r'
,)6 ..
Q " JniliQIJ J. Plant Physiol., Vol. XXX, No.4, pp. 41().4l4. 1987SHORT COMMUNICATION
EFFECT OF ABA AND KINETIN ON FAT AND PROTEIN DEGRADATION DURl:NGIMBIBITION OF DORMANT AND
NON-DORMANT GROUNDNUT SEEDS
ARUNA SHARMA, B.K. GHOSH AND U.K. SENGUPTA
Division of Plant Physiology, Indian Agricultural Research Institute, New Delhi-110 012
Total fat content of fresh non-dormant seeds of cv. J-II decreased with time during imbibition of water while little change was observed in fresb seeds of dormant cV. M-J3. A period of dry storage overcame dormancy in M-13 and Increased tbe rate of fat depletion. Treatment with A~ decreased, wbile that with kinetin increased the rate of fat hydrolysis. ABA was more effective in inbibiting protein hydrolysis rather than fat breakdown in germinating seeds while kinetin treatment enhanced fat hydrolysis much more than protein breakdown in dormant seeds. Seed storage, enbanced the fat hydrolysis even in non-dormant cultivar. Treatment with ABA. caused effects similar to those seen in dormant seeds indicating that dormancy may be
due to like inhibitor ABA.
Freshly harvested seeds may not germinate, when placed under apparent {)ptimal conditions of temperature, moisture and oxygen. The mechanism preventing germination is not fully understood but a period of dry storage or after Tipening (Khan, 1977) is able to overcome the block to the germinating process. Dormancy has been attributed to several causes (Amen. 1968; Black. 1972;
Villers, 1975). One of the causes is the relative concentration of certain growth regulating substances, which influences tbe metabolism of the seeds (Khan et. al.
1971; Ketering, 1973). Germination in non-dormant seeds occurs because of the favourable balance of growth hormones. Dormant seeds, however, do not 8erminate unless either some germinatmg agents are supplied exogenously or they are produced endogenously in response to environmental triggers (Jan and Amen,
~r"
r . .
FAT AND PROTBfN DEGRADATION BY ABA AND KIl'lBfll'l IN GROIJNDNlJf SBEDS 421
in fat and protein br"akdowa dLlring illlbibition of dormant and non·dormJ.nt goundnut seeds and how these are affected when dormancy is broken by kinetin -or imposed by ABA.
Seeds of two groundnut cultivars, J-Il and M-13 were collected from fields -of Indian Agricultural Research Institute, New Delhi. The seeds of cv. M-l3 remain .dormant upto 60-70 days after harvest under Delhi conditions while cv. J-ll is a Don-dormant cultivars (Sengupta et. 01. 1979). Seeds were used immediately after harvest and some were kept under dry storage for 90 days to eliminate dormancy.
Healthy and uniform seeds of both cultivars at two different stages 'of storage were soaked in solutions of ABA (2.5
x
1O-4M) and kinetin (2.5 X 10-4M)for 6 hours. Seeds were then transferred to petriplates (20 seeds/plate in three replicates) over moistended germinating paper in the same solution and allowed to germinate in BOD incubator at 30"C in dark. Distilled water was used as control. Fat and amino acid content was estimated only in cotyledons (seed coat and embryo removed) every 24 hours. Total fat content was estimated by cold percolation gravimetric method (Kartha and Sethi, 1957). 500 mg sample was ground and .filled in (30 em) glass column with layers of sodium sulphate. The fat was eluted with petroleum ether (BP 70-90°C) and expressed at per cent fresh weight of seeds. Total free amino acid (TFAA) was estimated after defatting the cotyledon with petroleum ether. The TFAA were extracted in ethanol and estimated by the method of Lee and Takahashi (1966).
It has been reported earlier that seed dormancy of groundnut cultivar M-13,
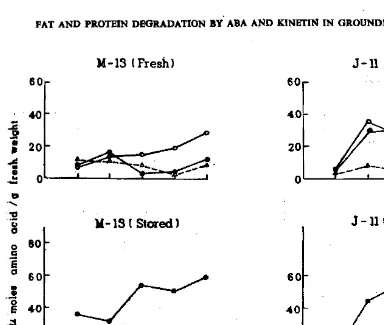
-can be broken by kinetin application (Sengupta et. 01. 1979). In the present study, the fat depletion rate of stored seeds of M-13, which had lost their -dormancy was much higher than that in fresb. seeds which were dormant (Fig. 1).
During first five days of imbibition, the depletion of the fat was 28% as against .85
%
in stored seeds. It was observed that even though no germination seen was in fresh dormant seeds of M-13 yet there was some fat hydrolysis. Fresh r seeds ofJ-ll which were non-dormant, showed comparatively slower rate of fat hydrolysis {46%) than the stored seeds (83%). It appears that some kind of inhibitors like those present in dormant M-l3 seeds-may be present in fresh seeds of J-Il also. Seed storage could reduce the inhibitor content or increase the content of germina 1ing promoting substances resulting in increased fat hydrolysis via the enzyme -System. Further work is needed to support this assumption It has also been seen
~arlier that though the fresh seeds of J-Il do germinate well yet the growth rate of
~mbryo is slower than that seen in case of stored seeds (Sengupta etc aI. 1971). The inhibitor reported, to be present in groundnut sc:eds is ABA (Ketering, 1973;
~arasim1lareddy and Swamy, 1979).
