EFFECT OF PHOTOPERIOD ON THE DEVELOPMENT OF SHOOT APEX OF PANlCUM MlLlACEUM L •
. ...~
M. S. TAYAL AND K.K. NANDA·
D.A.V. College, Muza§arnagar (U:P.)
Received on May 3, 1978
SUMMAIlY
This paper deals with the effect of photoperiod on the developmental changes in the shoot apex of Panicum mil;aceum during vegetative (formative), gametogenetic and embryogenetic phases. While the periods from sowing to panicle initiation and from panicle initiation to gametogenesis were the shortest under SD and longest under LD conditions, the period from the completion of gametogenesis to embryogenesis did not differ under the three photoperiods. The size of the inflorescence and the number of spikelois were also considerably reduced under SD conditions, although the sizeof . floral organs and the embryo did not differ much with -the photoperiod. It is con
cluded that thwhortening of vegetative period of this plant sown in September as compared to tnat sown earlier as reported in the previous communication is primarily due to the shorter days that prevail during the growing period of that sowing.
INTItODUCl10N
Tayal and Nanda (1917) have shown that in Panicum miliaceum an alteration in the environmental complex with the change in time of sowing has a· marked' effect on the rate with which events proceed in the formative. gametogenetic and embryo genetic phases. It was suggested there that those effects were probably due to a change in the day-length and temperature conditions which prevail during the growing period of plants of different sowings. This investigation was undertaken to elucidate this point and study the effect of photoperiod on the developmental events during different phases in the ontogeny of this plant.
MATI!RIALS AND METHODS
Puro line seeds of Panicum miliaceum var. Samai Co. I, were sown in 30 unglazed earthen pots (30 cm dia.) containing a 3 : 1 mixture
ot
garden soil and . farmayard manure. The pots were divided into 3 groups of 10 each. One group was exposed to long days (LD) with 18 hours of illumination, the other group to normaldays (ND) obtaining in nature at Muzatrarnagar from May to October, and the third one to short days (SD) consisting of 8 hours of sunlight alternating with 16
hours of dark.
The growing apices were collected first at weekly intervals but later on twice a week and serial microtome sections were prepared (for details see Tayal and Nanda,
1977).
The mean periods (in days) from sowing to panicle initiation, gamete formation and embryo formation were determined from the sections of shoot apices. These have been designed as 'period of panicle initiation', 'period of gametogenesis and 'period of embryogenesis, respectively. The periods for the initiation of different floral organs, micro and mega~sporogenesis and gametogenesis from the date of panicle initiation and for embryogenesis from the date of completion of gametogenesis. were also determined from these sections. The total number of spikelets formed on the main shoots of five tagged plants, was also determined.
REsuLTS
Vegetative phase : -The various changes in the growing apex till its transforma tion to the reproductive state are presented in Figs. 1-4, 12-15 and 21-24 for ND,
SD and LD plants respectively. The size of the shoot apex at successive develop mental stages is given in table I.
Under ND condition, the shoot apex is broader than long at l . y s (Fig. 1, Table I) but elongates slightly at 32 days (Fig. 2). At this stage a single layered tunica surrounding the homogeneous parenchymatous corpus can be seen in the meristem (Fig. 2 i). At subsequent samples, the shoot apex gradually increases in size (Table I), the increase being more in length so that its length and diameter be come more or less equal at 88 days (Fig. 3. Table I). This elongation probably marks
the initiation of the change of the shoot apex from vegetative to the reproductive ~
state. shows
The shoot apex increases both in length and diameter by 102 days and
a few small projections above the leaf primordia (Fig. 4, Table I). These
I
! projections are the primordia of the branches of panicle and appear to markthe first visible morphological change in the transition of the growing apex from
the vegetative to the reproductive state. The normal growth of leaf primordia below
1
ithese projections continues. The increase in size of the shoot apex during this· stage 1
is due to the increase in the number of cells and not due to increase in the size of cells (Tayal et al., of 1961).
