Indian J. Plant Physiol., Vol. XXVI, No.1, pp. 82-87 (March 1983)
EFFECT OF SHORT TERM STRESS ON FREE PROLINE
ACCUMULATION, RELATIVE WATER CONTENT AND
POTASSIUM CONTENT IN DIFFERENT PLANT PARTS OF
THREE COTTON GENOTYPES·
B. S. JANAGOUDAA, K. VENKATASUBBAIAH, K. V. JANAADHAN AND Y. C. PANCHAL Department of Crop Physiology,
Col ege of Agriculture, Dharwad (Received: August 30,1982)
SUMMARY
A pot culture experiment was carried out to study the effect of short term water stress on free proline accumulation, relative water content (RWC) and potassium (K+) content in different plant parts of three cotton genotypes, viz., Jaydhar. Bhagya and DCH-337. In all the genotypes stem showed higher accumulation of free proline under stress followed by leaf and root. Among the g3notypes, Bh3gya accumulated more proline in absolute quantities but with regard to number of times increase during stress, it was morl: in DCH· 337, indicating its tolerance to drought. Relative water content and potassium content differed among genotypes. Variety Jaydhar showed lesser reduction in RWe especially in stem and maintained high level of K+ even after alleviation of stress. Among the genotypes tested DCH-337 was found to be superior by maintaining higher proline even after alleviation of stress.
INTRODUCTION
Of late, attempts have been made to evaluate the genotypes of crop species for their drought tolerance based on their ability to accl:Imulate free proline as influenced by their relative water content (RWC) and potassium (K+) accumulation (Sinha and Rajagopal. 1975; Blum and Ebercon, 1976; Parameshwar, 1978; Viswanath, 1977). However, most of the workers have taken into account the free proline content in leaf only, with the assumption that its accumulation is more in leaf than in other plant parts. The proline accumulation in stem and root tissues of plant parts subjected to stress appears to be a result of translocation from the leaves (Singh et al., 1973). Thus, no detailed attempts have been made so far to look into the pattern of accumulation of proline under moisture stress in different plant parts. To elucidate this, a pot culture experiment was carried out with a short term water stress treatment.
MATERIALS AND METHODS
Three genotypes of cotton, viz., Jaydhar, Bhagya and DCH-337 were selected and grown in pots. Each set of three genotypes were grown in the same
..
STRESS ON COTTON GENOTYPES
83
pot to avoid large differences in soil moisture. The pots were filled with 15 kg of medium black soil. At the time of sowing, 2.0 g of urea, 3.0 g of single super phosphate and 0.75 g of muriate of potash were added to each pot. Three seedlings of each genotypes were maintained in each pot by thinning 20 days after sowing (DAS). When the seedlings were about 30 days old, moisture stress was imposed by withholding water supply and continued till ten days. Following this period, the plants were rewatered. At different time intervals after creating stress and also after alleviation of stress free proline, RWC and K+ content were estimated by following the methods of Bates et al. (1973) with due modifications and Barrs and Weatherly (1962), respectively. Proline was extracted and estimated from a final volume of 6 ml toluene. K+ content was estimated by following the method of Yoshida et al. (1972).
