Interactions Between Positive
and Negative Regulators
of
GCN4 Controlling
Gene Expression and Entry Into the Yeast Cell Cycle
Satoshi Harashima,’ Ernest
M.
Hannig and Alan G. Hinnebusch2
Laboratory of Molecular Genetics, National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, Maryland 20892
Manuscript received April 30, 1987 Accepted July 24, 1987
ABSTRACT
The GCN4 gene encodes a transcriptional activator in yeast whose expression is regulated at the translational level in response to amino acid availability. gcn3 mutations block derepression of GCN4
expression in starvation conditions. gcdl andgcd 12 mutations restore derepression of GCN4 expression in gcn3 deletion mutants, suggesting that GCN3 positively regulates GCN4 indirectly by antagonism of these GCD functions. gcdl and g c d l 2 mutations also lead to temperature-sensitive arrest in the G, phase of the cell cycle in gcn3 deletion mutants. The GCN3 allele completely suppresses both derepression of GCN4 expression and the temperature-sensitive growth conferred by gcd 12 mutations and partially suppresses these phenotypes in gcdl mutants. This suggests that the GCN3 product can promote or provide GCD function in nonstarvation conditions even though it opposes GCD function when cells are starved for amino acids. The gcn3-102 allele is completely defective for positive regulation of GCN4 expression; however, it mimics GCN3 in suppressing gcdl and g c d l 2 mutations and thus retains the ability to restore GCD function in nonstarvation conditions. These data suggest that GCN3, GCDl and G C D l 2 have closely related functions required for regulation of GCN4
expression and entry into the cell cycle. We suggest that GCN3 antagonizes the regulatory functions of GCDl and G C D l 2 in starvation conditions either by competing with these factors for the same sites of action or by modifying their structures by physical interaction.
ACCHAROMYCES cerevisiae responds to starvation
S
for any single amino acid by derepressing a large number of enzymes in several amino acid biosynthetic pathways. This cross-pathway regulatory response, known as general amino acid control, operates at the level of transcription of the structural genes that en- code the coregulated enzymes. Trans-acting factors encoded by G C N l - G C N 9 are required for derepres- sion of enzyme expression in starvation conditions, whereas the products of G C D l - G C D l 3 are needed for enzyme repression in nonstarvation conditions. Most gcd mutations are pleiotropic and lead to either tem- perature-sensitive or unconditional slow growth. Some gcdl alleles are also defective for the killer response, failing to maintain M dsRNA at the permis- sive temperature (reviewed in HINNEBUSCH 1986; NIEDERBERGER, AEBI and HUETTER 1986; GREEN- BERG et al. 1986; HARASHIMA and HINNEBUSCH 1986).Genetic analysis has led to a model in which G C N l
is the proximal positive effector in the general control system and is subject to negative regulation by multi- ple GCD genes in nonstarvation conditions. T h e GCD
factors are in turn negatively regulated by G C N l , GCN2 and GCN3 in response to amino acid starvation
’
Laboratory of Microbial Genetics, Department of Fermentation Tech- nology, Osaka University, Yamada-oka 2-1, Suita-shi, Osaka, 565, Japan.To whom correspondence should be addressed.
Genetics 117: 409-419 (November, 1987)
(WOLFNER et al. 1975; HINNEBUSCH and FINK 1983; NIEDERBERGER, AEBI and HUETTER 1986; GREEN-
BERG et al. 1986; HARASHIMA and HINNEBUSCH
1986).3 In support of this model, it was shown that
GCN4 protein binds specifically to cis-acting DNA sequences required for the derepression of structural genes subject to the general control (HOPE and STRUHL 1985; ARNDT and FINK 1986). In addition, several GCN and GCD genes have been shown to control the expression of a GCN4-lacZ fusion in the manner predicted by the genetic model (HINNEBUSCH
1985; HARASHIMA and HINNEBUSCH 1986). Four AUG codons are present in the 5’ leader of GCN4
mRNA and act in cis to repress translation of GCN4
protein in nonstarvation conditions. G C D l and
GCDIO-GCD13 are required for the inhibitory effects of the upstream AUG codons on GCN4 expression, whereas GCN2 and GCN3 are needed to suppress the effects of these sequences in starvation conditions (THIREOS, PENN and GREER 1984; HINNEBUSCH 1984; HINNEBUSCH 1985; MUELLER and HINNEBUSCH 1986; MUELLER, HARASHIMA and HINNEBUSCH 1987). In contrast to this translational mechanism, it has been
4 1 0 S. Harashima, E. M. Hannig and A. G. Hinnebusch
suggested that GCNG and GCN7 positively regulate GCNI at the transcriptional level (GREENBERG et al. 1986).
Some of the known GCN-encoded factors may be involved in recognition of the starvation signal (un- charged tRNA) that triggers the derepression of GCN4 expression (MESSENGUY and DELFORGE 1976). Others may be activated by this signal to antagonize the negative effectors encoded by GCD genes. We are using a genetic approach to identify GCN-encoded factors that might interact directly with GCD gene products. In a previous study, we observed allele- specific effects among gcdl and gcn3 mutations. T h e gcd 1-101 mutation leads to temperature-sensitive (Tsm-) growth at 3 6 ” . T h e gcn3-102 mutation was isolated as a partial suppressor of the Tsm- phenotype of the gcdl-101 mutation. By contrast, a gcn3-IO1 gcd 1-101 double mutant is inviable. T h e observation of markedly different phenotypes for these two dou- ble mutants was taken as an indication that the GCN3 and GCDl proteins might physically interact (HINNE-
BUSCH and FINK 1983). In this communication, we report additional instances of allele-specific effects involving gcdl and gcn3 mutations, and similar, more dramatic interactions occurring between gcn3 and gcdl2 mutations. Our findings suggest that GCN3, GCDl and GCD12 have closely related functions in the regulatory pathway that couples GCNI expression to amino acid availability. In addition, it seems likely that the regulatory and the essential functions carried out by GCDl and GCD12 are closely associated. We speculate that GCN3 competes with o r modifies the products of GCDl and GCD12 in starvation conditions in a way that alters the translational complex to permit more efficient translation of GCN4 mRNA.
