 63.3 (20.1) 71.4 (13.0)
Wp(line 1) 49.5 (24.5) 60.2 (19.8) 68.5 (17.6) 67.4 (15.4)
Wp(line 2) 60.6 (13.7) 56.3 (27.8) 60.2 (23.6) 74.0 (19.0)
Means are based on 17-20 vials.
TABLE 8
ANOVAs for fecundity data from reciprocal crosses between R
and W lines
Mean squares Effect
Tetracycline-treated d.f. lines Piru lines
Female type 1 2,917.1" 12,041.7'
Male type 1 152.2 5.7
Repl. lines (female type) 2 251.7 450.2
Repl. lines (male type) 2 126.7 42.7
Block 19 410.3* 559.4
Female X male type 1 237.3 213.3
Error 2791282 246.5 367.0
* P < 0.05.
a Female incompatibility type is not significant (P < 0.10) when
tested over replicate line term, but highly significant ( P < 0.001)
when tested over pooled error and replicate line terms.
*
Female incompatibility type is significant ( P < 0.05) when tested over replicate line term and highly significant ( P < 0.001) when tested over pooled error and replicate line terms.fixed effects, while replicate line was treated as a random effect. T h e effect of female incompatibility type was significant when tested over the replicate line term in the Piru line comparison, and was mar- ginally significant in the tetracycline line comparison. T o increase the degrees of freedom in the denomi- nator, the replicate line term was pooled with the error term on the basis of the P
>
0.25 criterion (SOKAL and ROHLF 198 1). T h e female incompatibility term was highly significant in both experiments when tested over the pooled error. There were no other significant effects in the ANOVAs apart from a block effect in the tetracycline line comparison. R females therefore had lower fecundity than W females regard- less of which males they mated with. Fecundity of the Rp females was 18% lower on average than that of Wp females, while the fecundity of the RR females was 8% lower on average than WRT females.Mating experiments: Mating success in large cages:
T h e number of matings obtained by R and W males in the large cages are given in Table 9 and were compared with
x2
tests. Mating success did not differ between the incompatibility types except in one of the comparisons between the mass-bred Piru stocks, when Wp males were more successful than Rp males. How- ever, this trend was not apparent in the other tests with these stocks or in the F1 comparisons of the other stocks.Deviations from random mating were examined by calculating G statistics for an association between male and female incompatibility type after organizing the counts into 2 X 2 tables. T h e
G
value was significant ( P<
0.05) for one of the comparisons (Highgrove Fls), where there was a tendency for negative assor- tative mating. However, there was no evidence for assortative mating in the other six trials ( P>
0.10 for the G values), indicating that mating was generally random with respect to incompatibility type.Remating success: Data for the experiments examin-
ing the effect of the first and second male on the tendency of females to remate are given in Table 10. Counts were analyzed by log-linear models, initially fitting a model with the main effects of first male, second male and remating categories. This model is given by
ln($;k) = U
+
ai+
b;+
C k ,TABLE 9
Mating success of males in large population cages indicated by number of males mating
R and W lines compared n w P Sanger (Fls from Rs(1,) X WS) 4 1 5 0 NS Highgrove (Fls from RH X WH(SS)) 7 5 5 6 NS Riverside/Melbourne (Fls from RR X W,) 4 3 4 1 NS Piru (Rp us. WP)
Rep. 1 3 2 7 4 CO.001
Rep. 2 3 3 4 2 NS
Rep. 3 47 41 NS
Rep. 4 47 38 NS
the first and second male interactions were fitted, and these changes were not significant at the 5% level. For the Highgrove stocks, AG was not significant for the first male interaction (AG = 1.04) but was signifi- cant ( P
<
0.05) for the second male (AG = 3.91). Neither the first male (AG = 0.30) nor second male (AG = 0.19) interactions were significant in the exper- iment with the Piru lines.These results suggest that the incompatibility type of the first male had no consistent effects on remating behavior. Females that engage in an incompatible mating are therefore not more likely to remate than females first mated to a compatible male. T h e only significant result was for a second male effect in the experiment with the Highgrove lines, when W males were more successful than R males. However this finding was not repeated in the other two experi- ments.
Sperm competition: Results are presented in Table 1 1. Statistical analyses involved comparisons of treat- ments by nonparametric Kruskal-Wallis tests rather than parametric tests because the presence of zero counts meant that distributions were skewed. As ex- pected, WRT(w) females that were first mated to wild males that were type W (WR,) produced more wild- type offspring than females first mated to RR males (1
+
2 us. 3+
4) in all three experiments. Similarly, WRT(w) females remated with white eye males that were type W (WRT,,)) produced more white eye prog- eny than females remated with RR(w) males (1+
3 us.2
+
4). These comparisons are both significant ( P<
0.001). Incompatibility was therefore present in re- mating females as well as in females mating for the first time.
Remating speed of incompatibility types indicated by the number of females remated
First Male Second Male Remated Not remated Sanger (FIS from Rsc1s) X WS)
W W 15 25
W R 15 25
R W 18 2 3
R R 25 21
W W 29 15
W R 14 28 R W 18 26
R R 17 2 2
W W 2 3 17
W R 29 14
R W 2 2 10
R R 20 2 0
Highgrove (Fls from RH X WH(.W))
Piru (Rp versus Wp)
Interactions between R and W sperm are indicated by comparisons 1 us.
