The Roles of Klenow Processing and Flap Processing Activities of DNA
Polymerase I in Chromosome Instability in
Escherichia coli
K12 Strains
Yuki Nagata, Kazumi Mashimo, Masakado Kawata and Kazuo Yamamoto
1Department of Biomolecular Sciences, Graduate School of Life Sciences, Tohoku University, Sendai 980-8578, Japan Manuscript received June 11, 2001
Accepted for publication October 15, 2001
ABSTRACT
The sequences of spontaneous mutations occurring in the endogenoustonBgene ofEscherichia coliin the ⌬polA and polA107 mutant strains were compared. Five categories of mutations were found: (1) deletions, (2) minus frameshifts, (3) plus frameshifts, (4) duplications, and (5) other mutations. The ⌬polAstrain, which is deficient in both Klenow domain and 5⬘→3⬘exonuclease domain of DNA polymerase I, shows a marked increase in categories 1–4. The polA107 strain, which is deficient in the 5⬘ → 3⬘ exonuclease domain but proficient in the Klenow domain, shows marked increases in categories 3 and 4 but not in 1 or 2. Previously, we reported that thepolA1 strain, which is known to be deficient in the Klenow domain but proficient in the 5⬘→3⬘exonuclease domain, shows increases in categories 1 and 2 but not in 3 or 4. The 5⬘→3⬘exonuclease domain of DNA polymerase I is a homolog of the mammalian FEN1and the yeastRAD27flap nucleases. We therefore proposed the model that the Klenow domain can process deletion and minus frameshift mismatch in the nascent DNA and that flap nuclease can process plus frameshift and duplication mismatch in the nascent DNA.
D
NA polymerase I (PolI) ofEscherichia colihas three polA1strain and found marked increases in deletions enzymatic activities acting as a 5⬘→3⬘DNA poly- and frameshifts (Agemizuet al.1999). ThepolA1 muta-merase, a 3⬘ → 5⬘ exonuclease that edits 3⬘ terminal tion provides normal 5⬘→3⬘exonuclease activity with nucleotides of nascent DNA, and a 5⬘→3⬘exonuclease no Klenow activity (Joyceet al.1985). Inspection of the that removes nucleotides from the 5⬘ end of DNA or sequences around deletion end-points and frameshift RNA (KornbergandBaker1992). Proteolytic cleavage points indicated that these mutations occurred at G:C of PolI separates the polypeptide chain into two active base pair clusters; for example, among the 20 deletion fragments (Brutlaget al.1969): the large C-terminal sites recovered, ranging in sizes from 4 to 12,532 bp, 8 fragment called “Klenow” contains the 5⬘→3⬘polymer- sites were associated with the GC clusters at both ends ase and 3⬘ → 5⬘ exonuclease activities, and the small of the junctions and 10 at either end of the junction, N-terminal contains only 5⬘ →3⬘exonuclease activity. and among 6 minus frameshifts, 5 were associated with The 5⬘→3⬘exonuclease is a homolog of mammalian a GC cluster. Thus, there seems to be a resemblanceFEN1and yeast RAD27 flap endonucleases (Robinset between deletions and frameshifts recovered from the
al.1994). polA1 strain and the phenomenon known as instability
PolI is believed to be involved in both DNA replication of microsatellite repetitive sequences.
and repair (see Kornberg andBaker 1992). During Length variation of simple repetitive DNA sequences replication, PolI is thought to remove the RNA primer is known to be associated with human colon cancer (Aal-that initiates Okazaki fragments replacing short oligonu- tonen et al.1993; Ionovet al. 1993;Thibodeauet al. cleotides with DNA. During repair, PolI can bind to 1993). Human cells that exhibit microsatellite instability single-strand breaks in the phosphodiester backbone, are frequently deficient in mismatch repair (Kroutil excise the damaged region, and restore the integrity of et al.1996). A substantial fraction of familial colorectal the double helix. cancer is associated with microsatellite instability and Mutations in the polA gene have been shown to in- mutations in mismatch repair genes (Kolodner1995). fluence the frequencies of chromosomal deletion and Defects in DNA mismatch repair inE. coliand yeast re-minus frameshift mutations (Coukell and Yanofsky sult in an increase in the instability of repetitive sequences 1970; Vaccaro and Siegel 1975; Savic and Romac (Levinson and
Gutman 1987; Strand et al. 1993). 1982;Fixet al.1987). We investigated spontaneous
mu-Thus, the mismatch repair system can stabilize microsat-tagenesis in the endogenous tonB gene of the E. coli
ellites. However, about half of the sporadic tumor cell lines with microsatellite instability do not contain muta-tions in the mismatch repair genes, which suggests that
1Corresponding author:Department of Biomolecular Sciences,
Gradu-other genes may also contribute to microsatellite
insta-ate School of Life Sciences, Tohoku University, Sendai 9808578, Japan.
E-mail: [email protected] bility (Liuet al. 1995). Mutations in theRAD27
by interpolation from experimental determination of ro, the
myces cerevisiaegene, a homolog of 5⬘→3⬘exonuclease of
median number of ColBrcells determined among the cultures
PolI, also destabilize microsatellites by a mechanism
dis-by the formula ro ⫽ M(1.24 ⫹ lnM). Analysis of variance
tinct from mismatch repair (Johnsonet al. 1995;Tish- (ANOVA) was used to examine differences in mutation rate koffet al.1997; Kokoskaet al. 1998). A defect in the among the strainspolA⫹,polA1,⌬polA, andpolA107.
Sequencing of the mutant DNA:For thetonBmutation assay, 5⬘→3⬘exonuclease of PolI has been shown to lead to
independent colonies of KK102 (⌬polA::Kmr) or KK103
an increase in poly(AC) repeat tract expansion (Morel
(polA107) were inoculated in 3 ml of Luria broth at 37⬚
over-et al. 1998). These results strongly suggest that PolI may
night, and 100-l aliquots of these overnight cultures were contribute to prevent the instability of repetitive se- plated on colicin plates to obtain several hundred mutant quences as one of the factors other than the mismatch colonies per plate. To collect tonBmutants, only one colBr
colony was chosen from each colicin plate, an approach that repair system in E. coli.
ensured each mutant analyzed was of independent origin. In this study, we examined the sequence specificity of
The DNA fragment including the mutanttonBgene was ampli-spontaneous mutations in the endogenous tonB gene
fied by polymerase chain reaction (PCR) using appropriate of strains deficient in PolI activities. As PolI-deficient primers (Figure 1) from genomic DNA that had been ex-strains, we chose⌬polAandpolA107alleles, deficient in tracted from thetonBmutant. After amplification, the concen-tration of the amplified DNA was determined from the inten-both Klenow and 5⬘ → 3⬘ exonuclease domains and
sity of the band of the proper size on electrophoresis of 1-l 5⬘→3⬘exonuclease, respectively. Study of spontaneous
samples on 0.7% agarose gels. Mutant sequences were deter-mutations in the ⌬polA strain suggested an enhanced
mined by the dideoxy chain termination method using an frequency of deletion, minus frameshift, plus frameshift, ABI automated sequencer model 373A.