MATERIALS AND METHODS
Yeast strain construction: T h e genotypes of all yeast strains employed are given in Table 1. Strain H96 (MATa GCD gcn2-101 gcn3-101 hisl-29 ura3-52 [HISI::lucZ, ura3- 52, integrated at uru3-52) (HARASHIMA and HINNEBUSCH 1986) is sensitive to the amino acid analogue 3-aminotriazole (AT“) as the result of its gcn2 and gcn3 mutations. T h e gcd strains in Table 2 were isolated from H96 as 3-aminotria- zole-resistant (ATr) revertants. T h e gcd mutations thus ob- tained also lead to temperature-sensitivity for growth (Tsm-) (HARASHIMA and HINNEBUSCH 1986). Strains shown in Ta- ble 3 are either gcd revertants isolated from H96 o r AT’ Tsm- meiotic segregants obtained from a back-cross be- tween a gcd gcn2-101 gcn3-101 revertant of H96 a n d strain H I 17, which has the same genotype as H96 except at MAT. Construction of the gcdl-501 strains in Table 4 began with H 4 2 4 (MATagcdl-501 gcn2-101 gcn3-101 hisl-29 inol ura3- 52 [HIS4::LacZ, URA3 integrated at ura3-52]), an AT’ Tsm- meiotic segregant from a back-cross of revertant H56 (HAR-
ASHIMA and HINNEBUSCH 1986) with H 1 17. H460 was de- rived from H424 by “inositolless death” to eliminate the URA3-containing plasmid carrying the HIS4::LacZ fusion in- tegrated at uru3-52 (HARASHIMA and HINNEBUSCH 1986). H 4 2 4 was crossed with strain H3 ( M A T a ura3-52 Leu2-3
Leu2-112) and a Ura- Tsm+’- meiotic segregant (H469) was selected with the presumptive genotype MATa uru3-52 inol gcdl-501. T h e GCN genotype of H469 was confirmed by subsequent crosses to H602 (MATagcn2-101 lys2) and H603 (MATa gcn3-101 lys2). 4 AT’ : 0 AT”, 3 AT’ : 1 AT” and 2 AT’ : 2 AT“ segregation was observed in both crosses, as expected for heterozygosity at GCDl and GCN2 o r GCN3 (GCD GCN strains a r e AT’). Strain H 6 2 9 is a Tsm+’- meiotic segregant from a 4 AT‘ : 0 AT” tetrad produced by the hybrid H469 X H602; H628 was obtained as a Tsm- AT‘ meiotic segregant from a 4 AT’ : 0 A T s tetrad produced by H 4 6 9 X H603. H562 is a Tsm+’- AT’ segregant isolated from a cross between H469 and H37 (MATa gcn4-101 met- 14 ura3-52).
T h e gcd12-503 strains in Table 4 were derived from H427, a MATa gcd1.2-503 gcn2-101 gcn3-101 hisl-29 inol ura3-52 [HZS4::lacZ, URA3, integrated at ura3-521 meiotic segregant from a cross between revertant H 6 4 (HARASHIMA and HINNEBUSCH 1986) and H I 17. H463 was obtained by inositolless death from H427. H626, H625 and H454 a r e meiotic segregants obtained from a hybrid between H427 and H 4 (MATa ura3-52 leu2-3 Leu2-112). T h e genotypes of these strains were confirmed by crosses to GCD gcn2-101, GCD gcn3-101 and GCD GCN tester strains: T h e presence of gcd12-503 in the GCN3 strains H626 and H 4 5 4 was verified by the reappearance of Tsm- meiotic segregants from crosses t o GCN2 gcn3-101 testers. T h e presence of gcn2-101 in H626 was shown by the 0 AT‘ : 4 A T s segre- gation in all asci produced by a hybrid formed with a gcn2- 101 tester. T h e presence of gcn3-101 in H625 was verified by the 2 AT’ Tsm- : 2 ATs Tsm+ segregation observed in all asci produced by a hybrid formed with agcn3-101 tester. Construction of strains H652 and H653 in Table 5 began with strain H647 (MATa gcd12-503 leu2-3 leu2-112), ob- tained from the same cross described above for obtaining H454. H647 was crossed with H650 (MATa gcn3::LEUZ ura3-52 leu2-3 leu2-112) and H651 (MATa gcn3::LEUZ gcd12-503 ura3-52 leu2-3 leu2-112) was isolated as a Leu+ AT‘ Tsm- meiotic segregant from the resulting hybrid. H651 was crossed t o H I 7 (MATa gcn3-102 ura3-52 leu2-3 leu2-112) and H652 was isolated as a Leuf AT’ Tsm- meiotic segregant; H653 was isolated as a Leu- AT“ Tsm+ segregant from the same cross. H824 was also a segregant from cross H647 X H 6 5 0 described above.
Construction of H654 and H660 in Table 5 began with a cross between H628 and H649 (MATa gcn3::URA3 ura3- 52 leu2-3 leu2-112) from which H657 (MATa gcn3::URA3 gcdl-501 u r d - 5 2 ) was isolated as a Ura+ AT‘ Tsm- segre- gant. H657 was crossed with H229 (MATa gcn3-102 ura3- 52 i n o l ) from which H 6 5 4 was obtained as a Ura- Tsm+’- segregant. H825 (MATa gcdl-501 gcn3-101 ura3-52 leu2-3 leu2-112 Lys2) was isolated as a Ura- AT’ Tsm- segregant from the cross between H628 and H649. H825 was crossed with H650 t o obtain H660 as a Leu+ AT‘ Tsm- segregant. EY 152 and EY 153 a r e meiotic segregants from a cross between H654 and H562. EY152 is Tsm+’- and failed to complement a gcn3 tester for its AT” phenotype; EY153 is Tsm- and complements a gcn3 tester for A T sensitivity.
TABLE 1 S. cerarisiae strains
41 1
Strain Genotvw
H4 H17 H54 H56 H57 H58 H59
H6 1
H62 H63 H64 H70 H75 H79 H96 H211 H454
M A T a ura3-52 leu2-3 leu2-112
M A T a gcn3-102 ura3-52 leu2-3 leu2-112 MATagcdlO-501 gcn2-101 gcn3-101 hisl-
2 9 ura3-52 [HIS4::lacZ, ura3-52,
integrated at ura3-521
M A T a gcdl-501 gcn2-101 gcn3-101 hisl-29 ura3-52 [HIS4::lacZ, ura3-52, integrated at ura3-521
M A T O G C D l l - 5 0 1 gcn2-101 gcn3-101 hisl- 29 ura3-52 [HIS4::lacZ, ura3-52,
integrated at ura3-521
29 ura3-52 [HIS4::lacZ, ura3-52,
integrated at ura3-521
29 ura3-52 [HIS4::lacZ, ura3-52,
integrated at ura3-521
2 9 ura3-52 [HIS4::lacZ, ura3-52,
integrated at ura3-521
29 ura3-52 [HISI::lacZ, ura3-52,
integrated at ura3-521
29 ura3-52 [HIS4::lacZ, ura3-52,
integrated at ura3-521
2 9 ura3-52 [HIS4::lacZ, ura3-52,
integrated at ura3-521
M A T a g c d l - 5 0 5 gcn2-101 gcn3-101 hisl-29 ura3-52 [HISI::lacZ, ura3-52, integrated at ura3-521
29 ura3-52 [HISI::lacZ, ura3-52,
integrated at ura3-521