RESULTS AND DISCUSSION Free Proline
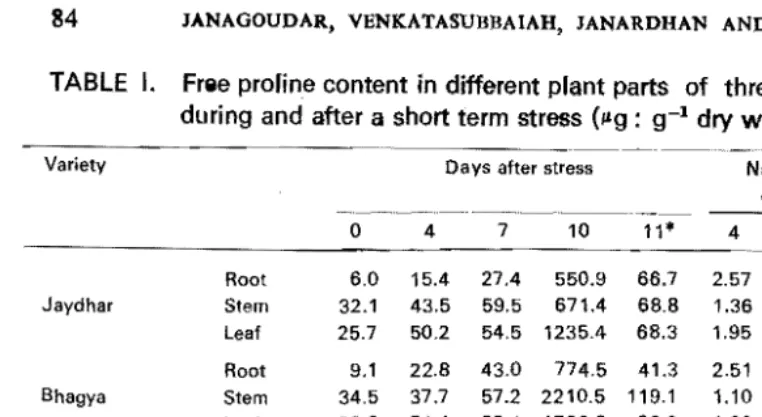
The data presented in Table I indicates genotypic differences in their ability to accumulate proline in different plant parts under common stress. The accumula tion of proline gradually increased with the increase in the period of stress reaching maximum with severe stress (ten days after withholding water). An abrupt and sharp decrease was observed at 24 h after rewatering. At early stage of stress (4 days after withholding water) high accumulation of proline was seen in leaves of DCH-337 both in absolute quantity (89.2 !kg) as well as increase in number of times (12.5) as compared to Bhagya and Jaydhar. In general, maximum amount was noticed in leaves followed by shoot and root in all genotypes. Under the conditions of mild stress (7 days after withholding water) also same trend was followed in all the genotypes on different plant parts. However, under severe stress, marked increase in proline was observed in stems of DCH-337 and Bhagya. Higher amount of proline was accumulated in Bhagya, whereas in terms of magnitude it was more in DCH-337 than in Bhagya and Jaydhar. Accumulation of proline in different plant parts when observed, Bhagya and DCH-337 accumulated maximum amount in stems, whereas Jaydhar accumulated more in leaves in terms of absolute quantities. Among the three genotypes, Bhagya accumulated more proline (4754 Ilg) under severe stress followed by DCH-337 (3550.2 !kg) and Jaydhar (2457.7 ,.g). From the results obtained, it is clear that DCH-337 had the ability to acCumulate consistently higher amounts of proline at all periods of stress and it was closely followed by Bhagya. However, in terms of absolute amount of increase, Bhagya was more responsive, but in terms of magnitude it was DCH-337. Marked decrease in proline content was observed at 24 h after alleviation of stress in all the genotypes. Thus, a reduction of 86.1, 91.7 and 95.3% was observed in DCH-337, Jaydhar and Bhagya, respectively. In all, DCH-337 was found to be superior over other genotypes by maintaining higher proline even after alleviation of stress (Table I).
Relative Water Content (RWC)
-84 JANAGOUDAR, VENKATASUBBAIAH, JANARDHAN AND PANCHAL
TABLE I. Free profine content in different pfant parts of three cotton genotypes during and after a short term stress (/lg : g-1 dry wt.)
- - - _ . _ - -..- . _.•
_
..._ . _ - - Variety Days after stress Number of times increaser
decrease oller zero day
o
4 7 10 11* 4 7 10 11*- - - _
.._ -.._.__
._---_
.• Root 6.0 15.4 27.4 550.9 66.7 2.57 4.57 91.98 87.9** Jaydhar Stem 32.1 43.5 59.5 671.4 68.8 1.36 1.85 20.90 89.8Leaf 25.7 50.2 54.5 1235.4 68.3 1.95 2.12 48.10 94.5
Root 9.1 22.8 43.0 774.5 41.3 2.51 4.78 85.10 94.7
Bhagya Stem 34.5 37.7 57.2 2210.5 119.1 1.10 1.66 64.10 94.6
•
Leaf 28.8 54.4 68.4 1769.0 62.3 1.89 2.38 61.40 96.5
Root 5.0 12.9 44.8 522.3 493 2.58 8.96 104.50 90.6 DCH-337 Stem 31.5 30.5 77.4 2494.9 331.1 -0.03 2.46 79.20 86.7 Leaf 7.1 89.2 94.9 553.0 114.2 12.56 13.37 77.90 79.3 Soil moisture (%) 24.5 17.0 12.1 8.9 26.2 1.98 293 59.90 91.5
*
After alleviation of stress.**Number of times decrease after alleviation of stress.
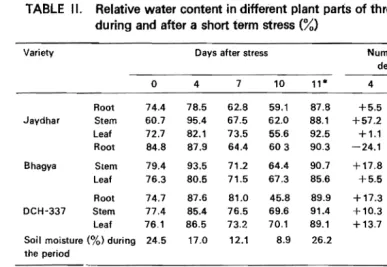
Under mild stress, the reduction in RWC was slight when compared with zero day, but it was marked when compared with RWC values under early stages of stress. Thus, the reduction was 2.0, 9.6 and 1.05% in Jaydhar, Bhagya and DCH-337, respectively as compared to zero day control. But it was 20.4, 21.0 and 11.1% in Jaydhar, Bh"gya and DCH-337, respectively compared to RWC at early stages of stress. Under severe stress the reduction in RWC was much more marked. In genera/' root showed more reduction followed by leaf and stem. Considerable increase in RWC was observed in all the genotypes when stress was alleviated after 24 h by rewatering. Genotypic differences showed that increase in RWC in Jaydhar, Bhagya and DCH-337, was to the extent of 23.1, 16.5 and 18.4%, respectively. Among different plant parts, increase was much more in shoots (24.3%) than in leaf (18.8%) and roots (14.5%). Higher increase in RWC was noticed in shoots of Jaydhar as compared to other genotypes at' ,,,.ell as other plant parts. In general, data indicated that genotypes differed in their ability to reduce RWC under a given commonstress. Jaydhar showed lesser reduction in RWC both under mild and severe stress, whereas Bhagya showed higher reduction. In DCH-337, marked reduction in RWC was seen only under severe stress. Thus, reduction in RWC did not increase proportionately with increase in stress in these genotypes. Further, within a genotype decrease in RWC increased free proline content but the genotypes differed in their capacity to accumulate proline at a comparable RWC values (Table 11).