and duplication at GC cluster sites and in thepolA107
strain revealed an enhanced frequency of plus
frame-shift and duplication at GC cluster sites. RESULTS
To extend our previous study of the effects ofpolA1
mutation on spontaneous formation of tonB mutants MATERIALS AND METHODS
(Agemizu et al. 1999), we measured the efficiency of Bacterial strains: E. coli K12 strains KK102 (⌬polA::Kmr
spontaneous tonB mutations of KK102 (⌬polA) and zig219::Tn10) and KK103 (polA107 zig219::Tn10), which are
KK103 (polA107). In these cases, the polA1 mutation a derivative of polA⫹ strain KK1 (Wang et al. 1996), were
eliminates most of the Klenow activity but does not affect used for collecting mutations. KK1 carries the Cmrgene (cat),
locatedⵑ1.6 kb upstream of thetonBgene on the chromo- 5⬘→3⬘exonuclease activity (Joyceet al.1985), whereas some (Kitamuraet al. 1995). The⌬polA::Kmrallele,polA107
the polA107 mutant lacks 5⬘→ 3⬘ exonuclease activity allele, andzig219::Tn10allele were derived from strains CJ278
but retains almost normal Klenow activity (Heijneker (⌬polA::Kmr; Joyce and Grindley 1984), KL406 (polA107;
et al.1993). The⌬polAmutation is a null mutation of the National Institute of Genetics, Japan), and ET1214 (zig219::
polAgene (JoyceandGrindley1984). The presence of Tn10;Pahelet al. 1979), respectively. To construct isogenic
polAstrains, thezig219::Tn10allele was first transduced into polAmutations was verified by examining UV sensitivity. CJ278 or KL406 using P1 phage (Iharaet al. 1985) and UV- We first measured the spontaneous mutation rate of sensitive (UVs) and tetracycline-resistant (Tetr) derivatives
KK1 (polA⫹), KK101 (polA1), KK102 (⌬polA), and KK103 were screened. KK1 was infected with P1 phage grown in
(polA107), which were selected as the colBrphenotype.
resultant strains and selected for Tetrtransductants that were
screened for UVs. Colicinogenic E. coli strain CA18 carries The ColBr mutation rate of KK1 was 1.91 ⫻ 10⫺8; of
colicin B factor (IshiiandKondo1972). T1 bacteriophage KK101, 2.29 ⫻ 10⫺7; of KK102, 1.52 ⫻ 10⫺7; and of
were included in the colicin plates. KK103, 9.70⫻10⫺8(Table 1). ANOVA was used to
exam-Reagents and media:Luria broth, L agar, and phosphate
ine differences in mutation rates among four strains buffer were prepared as described byAkasakaandYamamoto
and significant differences could be found among them (1991). Colicin plates were prepared as described byUematsu
et al.(1999). Chloramphenicol (Cm; 30 g/ml), ampicillin (F3, 8⫽14.382 andP⫽0.0002). Fisher’s post-hoc test (50g/ml), or kanamycin (Km; 50g/ml) were included, if showed that mutation rates were significantly different necessary, in Luria broth or L agar. Enzymes and reagents used
betweenpolA⫹andpolA1(P⫽0.002),polA⫹and⌬polA
for DNA manipulation were purchased from Takara Shuzo
(P⫽0.0038), andpolA⫹andpolA107(P⫽0.046). (Kyoto, Japan) and Applied Biosystems (Foster City, CA).
A total of 65 independent ColBrmutated clones from
Mutation rate:Mutation to colicin B and T1 phage resis-tance (ColBr) was determined for 10 independent cultures
KK102 and 67 from KK103 were collected and used of KK1 (polA⫹), KK101 (polA1), KK102 (⌬polA), and KK103 for DNA sequencing. DNA sequence analysis yielded a (polA107) in 3 ml of Luria broth after overnight growth. For
mutant sequence in 51 and 51 of these clones of KK102 assays scoring ColBr, independent cultures were plated directly
and KK103, respectively. Thus, we were not able to iden-onto colicin plate. ColBr colonies were scored after 48 hr
tify the tonB gene mutation from 14 ColBr clones of
incubation at 37⬚. Total viable cells were determined by serial
dilution with phosphate buffer, followed by plating on L agar. KK102 and 16 ColBrclones of KK103. For comparison,
Mutation rates, expressed as mutations per cell per generation, their distribution by class is listed in Table 2 along with were calculated by the method of the median (Leaand
Coul-previously published results from polA1 strain UA1
son1949) using the following formula: mutation rate⫽M/N,
(Agemizuet al.1999) andpolA⫹strain TM31 (Kitamura whereMis the calculated number of mutation events andN
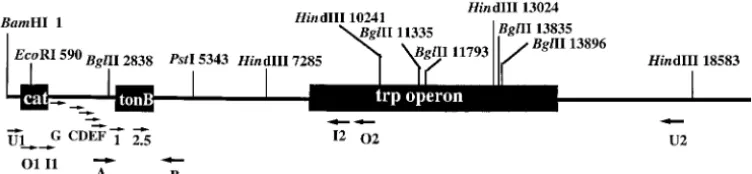
Figure1.—Physical map of min 28 ofE. colilinkage map tonB-trp region and the loca-tions of primers for PCR ampli-fication and sequencing ( Yama-muraet al.2000). Relevant re-striction sites with nucleotide numbers of the 19,495 bp DNA sequences in which 1 is the first G of the GATCBamHI site are included. In this numbering,catis from base pair 377–1036,tonBfrom base pair 2993–3727, and trpfrom base pair 14,864–8334. Arrows below the map indicate the primers used: A (5⬘-AAACGTGCTAAATGTGCCGG-3⬘, base pair 2702–2721), B (5⬘-CGATGTGGTGTGCTGCTATGC-3⬘, base pair 3953–3933), C (5⬘-GATCATCTTCAGGCAAGT-3⬘, base pair 1351–1368), D (5⬘-GGATTAAAACCTTCGCTTAA-3⬘, base pair 1689–1708), E (5⬘-ATGGAAAATATCAAGCAAAT-3⬘, base pair 2039–2058), F (5⬘-ATTATCTGCTTGTGGTGGTG-3⬘, base pair 2389–2404), G (5⬘-GGCTTCCATGTCGGCAGAAT-3⬘, base pair 968– 987), 1 (5⬘-TAAGGCCATGCATAAAGT-3⬘, base pair 2868–2885), 2.5 (5⬘-AGAAGCACCGGTGGTCA-3⬘, 3256–3272), O1 (5⬘-CCG GCCTTTATTCACATTCTTGCCCGCCTGATGAATGCTC-3, base pair 545–584), O2 (5⬘-CTGGCCCGCATCCTTATCCGACCATT GTGCGTGAGTTTC-3⬘, base pair 9760–9722), I1 (5⬘-ATCCGGAATTCCGTATGGCAATGAAAGACGGTGAG-3⬘, base pair 585– 619), I2 (5⬘-AGCGGATGATTGGCGAAGAAACCAAAGCGCAGATT-3⬘, base pair 9721–9687), U1 (5⬘-GCAGAATAAATAAATCCTGG TGTCCCTGTTGATACCGG-3⬘, base pair 200–237), U2 (5⬘-AGGCTTGTCATCATCGCGGGCAAACAAAAGCTCAAGGG-3⬘, base pair 18029–17992), I1 (5⬘-ATCCGGAATTCCGTATGGCAATGAAAGACGGTGAG-3⬘, base pair 585–619), I2 (5⬘-AGCGGATGATT GGCGAAGAAACCAAAGCGCAGATT-3⬘, base pair 9721–9687).