29 ura3-52 [HISI::lacZ, ura3-52,
integrated at ura3-521
M A T a GCD gcn2-101 gcn3-101 hisl-29 ura3-52 [HlS4::EacZ, ura3-52, integrated at ura3-52 ]
MATagcd12-501 gcn2-101 gcn3-101 hisl-
M A T a G C D l l - 5 0 2 gcn2-101 gcn3-101 hisl-
M A T a G C D l l - 5 0 3 gcn2-101 gcn3-101 hisl-
M A T a gcd10-503 gcn2-101 gcn3-101 hisl-
MATagcd12-502 gcn2-101 gcn3-101 hisl-
M A T a gcd12-503 gcn2-101 gcn3-101 hisl-
M A T a gcdl3-501 gcn2-101 gcn3-101 hisl-
M A T a g c d l l - 5 0 5 gcn2-101 gcn3-101 hisl-
M A T a gcdl-101 gcn3-102 ura3-52 M A T a gcd12-503 ura3-52
Strain GenotvDe
H460 H463 H466 H562 H595 H601 H605 H625 H626 H628 H629 H651 H652 H653 H654 H658 H659 H660 H748 H821 H824 F69 F98 EY 125 EY152 EY 153
M A T a gcdl-501 gcn2-101 gcn3-101 hisl-29
M A T a gcd12-503 gcn2-101 gcn3-101 hisl-
M A T a gcn2-101 gcn3-101 hisl-29 i n o l
M A T a gcdl-501 ura3-52 inol
M A T a gcdl-505 gcn2-101 gcn3-101 hisl-29 ura3-52 inol [HIS4::lacZ, ura3-52 inte- grated at ura3-52 ]
MATagcn2-101 lysl
M A T a gcd11-505 gcn2-101 gcn3-101 hisl- inol ura3-52
29 inol ura3-52
ura3-52
2 9 ura3-52 inol [HIS4::lacZ, ura3-52 in- tegrated at ura3-52 ]
M A T a gcd12-503 gcn3-101 ura3-52 M A T a gcd12-503 gcn2-101 ura3-52 leu2-3
M A T a gcdl-501 gcn3-101 ura3-52 lys2 M A T a gcdl-501 ura3-52 lys2
M A T a gcdl2-503 gcn3::LEUZ ura3-52 leu2-
M A T a gcd12-503 gcn3::LEUZ ura3-52
M A T a gcdl2-503 gcn3-102 ura3-52 leu2-3
M A T a gcdl-501 gcn3-102 ura3-52 gcdl-504 gcn2-101 gcn3-101 h i s 1 2 9 ura3-
52 [HISI::lacZ, ura3-52 integrated at
ura3-521
gcdl-506 gcn2-101 gcn3-101 hisl-29 ura3-
52 [HIS4::lacZ, ura3-52 integrated at
ura3-521
M A T a gcdl-501 gcn3::LEUZ ura3-52 lys2 leu2-3 leu2-112
M A T a gcn3-IO1 ura3-52 M A T a gcn3-101 lys2
M A T a gcd12-503 gcn3::LEUZ ura3-52 leu2-3 leu2-112
M A T a gcn3-102 inol ura3-52 [HISI::lacZ, U R A 3 integrated at ura3-521
M A T a gcdl-101 ura3-52
M A T a gcn3::LEUZ ura3-52 leu2-3 leu2-112 M A T a gcdl-501 gcn3-102 inol ura3-52 M A T a gcdl-501 ura3-52
leu2-112
3 leu.?-112
leu2-3 leu2-112
leu2-112
sequence (E. M . HANNIC a n d A. G. HINNEBUSCH, unpub- lished data). gcn3 deletion plasmids were constructed by replacing a 782-bp HindIII-ClaI fragment which contains 60% of the GCN3 coding sequences a n d 0.22 kb of 5’
flanking DNA with the 2.18-kb LEU2 XhoI-Sal1 fragment (Ep146) or with the 1.1-kb URA3 Hind111 fragment (Ep149) (E. M . HANNIG and A. G. HINNEBUSCH, unpublished data). Plasmid C102-9 is a low-copy plasmid containing the GCN2 gene (DRISCOLL PENN, THIREOS a n d GREER 1984) on a 7.1- kb fragment, kindly provided by MONICA DRISCOLL PE”.
Genetic methods: Yeast genetic techniques and culture media (SHERMAN, FINK and LAWRENCE 1974) as well as procedures for determination of 3-aminotriazole resistance a n d temperature sensitivity (HARASHIMA a n d HINNEBUSCH
1986) were described previously. 3-aminotriazole was used at 30 mM. Yeast transformation was d o n e according to the
method of Ito et aE. (1983). Gene replacements a t GCN3 were conducted by the method of ROTHSTEIN (1 983) using the 4-kb NruI-BglI fragment of Ep146 o r the 4-kb EcoRI- PvuI fragment of Ep149 for yeast transformation. DNA blot-hybridization analysis of total transformant DNA was conducted to confirm all gene replacements (E. M. HANNIC a n d A. G. HINNEBUSCH, unpublished data). T h e gcn3::URA3 andgcn3::LEU2 deletion alleles conferred an ATs phenotype in GCD strains a n d this phenotype segregated 2 A T s Ura+ (or Leu+) : 2 AT‘ Ura- (or Leu-) in meiotic analysis of hybrids formed between the deletion strains and GCN ura3-
52 leu2-3 leu2-112 strains.
412 S. Harashima, E. M. Hannig and A. G. Hinnebusch
Cell-cycle studies: For each strain analyzed, one-half o f an asynchronous culture growing exponentially in liquid YPD medium at 23' was shifted to 36' while the other half was maintained at 2 3 " . Six hours after the shift, an aliquot was taken from each culture, sonicated for 20 sec and the percentage o f unbudded cells was measured by light micro- scope examination of 300 cells.
RESULTS
g c d l and g c d l 2 mutations are suppressed by
GCN3: gcn2 and gcn? mutants are unable to derepress amino acid biosynthetic enzymes in starvation condi- tions. As a result, they are sensitive to 3-aminotriazole, an inhibitor of the HIS3 gene product. We previously described the isolation of gcd mutants by reversion of the 3-aminotrazole-sensitive (AT') phenotype of a
gcn2-I01 gcn?-101 double mutation. T h e gcd muta- tions we obtained also confer temperature-sensitive or unconditional slow growth (HARASHIMA and HINNE- BUSCH 1986). In the course of our genetic analysis, we crossed the gcd gcn2 gcn3 revertants by GCD GCN
strains and analyzed the meiotic products of the re- sulting hybrids. T h e simple expectation was to find in each ascus two Tsm-, 3-aminotriazole-resistant (AT') spores containing the gcd mutation involved in the cross. In most asci, the remaining two spores were expected to have a Tsm+ ATs phenotype since they lack a gcd mutation but contain either gcn2-I01 o r
gcn3-101. This proved to be the case in asci produced by hybrids containing g c d l 0 , g c d l l and gcdl? muta- tions; however, the crosses involving gcdl and g c d l 2
mutations gave rise to a significant number of asci in which either three or all four spores had a Tsm+ AT" phenotype (data not shown). This suggested that expression of the phenotypes of gcdl and g c d l 2 mu- tations in our revertants might be dependent upon the presence of gcn2-I01 o r gcn?-l01. We examined this possibility in two ways.