Potassium Content (K")
STRE~ ON COTTON GENOTYPES 85
TABLE II. Relative water content in different plant parts of three cotton genotypes during and after a short term stress (%)
Variety Days after stress Number of times increase!
decrease over zero day
0 4 7 10 11* 4 7 10 11*
Jaydhar Root Stem Leaf Root 74.4 60.7 72.7 84.8 78.5 95.4 82.1 87.9 62.8 67.5 73.5 64.4 59.1 62.0 55.6 603 87.S 88.1 92.5 90.3 +5.5 +57.2 +1.1 -24.1 -15.6 + 11.2 +1.1 -28.9 -20.6 +2.1 -23.5 -28.9 +18.0 +45.1 +27.2 +65
..
Bhagya StemLeaf 79.4 76.3 93.5 80.5 71.2 71.5 64.4 67.3 90.7 85.6 +17.8 +5.5 -10.3 -6.3 -18.9 -11.8 +14.2 +12.3 DCH-337 Root Stem Leaf 74.7 77.4 76.1 87.6 85.4 86.5 81.0 76.5 73.2 45.8 69.6 70.1 89.9 91.4 89.1 +17.3 +10.3 +13.7 +8.4 +1.2 -3.8 -39.7 10.1 -7.9 +20.3 +18.1 +17.1 Soil moisture (%) during
the period
24.5 17.0 12.1 8.9 26.2
*After alleviation of stress.
TABLE III. Potassium content in different plant parts of three cotton genotypes during and after a short term stress (meq. g-1 dry wt.)
Variety Days after stress
0 4 7 10 11*
Root 0.80 0.73 0.98 0.66 1.28
Jaydhar Stem 1.67 1.23 1.44 1.11 1.36
Leaf 1.17 0.76 0.8~ 1.05 1.34
Mean 1.21 0.91 1.11 0.94 1.33
Root 0.89 0.64 0.72 0.68 0.90
Bhagya Stem 1.63 1.22 1.51 1.36 1.73
Leaf 0.77 0.88 0.81 1.15 0.77
Mean 1.10 0.91 1.01 1.06 1.13
"
DCH-337 Stem Root 0.66 1.22 0.86 1.17 0.75 1.03 0.71 1.11 0.58 0.91Leaf 0.84 0.63 0.89 0.87 0.95
Mean 0.91 0.89 0.89 0.90 0.81
Soil moisture (%) during the period 24.5 17.0 12.1 8.9 26.2
elleviiltion of stress.
observed under early stages of stress but in mild stress an increase was seen. Even under severe stress the increase in K+ content continued in Bhagya, whereas It decreased in Jaydhar. At all the stages of stress, DCH-337 maintained more or less similar K+ content as that of control. After 24 h of rewaiering. Bhagya and Jaydhar showed increase but not DCH-337 (Table III).
Therefore. it is clear that as against the general belief. leaves accumulate more free proline than other plant parts. stem accumulated more than any other
---~-~.
r
86
JANAGOUDAR, VENKATASUBBAIH, JANARDHAN AND PANCHALplant part (except in Jaydhar). However, in the initial stages of stress, proline accumulation was more in leaves, but as the stress period increased the accumu lation was more in stem. The results confirm those of Singh et al. (1973) who suggested that the proline accumulation in stem of droughted plants appears to be
a result~ of translocation from other parts. Also the results support those of
Thompson et al. (1966), who suggested proline accumulation under anaerobic conditions. According to him anaerobic conditions would reduce the rate of Kreb's cycle reaction and hence the accumulation of a-ketoglutarate, a substrate for synthesis of glutamic acid which in turn form the precursor for proline. Though stress may not cause anaerobic condition, the hypothesis that Kreb's cycle gets disturbed under stress with concurrent accumulation of a-ketoglutarate is quite appealing.