2
and 3 us. 4 for the wild-type progeny and the results for these comparisons were consistent in the three experiments. Neither compar- ison is significant ( P>
0.10), which means that the number of wild-type progeny did not depend on whether the second mating involved a WRT(w) male or an R R ( ~ ) male. Sperm interactions are also indicated by comparisons 1 us. 3 and 2 us. 4 for the white progeny. These comparisons are non-significant ( P>
0. lo), indicating that progeny numbers from the sec- ond male’s sperm did not depend on whether the
female had mated initially to an RR male or a WRT male.
DISCUSSION
We will first summarize our experimental findings and compare them to previous results, then use some simple models to try to interpret the spatial and tem- poral frequency patterns of incompatibility types found in our California population samples.
Fitness experiments: Our laboratory tests provided no consistent evidence for egg-to-adult fitness com- ponent differences, nonrandom mating, differential male mating success or differential sperm competition between the two incompatibility types. In general, the TABLE 11
Number of wild-type (+) and white eye (w) progeny from double matings between different incompatibility types
First Second
+
Progeny w Progenymating mating N Mean SD Mean SD
1 . WK.,. R R ( W ) 19 8 . 3 7 6.97 16.89 10.17
2 . WR.,- WRT(x) 18 10.28 7 4 . 8 9 6 . 7 9 25.61
3. RR WmT(x) 15 2.40 3.76 13.53 11.04
944 A. A. Hoffmann, M. Turelli and L. G. Harshman
tests had sufficient power to detect differences on the order of 10% with probability 0.5. We did, however, find statistically significant reductions of 8% and 18% in the relative fecundities of R females in comparison to W females. T h e smaller value was obtained when comparing tetracycline-treated and untreated lines. NICRO and PROUT (1 990) compared the net “produc- tivity” (including both viability and fecundity effects) of R and W strains from Italy and found 6-1 1 % lower productivity for their R strain. Similar comparisons between treated and untreated lines in Table 10 of HOFFMANN and TURELLI (1988) implied significant fitness reductions of 10-3 1 %, with an average reduc- tion of 22%. These results suggest that R females may generally have significantly lower fecundity than W females, at least under laboratory conditions, and that fecundity may be the fitness component most affected by the incompatibility system. This is fortunate be- cause only fecundity differences can be readily esti- mated with flies from nature.
Our data from field-collected flies suggest that R females suffer a smaller fecundity disadvantage in nature than seen in the laboratory. Although the R females were less fecund in all four experiments, only the 17% difference observed in the Lake Cachuma sample is statistically significant. However, in the larg- est of our three Piru collections, a fecundity difference of 12% would have been been detected with proba- bility 0.8. Our data are consistent with the hypothesis that R females generally do suffer a fecundity disad- vantage, but they suggest that the disadvantage i n nature is likely to be closer to 10% than 20%. We cannot be certain that the deleterious effects are pro- duced by the incompatibility-causing infection. In the comparison of the Piru lines, incompatibility types may be associated with different mtDNA variants or other cytoplasmic Factors that affect fecundity. Tet- racycline treatment will probably not influence the mitochondrial genome, but it may influence other cytoplasmic factors that influence fitness. We found that tetracycline treatment did not affect the produc- tivity of WM lines (unpublished results), but this may not hold for other stocks.
Levels of incompatibility in the f i e l d We have shown that the incompatibility between R males and W females seen in laboratory crosses is also occurring in a polymorphic natural population. In all three analyses of field-collected females from Piru, the egg- hatch for W females was significantly less than for R females. No significant difference was found in our single Lake Cachuma sample. We believe that our data from Piru are more reliable, because the collec- tion site is a rural citrus grove in a predominantly agricultural area that harbors a dense
D.
simulans population. I n contrast, our Lake Cachuma collection site is a heavily used campground with a low densityof flies, many of which may be recent immigrants. Hence, the critical random mating assumption of Equation 1 may not apply to our Lake Cachuma sample. Even at Piru, the level of incompatibility inferred from the field-collected females is much lower than expected from our laboratory experi- ments. Letting H denote the relative number of off- spring produced by incompatible versus compatible crosses, our Piru estimates range from 0.52 to 0.81. Weighting these estimates by the inferred number of incompatible matings that contributed to them pro- duces a pooled estimate of H = 0.56 for our Piru samples. In contrast, for our laboratory experiments with R males of different ages (Figure 3), the relative egg hatch from incompatible crosses does not reach such high values unless the males are more than 2 weeks old.