T h e first approach was to cross the AT' Tsm- gcd gcn2-I01 gcn3-I01 revertants by GCD gcn2-I01 GCN?
and GCD GCN2 gcn?-l01 strains, thus making either
gcn2-I01 o r gcn?-I01 homozygous in each cross. In the case of gcdl and g c d l 2 revertants, we observed 2
AT' Tsm-: 2 ATs Tsm+ segregation in nearly every ascus produced by hybrids in which gcn?-I01 is homo- zygous. By contrast, hybrids which are homozygous for gcn2-I01 and heterozygous at GCN3 also gave rise to a significant number of 1 AT' Tsm-: 3 ATs Tsm+ and 0 AT' Tsm-: 4 ATs Tsm+ asci (Table 2). All hybrids constructed from revertants containing mu- tations in GCDIO, G C D l l or GCDl? gave rise to only 2 A T ' Tsm-: 2 AT" Tsm+ asci. These results suggest that the gcdl and g c d l 2 mutations present in the revertants are suppressed by the GCN? allele. In the case of the g c d l 2 mutations, suppression appeared to be complete; for gcdl mutations, suppression of both mutant phenotypes appeared to be incomplete (Table
2)-
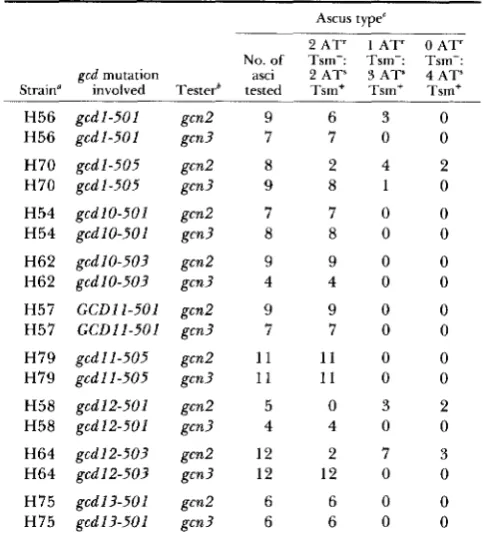
TABLE 2
Tetrad segregation from hybrids between gcd gcn2-101 gcn3-I01 revertants and GCD gcn2-101 GCN3 and GCD G C N 2 gcn3-101
tester strains
Ascus type'
2 A T ' 1 A T ' OAT'
No. of Tsm-: Tsm-: Tsm-: gcd mutation asci 2 A T ' 3AT" 4 A T ' Strain" involved Testerb tested Tsm+ Tsm+ Tsm+
H56 g c d l - 5 0 1 gcn2 9 6 3 0
H56 g c d l - 5 0 1 gcn? 7 7 0 0
H70 g c d l - 5 0 5 gcn2 8 2 4 2 H70 g c d l - 5 0 5 gcn3 9 8 1 0
H54 gcd10-501 gcn2 7 7 0 0
H54 gcd10-501 gcn3 8 8 0 0
H62 g c d I 0 - 5 0 3 gcn2 9 9 0 0
H62 gcd10-503 gcn? 4 4 0 0
H57 G C D I I - 5 0 1 gcn2 9 9 0 0
H57 G C D I I - 5 0 1 gcn? 7 7 0 0
H79 gcd11-505 gcn2 1 1 1 1 0 0
H79 g c d l l - 5 0 5 gcn? 1 1 1 1 0 0
H58 g c d l 2 - 5 0 1 gcn2 5 0 3 2
H58 gcd12-501 gcn3 4 4 0 0
H64 gcd12-503 gcn2 12 2 7 3
H64 gcd12-503 gcn3 12 12 0 0
H75 gcd13-501 gcn2 6 6 0 0
0
6 0
H75 g c d l 3 - 5 0 1 gcn3 6
All gcd strains have the genotype gcd gcn2-101 gcn3-101 hisl- 29 ura?-52 [HIS4::lacZ, URA? integrated at ura3-521.
The gcn2 and gcn3 tester strains are H601 (MATa GCD gcn2- I01 GCN3 l y s l ) and H821 (MATa GCD GCN2gcn3-101 lys2).
For the gcdl revertants, the actual ascus types observed from left to right were 2 AT' Tsm-: 2 ATs Tsm+, 1 AT' Tsm-: 1 AT'/" Tsm+/-: 2 ATs Tsm+ and 2 AT'" Tsm+/-: 2 AT" Tsm+, where AT"
and Tsm+/- designate weak AT resistance and weak temperature sensitivity, respectively.
Suppression of g c d l and g c d l 2 mutations by the cloned GCN3 gene: In the second approach, we trans- formed gcd gcn2-101 gcn?-I01 strains with single-copy plasmids carrying the cloned GCN2 o r GCN? genes. T h e results in Table 3 indicate that the GCN3 gene completely suppressed the AT' and Tsm- phenotypes of three g c d l 2 mutants and partially suppressed both phenotypes in five gcd I mutants. Transformation of the same strains with the GCN2 gene (Table 3) or with a gcn? frameshift mutation constructed in vitro (data not shown) had no effect on the phenotype of the gcdl
and g c d l 2 mutants. Transformation of two out of five
g c d l l gcn2-I01 gcn3-I01 mutants with the GCN? gene gave the opposite result of exacerbating the slow growth phenotype of these mutants. T h e phenotypes associated with all other mutant alleles of GCDIO, G C D l l and GCDl? were unaffected by introduction of the GCN2 o r GCN3 gene.
Interactions Among GCN4 Regulators 413
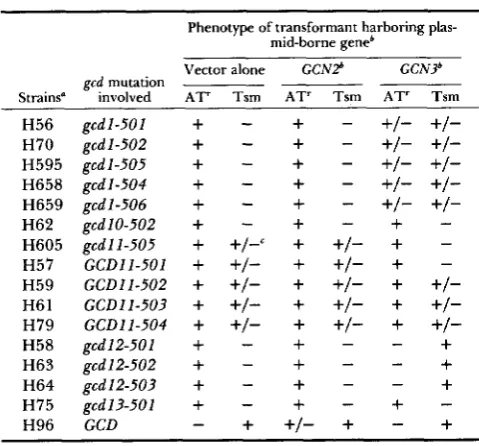
TABLE 4 TABLE 3
Suppression of gcdl andgcdl2 mutations by the cloned GCN3 gene in gcd gm2 gcn3 strains
Constitutive derepression of GCN4lacZ expression in gedl2- 503 and gedl-501 mutants is suppressed by GCN3
~~ ~
Phenotype of transformant harboring plas- mid-borne geneb Strains" H56 H70 H595 H658 H659 H62 H605 H57 H59 H6 1 H79 H58 H63 H64 H75 H96
gcd mutation involved
gcdl-501 gcd 1-502 gcdl-505 gcd 1-504 g ~ d 1-506 gcd 10-502 gcd 11 -505 GCDl l-501 G C D l l - 5 0 2 GCDI 1-503 G C D l l - 5 0 4 gcd 12-50 1 gcd 12-502 gcd 12-503 gcd13-501 GCD ~ Vector alone
AT' Tsm
+
-+
-+
-+
-+
-+
-+
+/-c+
+/-+
+/-+
+/-+
+/-+
-+
-+
-+
-+
-
G C N P
AT' Tsm
+
-+
-+
-+
-+
-+
-+
+/-+
+/-+
+/-+
+/-+
+/-+
-+
-+
-+
-+I-
+
GCN?
Strain" Genotvue AT' Tsm AT' Tsm
+/- +/- +/- +/- +/- +/- +/- +/- +/- +/-
+
-+
-+
-+
+/-+
+/-+
+/--
+
-
+
+
+
--
+
-All strains contain gcn2-101, gcn3-101, hisl-29, ura3-52 and a HIS4::lacZ fusion integrated at ura3-52. H595 also contains an inol mutation.