KK102 and KK103. The most frequent mutational event different sites, ranging in size from 4 to 102 bp. Six of these sites were flanked by repeated sequences (Table in the⌬polAstrain was deletion followed by⫺1
frame-shift, base substitution, and⫹1 frameshift in that order. 3, boldface letters) of two or more bases, implying a role of direct repeats for deletion formation (Albertini The types of mutation detected in the⌬polAstrain were
considerably different from those of thepolA107strain et al.1982). Four of the 12 deletion sites showed 2- to 4-bp repeats (Table 3, italic letters) in the immediate where ⫹1 frameshift dominated followed by
duplica-tion, deleduplica-tion, base substituduplica-tion, and⫺1 frameshift in vicinity of the endpoints, which appeared to indicate no clear association with deletion. Formation of deletion that order.
Deletion: As shown in Tables 1 and 2, the rates of endpoints between these homologies can be explained if it is assumed that one or two extra bases have been deletions in thepolA1strain (22.89⫻10⫺8⫻39/61⫽
14.65⫻10⫺8) and the⌬polAstrain (15.19⫻10⫺8⫻21/ added or deleted (Uematsuet al.1999). Three deletion
sites had no homology at the endpoint. Further analysis 51⫽6.23⫻10⫺8) were 38.5- and 16.4-fold, respectively,
higher than that of thepolA⫹strain (1.91⫻10⫺8⫻10/ of the DNA sequences at the sites of deletion events of
the ⌬polA strain revealed that six had a GC cluster 51 ⫽ 0.38 ⫻ 10⫺8). Table 3 shows the 21 deletions
identified from the⌬polAstrain, among which 12 were (⬎4-bp repeats; Table 3) within 3 bp of both endpoints.
TABLE 1
Rates of ColBrcells inpolAstrains
ColBrmutation rate
Relevant Median no. (mutations/cell/generation) Average Strains genotype of mutants (⫻108)a (⫻108)b
KK1 polA⫹ 116 3.01 1.91⫾0.95
35.5 1.31
57 1.41
KK101 polA1 440.5 18.25 22.89⫾4.62
456 22.93
885 27.48
KK102 ⌬polA 254 15.86 15.19⫾4.54
318.5 19.36
165 10.36
KK103 polA107 439.5 12.83 9.70⫾4.75
296 12.04
143.5 4.23
aIn each experiment, the rates of ColBrcells were determined by measuring the number of ColBrcells in
10 independent cultures and converting these numbers into a mutation rate by the method of the median (LeaandCoulson1949). Mutation rates for each strain represent averages among three sets of 10 independent cultures.
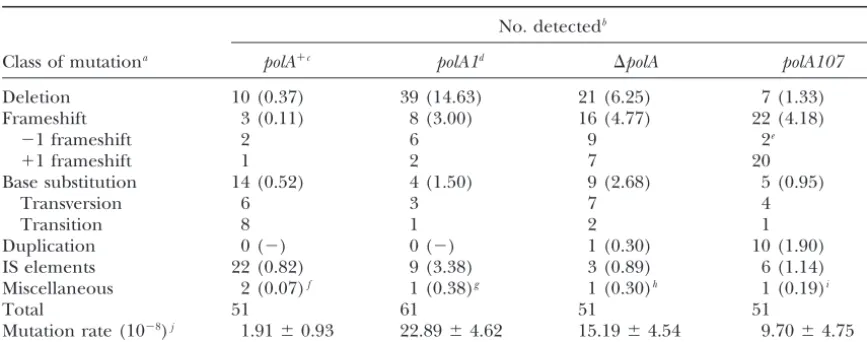
TABLE 2
Spontaneous endogenoustonBmutations inpolAstrains
No. detectedb
Class of mutationa polA⫹c polA1d ⌬polA polA107
Deletion 10 (0.37) 39 (14.63) 21 (6.25) 7 (1.33) Frameshift 3 (0.11) 8 (3.00) 16 (4.77) 22 (4.18)
⫺1 frameshift 2 6 9 2e
⫹1 frameshift 1 2 7 20
Base substitution 14 (0.52) 4 (1.50) 9 (2.68) 5 (0.95)
Transversion 6 3 7 4
Transition 8 1 2 1
Duplication 0 (⫺) 0 (⫺) 1 (0.30) 10 (1.90)
IS elements 22 (0.82) 9 (3.38) 3 (0.89) 6 (1.14) Miscellaneous 2 (0.07)f 1 (0.38)g 1 (0.30)h 1 (0.19)i
Total 51 61 51 51
Mutation rate (10⫺8)j 1.91⫾0.93 22.89⫾4.62 15.19⫾4.54 9.70⫾4.75
aIn this table,⫺frameshift refers to the deletion of one or two bases. A more than three-base deletion is
defined as a deletion. A⫹frameshift refers to the addition of one base. A more than two-base addition with base duplication is defined as a duplication.
bParentheses (⫻10⫺8) indicate the mutation rate of each subclass that was calculated as described in the
text.
cData, except for mutation rate, fromKitamuraet al.(1995). In this case, TM31 (polA⫹; AB1157 derivative)
instead of KK1 (polA⫹) was used.
dData, except for mutation rate, fromAgemizuet al.(1999). In this case, UA1 (polA1; CH931 derivative)
instead of KK101 (polA1) was used.
eIncludes one two-base deletion.
fOne GC→TG (nucleotides 3500 and 3501) and one four-base addition (boldface letters) at sequence GC
(nucleotides 3040 and 3041) to GGTTTC (Kitamuraet al.1995).
gInversion with G deletion (boldface letters) at sequence AGGTTAAAGT to AGTTTAAGT (nucleotides
3533–3538), which was counted as one mutation (Agemizuet al.1999).
hA 13-bp deletion in brackets with a G addition (boldface letter) at sequence GATTTA[CCTCGCCGC
TTCC]C to GGATTTAC (nucleotides 3023–3032) was counted as one mutation.
iT to A transversion and C deletion (boldface letters) at sequence GCGTTA to GGTAA (nucleotides 3647
and 3650), which was counted as one mutation.
jData from Table 1.