Several hypotheses can explain this discrepancy. First, most matings in nature may involve fairly old males. This hypothesis is difficult to test, because of the difficulty of determining the age of wild-caught flies; but there is little evidence to support it and much indirect evidence against it. ROSEWELL and SHOR- ROCKS (1 987) estimated that cosmopolitan species of Drosophila, including D. simulans, live only a few days in woodland habitats. However, survival rates vary with habitat. For example, D. mercatorum adults from dry field sites are only a few days old, while adults from humid sites are mostly older than two weeks (JOHNSTON and TEMPLETON 1982). Nevertheless, lab- oratory studies of D. melanogaster (reviewed by PAR- TRIDGE 1988) suggest that male reproductive success reaches a maximum within the first week of adult life and then steadily declines. In addition, we found in our male-age experiments that hatch rates in control crosses decreased significantly with male age. The hatch rates observed for wild-collected R females (0.92-0.98) are comparable to the highest ones ob- served in our laboratory controls (0.89-0.91), sug- gesting that most matings in nature involve young males.
Second, females may mate assortatively i n nature and/or W males may have a systematic mating advan- tage, although we find no repeatable evidence for either phenomenon in the laboratory. Our average estimate of H from the Piru field-collected females is 0.56. To reconcile this with an H value of 0.42, corresponding to laboratory crosses with 1 2-day-old males, we require that W females in nature be more than 1.5 times as likely to mate with a W male as an K male. Assortative mating of this magnitude was unlikely to be detected in our cage experiments i n which roughly 100 mating pairs were examined. How- ever, reconciling the field estimate with H = 0 . 3 ,
roughly 2.25 times as likely to mate with a W male as an R male. Our cage experiments, which examined both relative mating success of R vs. W males and departures from random mating, would have detected such large departures from equal mating success and random mating with probability greater than 0.5.
Thus, it seems unlikely that these factors are sufficient to account for the differences in our laboratory and field estimates of H . Testing for differential mating success in nature will require an assay for the incom- patibility type of wild-collected males.
Third, R males in nature may generally harbor a lower level of infection than males in the laboratory and may therefore be more compatible with unin- fected females. This hypothesis is indirectly supported by our finding that R females from nature occasionally produce W offspring, but R females in the lab do not. It could be directly tested with a quantitative assay for the level of infection.
Fourth, high temperatures in the field may suppress the incompatibility phenomenon. T h e temperatures at Piru routinely exceed 28 O during afternoons in the
late spring, summer and early fall. HOFFMANN, TUR-
ELLI and SIMMONS (1986) found that the incompati-
bility of R males and W females could be suppressed
by rearing larvae at 28". We have not investigated the effects of daily temperature cycles with maxima above this critical value, but it seems plausible that such cycles could contribute to decreased incompati- bility, possibly by reducing the level of infection. This temperature hypothesis can be tested by exposing a reference infected stock to field temperature condi- tions and checking levels of incompatibilty.
A fifth alternative is that W females in the polymor- phic populations surveyed have evolved some resist- ance to the incompatibility system. Alleles that in- crease the compatibility of W females with R males would clearly be favored in a population that has remained polymorphic for many generations. Al- though HOFFMANN and TURELLI (1988) found no evidence for genetic variation among W females in their incompatibility with R males, intense natural selection for many generations will be much more effective in uncovering such variation than our labo- ratory tests. This evolutionary-response hypothesis can be tested by comparing the incompatibility levels shown by W females from locations with and without R males.
W offspring from
R
mothers: As noted above, our laboratory and field data give strikingly different re- sults. In our laboratory experiments, R females have invariably produced R offspring, irrespective of their age or exposure to extreme temperatures. Despite extensive laboratory tests, we have previously re- covered both R and W progeny from only a single female who had inherited the infection paternally(HOFFMANN and TURELLI 1988). We postulated that this female had received only a low level of the infec- tion, which she did not transmit to all of her offspring. In contrast to this very rare event, we obtained both R and W sublines from 4 of 44 isofemale lines that were classified as type R when initially tested in the first laboratory generation from wild-caught females. Both R and W sublines were also obtained from another newly established isofemale line in the same collection that could not be assigned an incompatibil-
ity type. Moreover, two sublines initially characterized as W were characterized as R after additional gener- ations of laboratory culture. These observations sup- port the hypothesis that compatibility with our stand- ard R males is a quantitative phenomenon rather than a qualitative one. A particular density of the micro- organism may be necessary for complete compatibility and reliable maternal transmission. T h e data in BIN- NINGTON and HOFFMANN (1 989) suggest quantitative differences in infection levels between different R
lines and between males of different ages. T h e in- creased compatibility in crosses with old R males is consistent with the lower infection level observed in these males. For some reason, after several genera- tions of laboratory culture, R lines only produce R sublines. This may reflect the effects of selection in these cultures against females that do not carry a sufficient infection level to be compatible with their infected male sibs. Alternatively, conditions in nature may tend to suppress the infection. For instance, flies in some habitats may encounter antibiotics that par- tially cure the infection (STEVENS 1989). This is plau- sible because antibiotic-producing fungi are common on Drosophila resources.
Our Piru data provide a crude estimate of the fraction of W offspring from R mothers in nature. Ignoring the sublines from isofemale line 34 that could not be initially classified, three of the 430 sub- lines tested from R females were repeatably type W. Thus, we might expect at least 0.7% W offspring from R mothers. If there are many females in nature, like
our line 34, that produce significant fractions of both R and W offspring and show an intermediate level of compatibility with R males, the estimate 0.7% may be far too low.