'
GCN2 and GCN3 were contained on stable, low-copy, autono- mously replicating plasmids C102-9 and Ep69, respectively. YCp50 was used as the "Vector alone."' For g c d l l mutants, a Tsm phenotype of +/- indicates uncon- ditional slow growth.
sion in the gcd mutants shown in Table 4. T h e results demonstrate an excellent correlation between the de- gree of A T resistance, temperature-sensitivity and derepression of GCN4::lacZ expression in the various
gcd strains. As expected, the gcn3-I01 mutation blocked efficient derepression of GCN4::lacZ expres- sion in the ATs GCD gcn3-101 strain H748 compared to AT' GCD GCN strain H4. By contrast, gcn3-I01
led to higher levels of GCN4::lacZ expression in the presence of the gcd 1-501 o r gcd 12-503 mutations com- pared to the expression observed in the corresponding
gcd G C N 3 strains: gcd12-503 gcn3-101 strain H625 exhibited constitutive derepression of GCN4::lacZ
expression at a level 40- to 50-fold higher than the repressed level observed in fully wild-type strain H4. By contrast, gcd12-503 G C N 3 strain H454 exhibited
GCN4::lacZ expression very similar to that seen in
GCD GCN strain H4. Therefore, the G C N 3 allele completely suppressed the effect of gcd 12-503 on
GCN4::lacZ expression. In the case of gcdl-501, the
GCN3 allele did not completely eliminate constitutive derepression of GCN4::lacZ expression, but led to 3- to 5-fold lower expression than observed in the gcdl- 5 0 1 gcn3-101 strain. By contrast to the effects of gcn3- 101, the gcn2-101 allele led to lower GCN4::lacZ
expression relative to the G C N 2 allele in the presence or absence of mutations in GCDl o r GCDI2.
Disruption of GCN3 leads to efficient expression
GCN4::lacZ en- zyme activity
(units)b
R DR
H463 H626 H625 H454 H460 H629 H628 H562 H466 H748 H4
g c d l 2 gcn2 gcn3 gcd 1 2 gcn2 GCN3 g c d l 2 GCN2 gcn3 g c d l 2 GCN2 GCN3
gcd 1 gcn2 gcn3 gcd 1 gcn2 GCN3 gcdl GCN2 gcn3 gcd 1 GCN2 G C N 3
GCD gcn2 gcn3 GCD GCN2 gcn3 GCD GCN2 G C N 3
+
--
+
+
-+
+
+
-+
-+
+/-+
+/--
+
+
+
+
-
240 340 21 33 1000 830 35 150
1300 1500 390 290 2300 3200 450 730
27 15 44 69 20 225
a Each strain was transformed with plasmid p180, a stable, low- copy, autonomously replicating plasmid containing a GCN4::lacZ fusion.
* These values are the averages
of 2-3 assays on independent transformants that varied from the average by 30% or less. The repressing (R) conditions were 6 hr of growth to mid-exponential phase in minimal medium supplemented with leucine, isoleucine, valine and inositol as described (HINNEBUSCH 1985); the derepress- ing (DR) conditions were 6 hr of culturing in the same medium containing 5-methyltryptophan at 0.5 mM, following 2 hr of growth in the absence of 5-methyltryptophan.of gcdl and gcdl2 mutations: We replaced the gcn3- 101 mutation in several gcd strains with a gcn3 allele containing a deletion of 60% of the coding region plus an insertion of the yeast URA? o r LEU2 gene. In
GCD haploid strains, the gcn3::URA3 allele confers an ATs Tsm+ phenotype similar to that conferred by
gcn3-101. This suggests that G C N 3 is not essential in
GCD cells grown in non-starvation conditions and is only required for derepression in starvation condi- tions (E.
M.
HANNIG and A. G. HINNEBUSCH, unpub- lished data). When we replaced gcn3-101 withgcn3::URA3 in twogcd I 2 gcn2-I01 gcn3-I01 revertants (H63 and H64) and in the g c d l - 5 0 1 gcn2-I01 gcn3- 101 revertant H56, we found that the transformants exhibited an AT' Tsm- phenotype similar to that of the untransformed parent strains containing gcn3- 101. This suggests that the gcn3-101 mutation is sim- ilar to a null allele of G C N 3 and that elimination of the G C N 3 gene product is the necessary condition for full expression of g c d l and g c d l 2 mutant phenotypes. We observed only one instance of allele specificity in these gene replacement experiments: gcd 12-502 gcn2-I01 gcn3::URA3 transformants exhibit a more severe growth defect at the semipermissive tempera- ture than their isogenic parent strain containing the
gcn3-101 allele (Figure 1). This more extreme growth defect was always found to be associated with
414 S. Harashima, E. M. Hannig and A. G. Hinnebusch
F I G U R E 1 .--Kepklcement of gcn3-1OI with gcn3::tih\3 in ii grd12-502 nilltilnt kads t o iln incre;ised growti drfiw. Strain t{ti:< (Rrd12-502
gm2-101 gm3-101 ura3-52). shown on the left, and a Ura' transformant of H63 containinggcn3::UiRA3 in place ofgcn3-101. shown on the right. were plated for single colonies o n complete medium (YPD) and incubated at 30" for 3 days.
cross we also observed roughly equal numbers of Ura-
and Ura+ segregants which have the AT' Tsm- phe- notype indicative of the g c d l 2 mutation. This shows that g c d l 2 and gcn3 mutations a r e unlinked and rules out the possibility that g c d l 2 mutations a r e constitu- tively derepressing alleles of GCN3.
T h e data shown in Table 5 indicate that gcn3::LEU2
behaves like gcn3-I01 and leads to constitutive de- repression of GCN4::lacZ expression in a gcd12-503
strain. T h e degree of derepression is significantly lower in the gcd 1 2 gcn3::LEU2 strain compared to the
g c d l 2 gcn3-I01 strain. However, GCN4::lacZ expres- sion is clearly both constitutive and depressed in g c d l 2 gcn3::LEU2 cells by comparison with g c d l 2 G C N 3
cells, where expression is very similar to that observed in wild-type cells. GCN4::lacZ expression was also con- stitutive in thegcdl g&3::LEU2 strain shown in Table
5 ; however, in this case, the degree of derepression is comparable to that seen in the gcdl G C N 3 strain.