In addition, five sites had a GC cluster within 3 bp (polA107) was 25.9-, 41.0-, and 36.3-fold higher than that in thepolA⫹strain, respectively. Previous studies of of one endpoint. Furthermore, 7 of the 12 sites had
imperfect inverted repeats at one or both ends of the spontaneous mutagenesis in thepolA1strain (Vaccaro and Siegel 1975; Savic and Romac 1982; Fix et al.
endpoints (Table 3, arrow). Among 21 deletions, 1
par-ticular deletion (13 bp, nucleotides 3023–3035) ac- 1987; Agemizuet al. 1999) suggested the presence of minus frameshifts. Table 2 shows that 9 of 16 frameshifts counted for nine incidences. This site was also a very
strong deletion hotspot site in the polA1 strain (17 in the ⌬polA strain resulted from the loss of a base pair and 7 were due to a base pair gain. Two minus among 39 deletions;Agemizuet al.1999).
Table 3 also shows seven deletions identified in the frameshifts among 22 recovered in the present study of thepolA107strain were observed and 20 plus frameshifts KK103 (polA107) strain. Spontaneous deletion rate of
KK103 was 1.33 ⫻ 10⫺8 (9.70⫻ 10⫺8 ⫻ 7/51), which were also observed. Thus, the polA107 strain is a plus
frameshift mutator and the⌬polAstrain is a plus/minus was not significantly different from that observed in the
polA⫹strain (2⫽0.454,P⫽0.499). All of the deletions frameshift mutator. On the other hand, thepolA1strain
is a minus frameshift mutator. were flanked by repeated sequences. In addition, all the
deletions of thepolA107strain had a GC cluster at both The DNA sequences surrounding each of the minus and plus frameshift mutations recovered are depicted of the deletion endpoints and had imperfect inverted
repeats. Deletion at nucleotides 3395–3496 was counted in Tables 4 and 5, respectively. Table 4 indicates that four of nine cases of minus frameshifts recovered from three times and was also detected once in the ⌬polA
strain. Deletion at nucleotides 3437–3474 was detected the ⌬polA strain could be explained on the basis of slippage events in runs of direct sequences (Strei-in both KK102 and KK103 stra(Strei-ins.
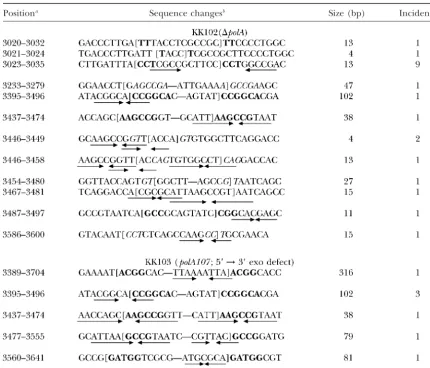
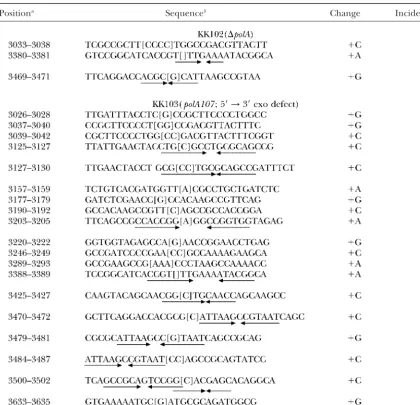
TABLE 3
Target sequences of deletion mutations in a⌬polAorpolA107strain
Positiona Sequence changesb Size (bp) Incidence
aNumbering is in accordance with Figure 1.
bThe precise positions of the breakpoints (in brackets) in those cases involving direct repeats are ambiguous
and could lie anywhere within the repeated sequences (boldface letters). The breakpoint shown represents only one possibility. The repeated sequences involved in deletion are in boldface letters, and those not involved are shown in italics. The inverted arrows under the sequence indicate imperfect inverted repeats.
two cases in the polA107 strain occurred at nonruns 10 of 19 sites of thepolA107strain plus frameshifts were associated with imperfect inverted repeats.
(Table 4). Eight of 9 minus frameshifts in the ⌬polA
strain were associated with GC clusters (Table 4). All Duplication:Whereas duplications were not recovered in wild-type orpolA1strains, 1 duplication was counted the minus frameshifts, except 3082–3084, were also
asso-ciated with imperfect inverted repeats. Table 5 illus- in the ⌬polAstrain and 10 duplications in thepolA107
strain (Table 2). The location and types of duplication trates 7 plus frameshifts identified in the⌬polAstrain
and 20 plus frameshifts in thepolA107strain. Among 7 events are presented in Table 6. Among 10 duplications in the polA107 strain ranging in size from 2 to 37 bp plus frameshifts in the⌬polAstrain, one event, addition
of A between nucleotides 3380–3381, was independently with the majority being 2 bp in length, 3 duplications (3097–3104, 3405–3443, and 3533–3538) occurred at recovered five times. Thus, 3 sites were detected in the
⌬polAstrain plus frameshifts, among which 1 was associ- sites flanking repeated sequences. All the duplications, including the above 3 duplications except 3167–3168 ated with a run of 4 Cs to form a run of 5 Cs and the
re-maining 2 were not associated with runs of bases. Among and 3634–3635, have mutation sites with a cluster of di-rect repeats or imperfect inverted repeats. These se-19 plus frameshift sites recovered from thepolA107strain,
6 were associated with a run of bases. Eighteen plus frame- quence characteristics of duplication are essentially the same as the characteristics of frameshift and deletion shift sites in thepolA107strain were associated with GC
TABLE 4
Location and types of minus frameshift mutations in a⌬polAorpolA107strain
Positiona Sequenceb Change Incidence
aNumbering is in accordance with Figure 1 in which 2993 is the first A of the ATG start and 3727 is the
last A of the TAA end of thetonBgene.
bWild-type sequence. One or two bases indicated in brackets are lost. The inverted arrows under the sequence
indicate imperfect inverted repeat.