There appears to be no relationship between RWC and free proline content in any plant part studied and these observations confirm the reports of Mehkri et al.
(1976). For instance, though a negative relationship could be seen between RWC and free proline content in leaves, such a theory will not hold good when stem is taken into consideration. The stem which showed high RWC also showed high free proline accumulation. Hence, the results suggest that the water status of the tissue is not a reflection on the proline content in plant parts. Further, it can be said that proline is not sYnthesized de novo in stem, but accumulates due to trans location from other prant parts, probably leaves.
Though K+ accumulation under stress did not follow any particular trend among the cotton genotypes tested, higher values in Jaydhar followed by Bhagya and lower values in DCH-337 were noticed. These observations confirm those of Viswanath (1977), who reported differences in K+ content under stress in ragi genotypes. Potassium treatment has been shown to increase free proline in maize leaf discs (Mukherjee, 1974) and in cucumber cotyledons (Udayakumar et al.,
1976).
Thus, it could be inferred that genotypes Jaydhar and Bhagya which accumulate higher K+ content could also accumulate higher amount of proline. Shashidhar et al. (1981) also showed similar trend of K+ and proline accumulation in groundnut genotype RS 218. However, when plants were rewatered, K+ content increased in Jaydhar and Bhagya followed by a sharp decrease in proline content. The significance of this relationship is not well understood at this stage. However, it may be pertinent to add here that K+ under moisture stress appears to have a role as an osmoregulent. Thus, it is worthwhile investigating further the
r~lationship between K+ accumulation and drought tolerance. . REFERENCES
Barrs, H. D. and Weatherly, P. E. (1962). A re-examination of the relative turgidity for estimating water deficits in leaves. Aust. J. BioI. Sci., 15: 413-28.
Bates, L S., Waldren, R. P, and Teare: I. D. (1973).Rapid detl)iminati'on offree proline in water stress . studies. Plant and Soil, 39: 205-08.
r
STRESS ON COTTON GENOTYPES
87
Blum. A. and Ebercon. A. (1976). Genotypic responses in sorghum to drought stress. III. Free proline accumulation and drought resistance. Crop Sci.• 16: 428-31.
Mehkri, A. A., Shashidhar, V. R" Udayakumar. N. and Krishnasastry. K. S. (1976). Screening of varieties for relative drought tolerance in groundnut. Indian J. Plant PhYSiol., 119: 108-11.
Mukherjee. I. (1974). Effect of potassium on proline accumulation in maize during wilting. Physiol. Plant, 31 : 288.
Parameshwar, G. (19(8). Screening varieties for relative drought tolerance in sorghum. M. Sc. (Agri.) thesis, Univ. Agric. Sci., Bangalore.
Shashidhar, V. R., Mekhri, A. A. and Krishnasastry, K. S. (1981). Potassium content and proline accumu lation following seed treatment with calcium chloride in groundnut varieties. Indian J. Plant Physiol., 24 ; 89·92.
Singh, T. N., Paleg, L G. and Aspinall, D. (1973). Stress metabolism. III. Variations in response to water deficit in barley plant. Aust. J. BioI. Sci.. 26 ; 65-76.
Singh, S. K. and Rajagopal. V. (1975). Effect of moisture stress on nitrate reductase activity and accumu' lation of proline in sorghum. Proc. Symposium on "Crop plant response to environmental stresses". I.A.R.I., New Delhi.
Thompson, J. F., Stewart, T. R. and Morris, C J. (1966). Changes in amino acids content of excised leaves during incubation. I. The effect of water content of leaves and atmospheric oxygen level.
Plant Physiol., 41 : 1578-84.
Udayakumar, M., Ramarao, S •• Prasad. T. G. and Krishnasastry, K. S. (1976). Effect of potassium on proline accumulation in cucumber cotyledons. New Phytol., 77 : 593-98.
Viswllnath, H. R. (1977). Evaluation of ragi genotypes for relative drought tolerance based on physiologi· cal parameters and field performance. M. Sc. (Agri.) thesis, Univ. Agric. Sci., Ban9alore. Yoshida, S. D., Forno, J. R, Cook and Gomez, K. H. (1972). Laboratory manual for physiological studies
of rice. I.R.R.I. Second ed., Los Bonos, Philippines .