gcn3-102 behaves like GCN3 and suppresses the
gcdl and gcdl2 mutations: Like other gcn3 mutations we examined, gcn3-102 leads to an AT' phenotype
and blocks derepression of GCN4-lac2 expression in
GCD cells (Table 5). However, gcn3-102 differs from
gcn3-I01 and gcn3::URA3 in suppressing the pheno- types of gcdl and g c d l 2 mutations. GCD gcn3-102
strain F69 was crossed with the gcdl-501 gcn2-101 gcn3-I01 revertant H 5 6 and the g c d l 2 g c n 2 - I 0 1 gcn3-
IO1
revertants H58, H 6 3 and H64. If gcn3-102 actslike gcn3-I01 and leads to expression of the gcdl and
g c d l 2 mutant phenotypes (AT' Tsm-) all asci pro- duced by these hybrids should exhibit 2 AT' Tsm-: 2
AT' Tsm+ segregation. However, in addition to such asci, we observed 1 AT' Tsm-: 3 ATs Tsm+ and 0 A T ' Tsm-: 4 ATs Tsm+ segregation, suggesting that suppression of thegcd mutations had occurred in these asci. Subsequently, we crossed gcd12-503 gcn3::LEU2
strain H 6 5 1 with G C D l 2 gcn3-102 strain H 17. As above, all three types of asci were produced by this hybrid and all Leu- segregants (which contain gcn3- 1 0 2 ) were A T ' Tsm+ whereas all AT' Tsm- segre- gants were Leu+. These results demonstrate directly that the mutant phenotypes associated with gcd12-503
Interactions Among GCN4 Regulators 415
TABLE 5
Allele-specific interactions between gcn3, gcdl and gcdl2 mutations affect expression of C C N 4 l a e Z
GCN4::lac.Z en- zyme activity
(units)'
Strain" Genotvwb AT' Tsm R DR
H625 H652 H454 H653
H628 H660 H562 H654 H748 EY 125 H4 H17
gcd 1 2 gcn3-10 1 gcd 1 2 gcn3::LEUZ g c d l 2 GCN3 g c d l 2 gcn3-102
gcdl gcn3-101 gcdl gcn3::LEUZ gcdl GCN3 gcd lgcn3-102
GCD gcn3-101 GCD gcn3::LEUZ
GCD GCN3
GCD gcn3-102
+
-+
-+
+
+
+
-+
--
+
+/-+
+/-+
-
+
+
+
+
-
-
1000 210 35 15
2300 550 450 230
44 28 20 14
830 190 150 24
3200 600 730 190
69 28 225 26
a Each strain was transformed with plasmid p180, a stable, low- copy, autonomously replicating plasmid containing a GCN4::lacZ fusion.
*
All strains are GCN2.These values are the averages of 2-3 assays on independent transformants that varied from the average by 30% or less. Repress- ing (R) and derepressing (DR) growth conditions were the same as described in Table 4.
nants. As a final proof of this conclusion, we carried out a cross between g c d l 2 - 5 0 3 gcn3::LEU2 strain H824 and gcd12-503 gcn3-102 strain H653 and ob- served
2
AT" Tsm- Leu':2
ATs Tsm+ Leu- segre- gation in 10/10 asci produced by this hybrid.In agreement with these findings, Table 5 shows that GCN4::lacZ expression in a gcd12-503 gcn3-102
strain was 50-fold lower than that seen in a gcd12-503 gcn3-101 strain, and was indistinguishable from that seen in a GCD gcn3-102 strain. In agcn3-102 g c d l - 5 0 1
double mutant, GCN4-lac2 expression was lower than that observed in any other g c d l - 5 0 1 strain we exam- ined, particularly in a gcdl-501 gcn3-101 mutant. We conclude that the gcn3-102 allele closely resembles
G C N 3 in completely suppressing the phenotypes of the gcd 1 2 mutations and partially suppressing the gcd 1
mutations.
Allele specificity in the interaction between gcn3- 102 andgcdl-101: gcn3-102 was isolated as a suppres- sor of the Tsm- phenotype of the gcdl-101 mutation in a G C N 3 strain (HINNEBUSCH and FINK 1983). As expected, a hybrid between gcdl-101 gcn3-102 double mutant H211 and gcdl-101 G C N 3 single mutant F98 gave rise predominantly to asci exhibiting 2 Tsm+/-: 2 Tsm- segregation. We showed above that gcn3-102
also partially suppresses the Tsm- phenotype of a
gcdl-501 mutation. In a cross identical to H211 X F98 just described, except that gcdl-501 replaces g c d l - 1 0 1
(EY 152 X EY 153), all four spores in every tetrad were Tsm+/-; however, two degrees of temperature-sensi- tivity were evident. Two ascospores in each tetrad
exhibited a less severe temperature-sensitivity than their two sister spores. We demonstrated by comple- mentation testing for the ATs phenotype that all of the less temperature-sensitive ascospores from this cross contain the gcn3-102 allele. Therefore, gcdl-501
is also suppressed more efficiently by gcn3-102 than by GCN3. However, the degree of suppression o f g c d l - 5 0 1 by gcn3-102 in cross EY 152 X EY 153 was signif- icantly less than we observed for gcdl-101 in cross H211 X F98. As a result, at least one component of the suppressive interaction between gcn3-102 and
gcdl-101 may be specific to these two alleles.
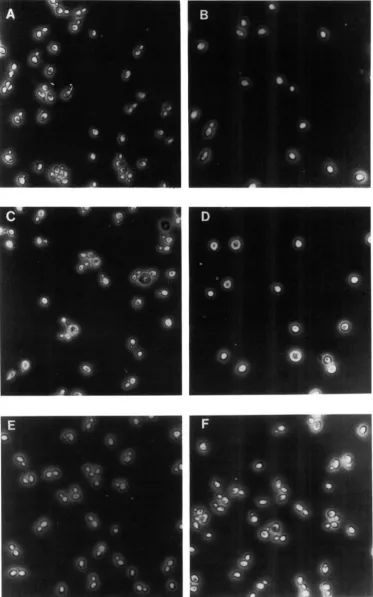
gcdl and gcdl2 mutants arrest in the GI phase of the cell cycle at the restrictive temperature: Because the previously isolated gcdl-101 mutation leads to GI arrest at 36" (WOLFNER et al. 1975), we examined the terminal phenotype of our newly isolated temperature sensitive gcd revertants. Figure
2
shows that g c d l - 5 0 1strain H56 and gcd12-503 strain H64 accumulate as spherical unbudded cells after 6 h r of incubation at 36". By contrast, the Tsm- gcdlO (H62) and g c d l 3
(H75) mutants failed to accumulate with a character- istic morphology when treated in the same way (data not shown).
DISCUSSION
At temperatures permissive for their growth, g c d l
and gcd 1 2 mutants exhibit derepressed expression of genes encoding enzymes subject to general amino acid control. This phenotype is attributable to increased expression of G C N 4 , the direct positive regulator of these genes (HINNEBUSCH 1985; HARASHIMA and HIN- NEBUSCH 1986). T h e G C D l and GCD1.2 products re- press G C N 4 expression at the translational level by controlling the inhibitory effects of AUG codons lo- cated in the 5' leader of G C N 4 mRNA (HINNEBUSCH 1985; MUELLER and HINNEBUSCH 1986; MUELLER, HARA~HIMA and HINNEBUSCH 1987). At the restrictive temperature, g c d l and g c d l 2 mutants arrest in the GI phase of the cell cycle. This temperature-sensitive phenotype remains evident after inactivation of
G C N I , showing that at least one component of the Gcd- growth defect is unrelated to overexpression of GCN4 protein in the gcd mutants (HARASHIMA and HINNEBUSCH 1986). Therefore, it appears that G C D l
and G C D l 2 either execute o r regulate the expression of an essential function, in addition to their role in translational control of GCN4.