generated by slip-back misalignment of the newly syn- minus frameshifts, (3) plus frameshifts, and (4) duplica-tions. The orders of the occurrences of these categories thesized fragments of nucleotides at the potentially
stacked inverted repeats as suggested byIkehataet al. were different from different alleles of the polA gene. The order of occurrence by category in the⌬polAstrain (1989). It is thus evident that the polA107 strain is a
duplication mutator. was deletions, minus frameshifts, plus frameshifts, and duplications, while that inpolA107was plus frameshifts
Base substitution: We observed nine and five base
substitutions among the 51 mutations detected in and duplications with no increases in deletions or minus frameshifts as compared to the wild-type strain. In the KK102 and KK103, respectively, giving a base
substitu-tion rate of 15.19 ⫻ 10⫺8 ⫻ 0.18 ⫽ 2.73 ⫻ 10⫺8 and polA1strain, deletions and minus frameshifts occurred
predominantly in this order with no increases in duplica-9.70⫻10⫺8⫻0.10⫽0.970⫻10⫺8, respectively, which
was essentially the same as that in thepolA⫹cells (1.91⫻ tions or plus frameshifts. Thus, there was a quite clear parallel between the categories of mutations and defects 10⫺8⫻0.27⫽0.516⫻10⫺8; Table 2). Therefore,⌬polA
andpolA107mutations do not seem to affect base substi- of domains in PolI, Klenow, and 5⬘ →3⬘exonuclease. As summarized in Table 8, the Klenow domain is in-tution mutagenesis in the endogenoustonBgene. This
conclusion is consistent with that of analysis of base volved in preventing deletions and minus frameshifts and the 5⬘ → 3⬘ exonuclease domain is involved in substitution in thepolA1strain (Table 2 andAgemizu
et al. 1999). Table 7 shows the location and types of preventing duplications and plus frameshifts.
base substitution mutations. The sequence analysis in this study using the⌬polA
and polA107 strains together with our previous study using thepolA1strain (Agemizuet al.1999) indicates a DISCUSSION
sequence specificity surrounding the mutation sites. Ten of 12 sites of deletions, 8 of 9 sites of minus Using⌬polAandpolA107alleles, together with
previ-frameshifts and 2 of 3 sites of plus previ-frameshifts recovered ous results obtained in thepolA1mutant strain, we
exam-from the⌬polAstrain were strongly associated with GC ined spontaneously occurring sequence changes in the
clusters. Seventeen of 19 sites of plus frameshifts and 5 endogenoustonBgene. It was shown that spontaneous
of 10 sites of duplications recovered from thepolA107
mutations different from the wild-type strain are
TABLE 5
Location and types of plus frameshift mutations in a⌬polAorpolA107strain
Positiona Sequenceb Change Incidence
aNumbering is in accordance with Figure 1 in which 2993 is the first A of the ATG start and 3727 is the
last A of the TAA end of thetonBgene.
bWild-type sequence. Brackets indicate the site where one base was gained. The inverted arrows under the
sequence indicate imperfect inverted repeats.
Furthermore, in thepolA1strain, GC clusters were seen found in eukaryotes and prokaryotes. As mentioned in the Introduction, microsatellite length variation is known in 18 of 20 sites of deletions and 5 of 6 sites of minus
frameshifts. The GC cluster results in formation of a to be associated with a number of genetic diseases in-cluding colon cancer (Aaltonenet al.1993;Ionovet al.
cluster of direct repeats as well as inverted repeats
(Ta-bles 3–7). Thus, complex secondary structure adjoined 1993; Thibodeau et al. 1993; Richards and Suther-land1994;AshleyandWarren1995). Previous studies its endpoints, and misalignment facilitated by this
sec-ondary structure may be responsible for the occurrence in E. coli and yeast have shown that defects in DNA mismatch repair result in an increase in the instability of deletions, frameshifts, and duplications observed in
polAstrains (BzymekandLovett2001). of repetitive sequences (LevinsonandGutman1987; Strandet al.1993). However, the observation that mi-The observation that far more than half of the
dele-tion, minus frameshift, plus frameshift, and duplication crosatellite instability occurs often in cells with a func-tional mismatch repair system suggested that other genes events recovered from thepolAmutant strains have
end-points in the GC cluster sequence suggested that GC may contribute to prevent repetitive sequence instabil-ity. Here, we suggested that thepolA⫹gene also stabilizes clusters inE. coli are quite unstable repetitive DNA
TABLE 6
Location and types of duplication mutations in a⌬polAorpolA107strain
Positiona Sequenceb Base pairs gained (bp)
aNumbering is in accordance with Figure 1.
bWild-type sequence. Brackets with boldface letters indicate bases duplicated. The inverted arrows under the sequence indicate
imperfect inverted repeats.
suggests two points: (1) the same mechanisms may be mismatch in the nascent DNA, the 5⬘→3⬘exonuclease effects nascent DNA mismatch removal by flap nuclease operating to facilitate addition or deletion of bases at
tandemly repeated sequence motifs (microsatellites) (Figure 2A). On the other hand, sequence shortening in the nascent DNA would form a mismatch bulge in and sequences without any motifs, the only difference
being the frequency of occurrence, and (2) 5⬘ → 3⬘ the template DNA. In this case, 3⬘→5⬘exonuclease of Klenow digests nascent DNA to solve sequence variation exonuclease of PolI processes the mismatch
intermedi-ate that will facilitintermedi-ate length expansion, plus frameshift intermediates (Figure 2B). In this case, it is assumed that sequence expansion/shortening occurs during the and duplication, and Klenow of PolI processes the
mis-match intermediate that will facilitate length shorten- DNA replication step, not during the postreplication step (see laterdiscussion).
ing, minus frameshift and deletion.
PolI, encoded by the polA gene, functions in both RAD27mutants ofS. cerevisiaeshow a marked increase in repeat expansion (Johnson et al.1995;Kokoskaet
repair and replication (seeKornbergandBaker1992).
In the absence of exogenous DNA-damaging agents, the al. 1998). The RAD27 gene encodes a 5⬘ → 3⬘ exo-nuclease with structural and functional homology to processing of Okazaki fragments during lagging-strand
synthesis is probably its main function. We have shown the mammalianFEN1and to the 5⬘→ 3⬘ exonuclease domain of PolI (Robinset al.1994). These nucleases, in this study that deletion of 5⬘ → 3⬘ exonuclease
in-creases the rate of sequence expansion and that of known to function in lagging-strand synthesis, remove the 5⬘flap generated by displacement synthesis by endo-Klenow sequence shortening. Structural intermediates
of repetitive sequence variation in this case may be nucleolytic cleavage (Robinset al.1994). The mutation of 5⬘ → 3⬘ exonuclease in E. coli leads to a marked formed during lagging-strand synthesis. It was
demon-strated previously that replication-dependent frameshift increase in repeat expansion that also was found in RAD27 mutants. Finally, Morel et al. (1998) demon-mutagenesis occurred in the lagging strand (Iwakiet
al. 1996). Thus, we propose a model that interprets strated that deletion in 5⬘→3⬘exonuclease of PolI led to a marked increase in poly(AC) repeat expansion. processing sequence variation by PolI (Figure 2).