422 ARUNA SHARMA BY AI..
1(':'13 (Fresh) J -11 (Fresh)
10
80
... "'"
60 60
.
40 r 40
I::
.!
i 20 20
...-__A--_
c;
1
0 0
~! 100 )4-13 (Stored) 100
J
-11 (Stored)j
80 10..
60
I.
' 60
40 40
...,..,A
Ar-""""'~--A_-_A'
20 20
0 0 6 120
24 48 72 96 120
HoW'S Glter 8ookmo
Fig.. 1 : Meet
or
ABA and Kinetin on fat degra.dation during imbibitionor
dormant and non-dormant groundnut seeds.<. •
Control, 0---0 Kinetin and /:1-/:1ABA). The pattern of protein hydrolysis. as seen . from the content of free aminoacids,
wassimilar
to thatseen
for fat hydrolysis in both cultivars (Fig. 2). Stored seeds of M-13 showed higher total free amino acids than the fresh ones while fresh J-ll seeds which are non-dormant also showed slowed slower rate of amino acid accumulation than the stored seeds. Treatment with ABA inhibited the germina tion of stored M-13as
well as fresh and stored J-Il seeds~ Fresh M·13 ,seeds did not germinate any way due to being dormant.ABA
treatment also reduced fat hydrolysis and protein bnatdown in both fresh as well as,stored seeds of both cultivars. It was found that ABA· treatment inhibited fat hydrolysis to a much lesser extent as·, compared to, protein breakdown in germinating seeds of both c~vars.y . , •
Thus it apPears that ABA was more inhibitory to the enzymes involved in , protein break down' than those involved in fat hydrolysis. KiJietin on the other
hand caused enhanced fat depletion and amino acid aecumUlationin'dotmanl
seeds
FAT AND PllOTElN DIlGllADATION BY ABA AND KINBTlN IN GllOUNDNlIT SEEDS 423
M-13 (Fresh) J-U (Fresh)
,
60 60
.'
4.0 40
-i
..
20 20
•
1
..
.0-
..
0 ...:g
J -
U (Stored)11-13 ( Stored J
8
80 .5 0
&
CI 60 60
.J
2
40 404
20 . 20
•
...-A...~6 - - - ... _ - & - . - - 6 . - _ _A
Y"
--A-_
O· 0
...
3t 48 ?286 120 24 ''48 12 86 120
Houn alter soatlDg
Fig 2: Effect of ABA and kinetin on amino acid content during imbibition of dormant and non-dormant groundnut seeds. ( . • Control. 0--0 Kinetin and b..-b..ABA).
-of M-13. The rate of fat depletion however was enhanced much more than that -of amino acid accumulation. Kinetin was ineffective in increasing free amino acid -contentJn fresh non-dormant seeds of J-II.
424
p4
U-r--·!
ARUNA SHARMA ET AL.
REFERENCES
Amen, R.D. (1968). A model of seed dormancy. Bot. Rev., 34: 1.
Black, M. (1972). Control processes in germination and dormancy. Oxford Biology Readers. 20: r Jan, R. C. and Amen, R.D. (1977) What is germination. In Physiology and Biochemistry of Seed
Dormancy and Germination. Elsevier/North Holland Biomedical Press. 8.
K artha, A.R.S. and Sethi, A.S. (1957). A cold percolation method fer rapid gravimetric estimatiOD of oil in small quantities. Indian J. Agr:ic. Sci., 27 : 211.
Ketering. D.L. (1973). Germinatior; inhibitors. Seed Sci. & Technol. 1 : 305.
Kban, A.A.. Heith, C.E., Waters, E.C., Anojl'lu, c.c. and Anderson. L. (1971). Discovary of new role of cytokinin in seed dormancy and germination. Search Agric. 1 : 1
Khan, A.A., (1977). Seed dom-ancy.: Changing concepts and theories. In The Physiology and Biochemistry of Seed Dormancy and Germination. Elsevier/North Holland Bio~edical
Press, 29.
Lee, Y.P. and Takahashi, T. (1966). An improved colorimetric determination efamino acids with the use the ninhydrin. Analyt. Biochem., 14 : 71.
Narasimhareddy,S.B. and Swamy, P.M. (1979). Abscisic acid like inhibition and cytokinins in. the developing seeds of dormant and non·dormant varieties of peanut (Arachis. hypogea L).
Sengupta, U.K., Sirehi, G.S., Kairr>, M.R.S. and Pokhryal, T.e. (1977). Effect of growth regulators on seed germination of groundnut. Indian
.
J. Plant Physiology. 20 : 91.4.,
Sengupta, U.K., Pandey, M. and Sirohi, G.S. (1979). Effect of growth regulators on seed germination in groundnut. : Interaction studies. Seed Ires., 7: 114.