T h e non-derepressible phenotype of GCD gcn3:: U R A 3 strains clearly demonstrates that inactivation of
G C N 3 interferes with derepression of G C N 4 in star- vation conditions and verifies that G C N 3 is a positive effector of GCN4. Recessive mutations in either G C D l
or G C D l 2 overcome the derepression defect in
416 S. Harashima, E.
M.
Hannig a n d A. G. HinnebuschFIGURE 2.-gcdl and gcd.12 mutants arrest as unbudded cells at 36". g c d l strain H56 (A and B), g c d l l strain H64 (C and D) and CCD
Interactions Among GCN4 Regulators 417
G C D l 2 repressing functions in starvation conditions (HARASHIMA and HINNEBUSCH 1986). T h e surprising conclusion from the experiments presented here is that the G C N 3 product also appears to promote the negative regulatory function and the essential func- tion carried out by the products of these GCD genes in nonstarvation conditions. Thus, the gcn3-101 and
gcn3::URA3 mutations enhance the derepression of
GCN4::lacZ expression and the temperature sensitivity associated with gcd 1 mutations, and are absolutely required for expression of these phenotypes in g c d l 2
mutants. T h e G C N 3 allele completely suppresses both phenotypes for all three gcd 1 2 alleles and partially suppresses each of five g c d l mutations we examined. These findings suggest that the absence of G C N 3 gene product is the necessary condition for full expression of g c d l and g c d l 2 mutant phenotypes.
Suppression of the gcd mutations by GCN3 could be explained in several ways. One possibility is that G C N 3
can a t least partially complement the loss of G C D l or
G C D l 2 function. In this view, G C N 3 and these two
GCD genes carry out analogous functions, as in the case of the duplicated yeast histone H2B genes and the yeast RAS genes, to name only a few examples (RYKOWSKI et al. 1981; KATAOKA et al. 1984; TATCH- ELL et al. 1984). Such functional complementation would imply that G C N 3 can provide GCD function in nonstarvation conditions even though it acts to oppose
GCD function in starvation conditions. This suggests the interesting possibility that G C N 3 acts as a positive effector of GCN4 by competing with GCDl and
GCD12 in the regulatory mechanism in conditions of amino acid starvation. It then functions inefficiently in these circumstances as a GCD factor in repressing
GCN4 expression (Figure 3A). gcn3-102 is completely defective for G C N 3 positive regulatory function, yet it behaves like GCN3 in restoring a degree of GCD
function to gcdl mutants and in completely suppress-
i n g g c d l 2 mutations. T h e complementation model can account for this finding by proposing that the gcn3- 1 0 2 mutation leaves intact the ability of the G C N 3
product to provide GCD function in repressing con- ditions. T h e gcn3-102 mutation impairs only the abil- ity of the GCN3 protein to respond correctly to the inducer in derepressing conditions after it replaces the GCD gene product in the regulatory mechanism (Figure 3A).
Another possible explanation for our results is that the GCN3 and gcn3-102 gene products suppress the phenotype of gcdl and g c d l 2 mutations by stabilizing thermolabile proteins encoded by these gcd alleles. According to this model, the G C N 3 protein forms a complex with the G C D l and G C D l 2 proteins. gcn3- 101, the allele present in the strains in which the gcdl
and g c d l 2 mutations were isolated, and gcn3::URA3
would encode products incapable of complex forma-
tion with the GCD proteins. As a result, the thermol- ability of the gcd products would be expressed in gcn3- 101 and gcn3::URA3 strains, thus leading to a Gcd- phenotype. By contrast, the products of the G C N 3
and gcn3-102 alleles would stabilize the mutant g c d l
and g c d l 2 proteins, thus suppressing the Gcd- phe- notype (Figure 3B).
T h e idea of complex formation between G C N 3 and
GCD proteins is attractive because it offers a mecha- nism by which G C N 3 could antagonize GCD function in starvation conditions: Modification of the G C N 3
protein in response to starvation could alter the struc- ture and activity of the GCD proteins with which it interacts. In this model, the gcn3-102 product would be capable of complex formation with GCD proteins and could therefore suppress the phenotypes of gcd
mutations; however, it would be unable to antagonize the GCD factors in starvation conditions and incapable of promoting derepression of GCN4 expression.
T h e gcn3-102 mutation suppresses the temperature sensitivity of the g c d l - 1 0 1 mutation more efficiently than the G C N 3 allele (HINNEBUSCH and FINK 1983). In addition, measurements of GCN4-lacZ expression suggest that gcn3-102 leads to greater repression of
GCN4 in gcdl-101 strains than does the G C N 3 allele (HINNEBUSCH 1985). These findings suggest that the
gcn3-102 product is more efficient than the G C N 3
product at restoring GCDl function in gcdl-101 cells. It was suggested previously that the gcn3-102 muta- tion leads to a specific change in the G C N 3 protein that compensates for a structural alteration in the product of g c d l - 1 0 1 with which it interacts (HINNE- BUSCH and FINK 1983). This explanation is in accord with the model we present in Figure 3B. However, we have shown here that gcn3-102 is also more effi- cient than G C N 3 in suppressing a different g c d l mu- tation isolated independently of gcn3-102. While the suppressor activity of gcn3-102 is considerably less for the new gcdl mutation than for gcdl-101, the absence of unequivocal allele specificity in these interactions prevents us from arguing more strongly in favor of a protein-protein interaction between G C N 3 and G C D l .
T h e complementation model in Figure 3A can easily account for the suppressor activity of gcn3-102 by proposing that the product of this allele has greater
GCD function than the wild-type G C N 3 product. A fact that is more difficult to explain by complex formation is that the gcn3::URA3 and gcn3-101 muta- tions have no effect on either the essential or regula- tory functions of the G C D l and G C D l 2 gene products in GCD cells. To account for this observation, it could be proposed that the GCD gene products have associ- ations with other proteins, in addition to the G C N 3
418 S. Harashima, E. M. Hannig and A. G. Hinnebusch
B
Starvation
Non-starvation
A
N
on-sta rvation
Starvation
GCD12 GCN3
GCDl2 GCN3 /\GCN3
GCD 12
rYnducer
6
1.
GCN4mRNA5
I=
.-+
3'gcdl2 GCN3
I
-
gcd 12 GCN3
! I
L
-
gcdl2 gcn3: :URA3I
gcdl2 gcn3: URA3
8
3 '
v
ncd12 gcn3- 102 gcd 12 gcn3- 102
FIGURE 3.-Two models to explain the interactions between GCDlZ and GCN3. (A) Functional complementation of a g c d l 2 mutation by G C N 3 and gcn3-102. Under nonstarvation conditions, the G C N 3 product can substitute for the g c d l 2 product and repress translation of GCN4 mRNA. The positive regulatory function of G C N 3 in wild-type cells results from replacement of the G C D l 2 product by the functionally analogous GCN? product. This replacement depends upon the action of an inducer present only in starvation conditions. Interaction with the inducer also decreases the efficiency of the GCN? product in repressing translation, thus allowing increased expression of GCN4 protein. The gcn3-102 product can efficiently repress translation in a g c d l 2 mutant in non-starvation conditions and it retains this function in starvation conditions because it fails to interact with the inducer. (B) Direct interaction between the products of G C D l 2 and GCN? orgcn?- 102. Suppression of a g c d l 2 mutation by the GCN? product is explained by an interaction between the two proteins that corrects an instability in the g c d l 2 product. Only when G C N 3 protein is missing is the g c d l 2 product instability manifested. In response to the influence of an inducer present in starvation conditions, the G C N 3 product alters the configuration of the G C D l 2 or g c d l 2 products and renders them incapable of efficiently repressing translation of GCN4 mRNA. The gcn3-102 product retains the ability to interact with and stabilize the g c d l 2 product but cannot alter its configuration to allow derepression because it fails to interact with the inducer. In both models, interactions that lead to proper repression of GCN4 expression in nonstarvation conditions also suppress the temperature-sensitive growth defect associated with g c d l 2 mutations.