Dur-ing laggDur-ing-strand synthesis, sequence expansion occurs These results are consistent with a role for this class of 5⬘→3⬘exonuclease in removing single-strand nascent in the nascent DNA, which forms a mismatch bulge in
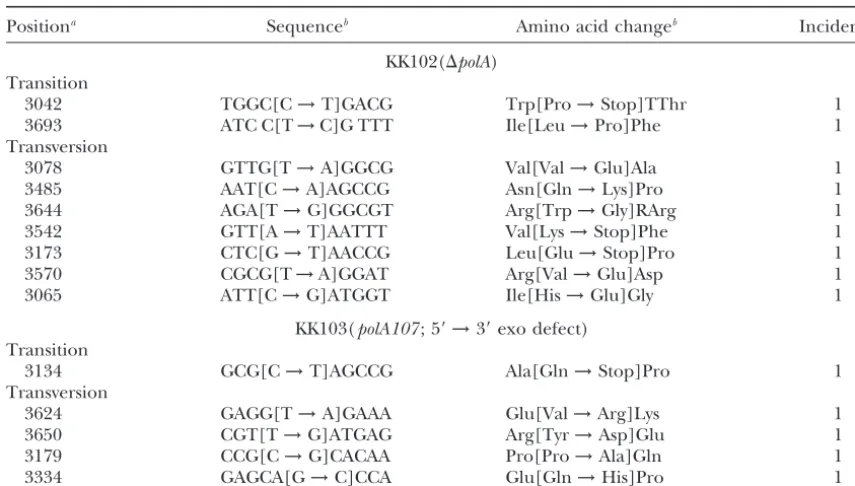
TABLE 7
Location and types of base substitution mutations in a⌬polAorpolA107strain
Positiona Sequenceb Amino acid changeb Incidence
KK102(⌬polA) Transition
3042 TGGC[C→T]GACG Trp[Pro→Stop]TThr 1
3693 ATC C[T→C]G TTT Ile[Leu→Pro]Phe 1
Transversion
3078 GTTG[T→A]GGCG Val[Val→Glu]Ala 1
3485 AAT[C→A]AGCCG Asn[Gln→Lys]Pro 1
3644 AGA[T→G]GGCGT Arg[Trp→Gly]RArg 1
3542 GTT[A→T]AATTT Val[Lys→Stop]Phe 1
3173 CTC[G→T]AACCG Leu[Glu→Stop]Pro 1
3570 CGCG[T→A]GGAT Arg[Val→Glu]Asp 1
3065 ATT[C→G]ATGGT Ile[His→Glu]Gly 1
KK103(polA107; 5⬘→3⬘exo defect) Transition
3134 GCG[C→T]AGCCG Ala[Gln→Stop]Pro 1
Transversion
3624 GAGG[T→A]GAAA Glu[Val→Arg]Lys 1
3650 CGT[T→G]ATGAG Arg[Tyr→Asp]Glu 1
3179 CCG[C→G]CACAA Pro[Pro→Ala]Gln 1
3334 GAGCA[G→C]CCA Glu[Gln→His]Pro 1
aNumbering is in accordance with Figure 1 in which 2993 is the first A of the ATG start and 3727 is the
last A of the TAA end of thetonBgene.
bBases and amino acid residues shown at the left and the right in the brackets are those of the normal and
mutant genes, respectively.
Recently,Kokoskaet al.(1998) andJinet al.(2001) flap DNA with this region can produce a DNA molecule with unpaired repeats. The unpaired repeats on the examined the effects on POL3 mutation ofS. cerevisiae
on the stability of microsatellites and found the in- nascent strand can lead to additions. Alternatively, the terminal ribonucleotide on the Okazaki fragment can creased rate of deletion formation. POL3 codes for DNA
polymerase␦, which has 5⬘→3⬘DNA polymerase and lead to DNA synthesis block and activation of 3⬘→ 5⬘ exonuclease without DNA flap. If a secondary structure 3⬘ → 5⬘ exonuclease activities (Morrison et al. 1991;
Simonet al.1991). Taking together the results of RAD27 is formed on the resulting single-strand region, subse-quent synthesis across the gap can lead to a mismatch mutants and POL3 mutants, these authors proposed a
model as follows: during the postreplication Okazaki repeat on the template strand. This mismatch can lead to deletion. In this model, RAD27 and POL␦have anti-fragment processing step, because of the delay to
re-move the terminal ribonucleotide due to RAD27 muta- mutator roles to prevent unpaired mismatch formation during postreplication Okazaki fragment processing. tion, displacement of the end of the Okazaki fragment
by continued synthesis from an adjacent primer results Tishkoffet al. (1997) proposed an essentially similar model. The present model presented in Figure 2 shows in the formation of a DNA flap that activates the 3⬘→
5⬘exonuclease of DNA polymerase, producing a single- that 5⬘→3⬘exonuclease and Klenow activities are con-sidered to involve the removal of polymerization errors. strand gap adjacent to the flap. Reassociation of the
TABLE 8
Correlation between PolI domain and categories oftonBmutations
PolI activity Categories of mutations
Allele Klenow 5⬘→3⬘exonuclease Deletions Minus fs Plus fs Duplication
polA1 ⫺ ⫹ ⫹ ⫹ ⫺ ⫺
⌬polA ⫺ ⫺ ⫹ ⫹ ⫹ ⫾
polA107 ⫹ ⫺ ⫺ ⫺ ⫹ ⫹
deletion (dinB-yafN) allele that inactivates downstream
yafOandyafP(Courcelleet al.2001) was used. Thus, the evidence for PolIV is ambiguous (McKenzieet al.
2001). And thus, whether PolIII alone is involved in repetitive sequence instability or PolII, PolIV, or PolV are also involved remains to be determined.
We thank Drs. A. Nishimura (National Institute of Genetics) and C. M. Joyce (Yale University) forE. colistrains. We also thank two anonymous reviewers for comments and suggestions on the manu-script. This work was supported by Grants-in-Aid for Scientific Re-search from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
LITERATURE CITED
Aaltonen, L. A., P.Peltomaki, F. S.Leach, P.Sistonen, L. Pylkka-nenet al., 1993 Clues to the pathogenesis of familial colorectal cancer. Science260:812–816.