GCN3 would then result in destabilization of the com- plex and a Gcd- phenotype. T h e complementation model can account for the same observation by pro- posing that GCN3 can provide a degree of GCD func- tion in the absence of GCDl o r GCD12, but its contri- bution is dispensible in GCD cells grown in non-star- vation conditions.
Regardless of which model is correct, our data strongly suggest that GCN3, GCDl and GCD12 have closely related functions in the translational control of GCN4. This makes these gene products excellent candidates for factors that function at the interface between the positive and negative regulators control- ling GCN4 expression. In addition, we have noted a good correlation between the effects of gcn3 muta- tions on the regulatory and the essential functions of
G C D l and GCD12: With only one exception, the
severity of the Tsm- phenotype of a gcd mutant was directly correlated with the degree of constitutive derepression of GCN4: :lacZ expression. T h e effects of
GCN3 upon both phenotypes suggests that the essen- tial and regulatory functions of the GCD proteins are interrelated. Given that GCN4 is regulated by these factors at the translational level (MUELLER et aE. 1987), we speculate that the essential functions carried out by the GCD factors affect protein synthesis. In this view, the role of GCN factors would be to alter or compete with one o r more GCD factors whose func- tions in the translational complex are required to repress translation of GCN4 mRNA. Our genetic ob- servations make GCN3 a likely candidate for a factor that participates directly in such interactions with the products of GCDl and GCD12.
Interactions Among GCN4 Regulators 419
LER for valuable comments on the manuscript, ALICE MA for technical assistance and DOTTIE ALLOR for preparation of the manuscript.
L I T E R A T U R E C I T E D
ARNDT, K., and G. R. FINK, 1986 GCN4 protein, a positive tran- scription factor in yeast, binds general control promoters at all 5' TGACTC 3' sequences. Proc. Natl. Acad. Sci. USA 83:
Temporal analysis of general control of amino acid biosynthesis in Saccha- romyces cereuisiae: role of positive regulatory genes in initiation and maintenance of mRNA derepression. Mol. Cell. Biol. 4:
GREENBERG, M. L., P. L. MYERS, R. C. SKVIRSKY and H. GREER, 1986 New positive and negative regulators for general con- trol of amino acid biosynthesis in Saccharomyces cereuisiae. Mol. Cell. Biol. 6: 1820-1829.
Multiple GCD genes required for repression of GCN4, a transcriptional activator of amino acid biosynthetic genes in Saccharomyces cereuisiae. Mol. Cell. Biol. 6: 3990-3998.
Evidence for translational regulation of the activator of general amino acid control in yeast. Proc. Natl. Acad. Sci. USA 81: 6442-6446.
A hierarchy of trans-acting factors modulates translation of an activator of amino acid biosynthetic genes in Saccharomyces cerevisiae. Mol. Cell. Biol. 5 2349-2360. HINNEBUSCH, A. G., 1986 The general control of amino acid
biosynthetic genes in the yeast Saccharomyces cerevisiae. CRC Crit. Rev. Biochem. 21: 277-317.
HINNEBUSCH, A. G., and G. R. FINK, 1983 Positive regulation in the general amino acid control of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 8 0 5374-5378.
GCN4 protein, synthesized in vitro, binds HIS4 regulatory sequences: Implications for general control of amino acid biosynthetic genes in yeast. Cell 43: 177-
188.
ITO, H., Y. FUKUDA, K. MURATA and A. KIMURA, 1983 8516-8520.
DRISCOLL PE", M., G. THIREOS and H. GREER, 1984
520-528.
HARASHIMA, S., and A. G. HINNEBUSCH, 1986
HINNEBUSCH, A. G., 1984
HINNEBUSCH, A. G., 1985
HOPE, I. A., and K. STRUHL, 1985
Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 153: 163-168.
KATAOKA, T., S. POWERS, C. MCGILL, 0. FASANO, M. GOLFARB, J. STRATHERN, J. BROACH and M. WIGLER, 1984 Genetic anal- ysis of yeast R A S l and RASZ genes. Cell 37: 437-445.
LUCCHINI, G., A. G. HINNEBUSCH, C. CHEN and G. R. FINK, 1984 Positive regulatory interactions of the HIS4 gene of Saccharomyces cerevisiae. Mol. Cell. Biol. 4: 1326-1333.
Role of transfer ribonu- cleic acids in the regulation of several biosyntheses in Saccha- romyces cereuisiae. Eur. J. Biochem. 67: 335-339.
MUELLER, P. P. and A. G. HINNEBUSCH, 1986 Multiple upstream AUG codons mediate translational control of GCN4. Cell 45: 20 1-207.
MUELLER, P. P., S. HARASHIMA and A. G. HINNEBUSCH, 1987 A segment of GCN4 mRNA containing the upstream AUG co- dons confers translational control upon a heterologous yeast transcript. Proc. Natl. Acad. Sci. USA. 84: 2863-2867.
Identification and characterization of four new GCD genes in Saccharomyces cerevisiae. Curr. Genet. 1 0 657-664.
ROTHSTEIN, R. J., 1983 One-step gene disruption in yeast. Meth- ods Enzymol. 101: 202-21 1.
RYKOWSKI, M. C., J . W. WALLIS, J. CHOE and M. GRUNSTEIN, 1981 Histone 2B subtypes are dispensable during the yeast cell cycle. Cell 2 5 477-487.
SHERMAN, F., G. R. FINK and C. LAWRENCE, 1974 Methods of Yeast Genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
TATCHELL, K., D. CHALEFF, D. DEFEO-JONES and E. SCOLNICK, 1984 Requirement of either of a pair of ras-related genes of Saccharomyces cerevisiae for spore viability. Nature 3 0 9 523- 527.
5' Untranslated sequences are required for the translational control of a yeast regulatory gene. Proc. Natl. Acad. Sci. USA 81: 5096-5100. WOLFNER, M., D. YEP, F. MESSENGUY and G. R. FINK, 1975
Integration of amino acid biosynthesis into the cell cycle of Saccharomyces cerevisiae. J. Mol. Biol. 9 6 273-290.
Communicating editor: E. W. JONES
MESSENGUY, F., and J. DELFORGE, 1976
NIEDERBERGER, P., M. AEBIand R. HUETTER, 1986