Agemizu, Y., N. UematsuandK. Yamamoto, 1999 DNA sequence analysis of spontaneoustonBdeletion mutations in apolA1strain ofEscherichia coliK12. Biochem. Biophys. Res. Commun.261:
584–589.
Akasaka, S., andK. Yamamoto, 1991 Construction ofEscherichia coliK12phrdeletion and insertion mutants by gene replacement. Mutat. Res.254:27–35.
Albertini, A. M., M. Hofer, M. P. CalosandJ. H. Miller, 1982 On the formation of spontaneous deletions: the importance of short sequence homologies in the generation of large deletions. Cell29:319–328.
Ashley, C. T.,Jr., andS. T. Warren, 1995 Trinucleotide repeat
expansion and human disease. Annu. Rev. Genet.29:703–728.
Figure2.—Processing of length variation during lagging- Bebenek, K., C. M. Joyce, M. P. FitzgeraldandT. A. Kunkel, 1990 The fidelity of DNA synthesis catalyzed by derivatives ofEscherichia strand synthesis by DNA polymerase I. The model assumes
coliDNA polymerase I. J. Biol. Chem.265:13878–13887.
that mismatch bulges will lead to plus frameshift or duplication
Bonner, C. A., S. K. Randall, C. Rayssiguier, M. Radman, R. Eritja
if present on the nascent strand (A) or to minus frameshift
et al., 1988 Purification and characterization of an inducible
or deletion if present on the template strand (B). Escherichia coliDNA polymerase capable of insertion and bypass
at abasic lesions in DNA. J. Biol. Chem.263:18946–18952. Brutlag, D., M. R. Atkinson, P. SetlowandA. Kornberg, 1969
An active fragment of DNA polymerase produced by proteolytic
The deletions, frameshifts, and duplications recovered
cleavage. Biochem. Biophys. Res. Commun.37:982–989.
in this study were observed within GC-rich sequences Bzymek, M., andS. T. Lovett, 2001 Instability of repetitive DNA
sequences: the role of replication in multiple mechanisms. Proc.
(Tables 3–7). These repetitive sequences support the
Natl. Acad. Sci. USA98:8319–8325.
formation of secondary structures. Thus, the replication
Coukell, M. B., andC. Yanofsky, 1970 Increased frequency of
of repeats that have such a secondary structure by DNA deletions in DNA polymerase mutants ofEscherichia coli. Nature 228:633–635.
polymerase III holoenzyme (PolIII) may lead to
addi-Courcelle, J., A. Khodursky, B. Peter, P. O. Brownand P. C.
tion or deletion of bases. This argument is supported
Hanawalt, 2001 Comparative gene expression profiles
follow-by the finding that frameshift mutations were found in ing UV exposure in wild-type and SOS-deficientEscherichia coli.
Genetics158:41–64.
products ofin vitroDNA synthesis with weakly processive
De Boer, J. G., andL. S. Ripley, 1988 Anin vitroassay for frameshift
DNA polymerases or replicative DNA polymerases that
mutations: hotspots for deletions of 1 bp by Klenow-fragment
lack proofreading capacities (Kunkel 1986; de Boer polymerase share a consensus DNA sequence. Genetics 118:
181–191.
andRipley1988;Bebeneket al.1990;MoandSchaaper
Fix, D. F., P. A. BurnsandB. W. Glickman, 1987 DNA sequence
1996). Thus, repetitive sequence instability may be
analysis of spontaneous mutation in apolA1strain ofEscherichia caused by PolIII replication errors. As mentioned above, coli indicates sequence-specific effects. Mol. Gen. Genet.207:
267–272.
SOS induction is a microsatellite-destabilizing factor. In
Heijneker, H. L., D. J. Ellens, R. H. Tjeerde, B. W. Glickman, B.
E. coli, three DNA polymerases, PolII (polB;Bonneret al.
Van Dorpet al., 1973 A mutant ofEscherichia coliK12 deficient
1988), PolIV (dinB;Wagneret al.1999), and PolV (umuCD; in the 5⬘-3⬘exonucleolytic activity of DNA polymerase I. II.
Purifi-cation and properties of the mutant enzyme. Mol. Gen. Genet.
Reuvenet al.1999;Tanget al.1999), are induced as part
124:83–96.
of the SOS response.Napolitanoet al.(2000)
demon-Ihara, M., Y. OdaandK. Yamamoto, 1985 Convenient construction
strated that PolII and PolV are necessary and sufficient of strain useful for transducingrecAmutations with bacteriophage
P1. FEMS Microbiol. Lett.30:33–35.
for⫺1 and ⫺2 frameshift mutagenesis induced by the
Ikehata, H., T. Akagi, H. Kimura, S. AkasakaandT. Kato, 1989
chemical mutagensN-2-acetylaminofluoren, respectively,
Spectrum of spontaneous mutations in a cDNA of the human
and benzo(␣)pyrene-induced⫺1 frameshift mutagene- hprtgene integrated in chromosomal DNA. Mol. Gen. Genet.
219:349–358.
Ionov, Y., M. A. Peinado, S. Malkhosyan, D. ShibataandM. Peru- instability inEscherichia coli. Proc. Natl. Acad. Sci. USA95:10003– 10008.
cho, 1993 Ubiquitous somatic mutations in simple repeated
sequences reveal a new mechanism for colonic carcinogenesis. Morrison, A., J. B. Bell, T. A. KunkelandA. Sugino, 1991
Eukary-otic DNA polymerase amino acid sequence required for 3⬘→5⬘
Nature363:558–561.
Ishii, Y., andS. Kondo, 1972 Spontaneous and radiation-induced exonuclease activity. Proc. Natl. Acad. Sci. USA88:9473–9477. Napolitano, R., R. Janel-Bintz, J. WagnerandR. P. Fuchs, 2000 deletion mutations inEscherichia colistrains with different DNA
repair capacities. Mutat. Res.16:13–25. All three SOS-inducible DNA polymerases (Pol II, Pol IV and Pol
V) are involved in induced mutagenesis. EMBO J.19:6259–6265. Iwaki, T., A. Kawamura, Y. Ishino, K. Kohno, Y. Kanoet al., 1996
Preferential replication-dependent mutagenesis in the lagging Pahel, G., F. R. BloomandB. Tyler, 1979 Deletion mapping of
thepolA-metBregion of theEscherichia colichromosome. J. Bacte-DNA strand inEscherichia coli. Mol. Gen. Genet.251:657–664.
Jin, Y. H., R. Obert, P. M. Burgers, T. A. Kunkel, M. A. Resnick riol.138:653–656.
Reuven, N. B., G. Arad, A. Maor-ShoshaniandZ. Livneh, 1999
et al., 2001 The 3⬘→5⬘exonuclease of DNA polymerase␦can
substitute for the 5⬘flap endonuclease Rad27/Fen1 in processing The mutagenesis protein UmuC is a DNA polymerase activated
by UmuD⬘, RecA, and SSB and is specialized for translesion repli-Okazaki fragments and preventing genome instability. Proc. Natl.
Acad. Sci. USA98:5122–5127. cation. J. Biol. Chem.274:31763–31766.
Richards, R. I., andG. R. Sutherland, 1994 Simple repeat DNA Johnson, R. E., G. K. Kovvali, L. PrakashandS. Prakash, 1995
Requirement of the yeast RTH15⬘ to 3⬘ exonuclease for the is not replicated simply. Nat. Genet.6:114–116.
Robins, P., D. J. Pappin, R. D. WoodandT. Lindahl, 1994 Struc-stability of simple repetitive DNA. Science269:238–240.
Joyce, C. M., andN. D. Grindley, 1984 Method for determining tural and functional homology between mammalian DNase IV and the 5⬘-nuclease domain of Escherichia coli DNA polymerase whether a gene ofEscherichia coliis essential: application to the
polAgene. J. Bacteriol.158:636–643. I. J. Biol. Chem.269:28535–28538.
Savic, D. J., andS. P. Romac, 1982 Powerful mutator activity of the Joyce, C. M., D. M. Fujii, H. S. Laks, C. M. Hughes andN. D.
Grindley, 1985 Genetic mapping and DNA sequence analysis polA1mutation within the histidine region ofEscherichia coliK-12. J. Bacteriol.149:955–960.
of mutations in thepolAgene ofEscherichia coli. J. Mol. Biol.186:
283–293. Simon, M., L. GiotandG. Faye, 1991 The 3⬘→5⬘exonuclease
ac-tivity located in the DNA polymerase delta subunit of
Saccharo-Kitamura, K., Y. Torii, C. MatsuokaandK. Yamamoto, 1995 DNA
myces cerevisiaeis required for accurate replication. EMBO J.10:
sequence changes in mutations in thetonBgene on the
chromo-2165–2170.
some of Escherichia coli K12: insertion elements dominate the
Strand, M., T. A. Prolla, R. M. LiskayandT. D. Petes, 1993 Desta-spontaneous spectra. Jpn. J. Genet.70:35–46.
bilization of tracts of simple repetitive DNA in yeast by mutations Kokoska, R. J., L.Stefanovic, H. T.Tran, M. A.Resnick, D. A.
affecting DNA mismatch repair. Nature365:274–276.
Gordeninet al., 1998 Destabilization of yeast micro- and
minisa-Streisinger, K., T. Okada, J. Emrich, J. Newton, A. Tsugitaet al., tellite DNA sequences by mutations affecting a nuclease involved
1966 Frameshift mutations and the genetic code. Cold Spring
in Okazaki fragment processing (rad27) and DNA polymerase␦
Harbor Symp. Quant. Biol.31:77–84.
(pol3-t). Mol. Cell. Biol.18:2779–2788.
Tang, M., X. Shen, E. C. Frank, M. O’Donnell, R. Woodgateet
Kolodner, R. D., 1995 Mismatch repair: mechanisms and
relation-al., 1999 UmuD⬘2C is an error-prone DNA polymerase,
Esche-ship to cancer susceptibility. Trends Biochem. Sci.20:397–401.
richia colipol V. Proc. Natl. Acad. Sci. USA96:8919–8924. Kornberg, A., andT. A. Baker, 1992 DNA polymerase I ofE. coli,
Thibodeau, S. N., G. BrenandD. Schaid, 1993 Microsatellite insta-pp. 113–159 inDNA Replication, Ed. 2. W. H. Freeman, New York.
bility in cancer of the proximal colon. Science260:816–819. Kroutil, L. C., K. Register, K. BebenekandT. A. Kunkel, 1996
Tishkoff, D. X., N. Filosi, G. M. GaidaandR. D. Kolodner, 1997 Exonucleolytic proofreading during replication of repetitive
A novel mutation avoidance mechanism dependent onS. cerevisiae
DNA. Biochemistry35:1046–1053.
RAD27is distinct from DNA mismatch repair. Cell88:253–263. Kunkel, T. A., 1986 Frameshift mutagenesis by eucaryotic DNA
Uematsu, N., C. Matsuoka, Y. Agemizu, E. NagoshiandK. Yama-polymerases in vitro. J. Biol. Chem.261:13581–13587.
moto, 1999 Asymmetric crossing over in the spontaneous for-Lea, D. E., andC. A. Coulson, 1949 The distribution of the numbers
mation of large deletions in thetonB-trpregion of theEscherichia
of mutants in bacterial populations. J. Genet.49:264–285.
coliK-12 chromosome. Mol. Gen. Genet.261:523–529.
Levinson, G., and G. Gutman, 1987 High frequency of short
Vaccaro, K. K., andE. C. Siegel, 1975 Increased spontaneous rever-frameshifts in poly-CA/GT tandem borne by bacteriophage M13
sion of certain frameshift mutations in DNA polymerase I defi-inEscherichia coliK-12. Nucleic Acids Res.15:5323–5338.
cient strains ofEscherichia coli. Mol. Gen. Genet.141:251–262. Liu, B., N. C. Nicolaides, S. Markowitz, J. K. Willson, R. E. Par- Wagner, J., P. Gruz, S. R. Kim, M. Yamada, K. Matsuiet al., 1999
sonset al., 1995 Mismatch repair gene defects in sporadic colo- ThedinBgene encodes a novel E. coliDNA polymerase, DNA
rectal cancers with microsatellite instability. Nat. Genet.9:48–55. pol IV, involved in mutagenesis. Mol. Cell4:281–286.
McKenzie, G. J., P. L. Lee, M. J. Lombardo, P. J. HastingsandS. M. Wang, X., K. KitamuraandK. Yamamoto, 1996 Mutagenic speci-Rosenberg, 2001 SOS mutator DNA polymerase IV functions in ficity of N-methyl-N⬘-nitro-N-nitrosoguanidine in thetonBgene on
adaptive mutation and not adaptive amplification. Mol. Cell7: the chromosome ofEscherichia coli recA⫹andrecA⫺cells. Biochem.
571–579. Biophys. Res. Commun.227:334–339.
Mo, J. Y., andR. M. Schaaper, 1996 Fidelity and error specificity Yamamura, E., T. Nunoshiba, M. KawataandK. Yamamoto, 2000
of the alpha catalytic subunit ofEscherichia coliDNA polymerase Characterization of spontaneous mutation in theoxyRstrain of
III. J. Biol. Chem.271:18947–18953. Escherichia coli. Biochem. Biophys. Res. Commun.279:427–432.
Morel, P., C. Reverdy, B. Michel, S. D. EhrlichandE. Cassuto,