The sequence of changes in the growing apex of SD plants (Figs. 12-15) is
I
EFFECr OP PHOTOPEllIOD ON SHOOT APEX OP PANICUY MILIACEUM 3
4 M.S. TAYAL AND 1t.1t. NANDA
,i
""
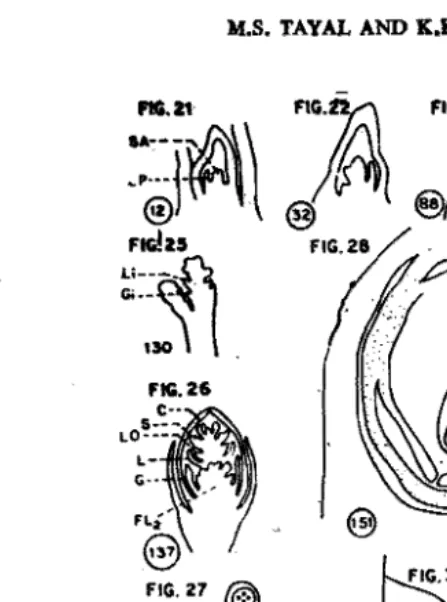
Fig. 1-30. Semidiagrammatic median longitudinal sections of the growing apex at different stages of development of Panicum miliaceum under different photoperiods.
Figs. 1-11=ND, Figs. 12-20=SD, Figs. 21-30=LD. SA=Shoot Apex, Lr>=Leaf Primodium, Gi=Glume inifial. SP=Spikelet, Li=Lemma initial, GL=Glume, FLt=Flower I. FLt=F1ower 2, FL3=Flower 3, Ci=CarpeI initial, Si=Stamen initial.
C=CarpeI, S=Stamen, Lo=Lodicule, P=Palea, L=Lemma, RP-Rachis Primordia, E=Embryo, En=Endosperm.
(Magnification in all Figs. X 60 except Figs, 2i. 9i, 10, 18, 19i, 28 & 29 x 210).
state after 7 days as is evident from the prominent protrusions representing panicle branches (Fig. 15). The vegetative phase is, thus completed within about 53 days in SD plants as compared with 102 days in ND ones.
EFFECf OF PHOTOPliRIOD ON SHOOT APEX OF PANICUM MILIACEUM 5
Table 1. Length and diameter (in p.) of the shoot apex during different stages of development of panicum miliaceum.
TiJne..Days
from sowing SD
Photoperiodic Treatment
No lJ)
1 2 3 4
12 2S
32
40 53 fO 88 102
116
SO+66 75+83
15+15 374+166
42+75
58+66 66+83
83+100 100+105 266+186
-33+58
42+70 40+12
50+75 59+75
158+133
21). The shoot apex at this stage is smaller than that at a similar stage of NO and SO plants. The growing apex increases very slowly in size till 88 days (Figs. 22, 23, Table I) but elongates appreciably at 130 days when the primordia of panicle branches are also initiated (Fig. 24). The:first visible morphological change in the transition of the growing apex from vegetative to reproductive state, therefore, is observed after 116 days in these plants as compared with 102 days in NO and 53 days in SO ones. The initiation of panicle bninches results from the anticlinal division of both tunica and corpus cells (Fig. 24). The cell size does not change much during development
•
till the initiation of panicle.An idea about the rate with which developmental process proceeds upder the three photoperiods can be had from table I. The increase in size of the growing apex-both in length and diameter is very slow in LO plants. It is more rapid in NO ones and most rapid in SO plants. Thus, SO plants attain a size of :: 74
x
066 1.1. with in :-3 days. In ND and LO plants this size of the growing apex is not reached even after 102 and 116 days, respectively.Gametogenetic phase; This phase relates to the developmental changes from panicle initiation to gamete formation. These are shown in Figs. 5-9, 16·18 and 25-29 in NO, SD and LO plants, respectively.
M.S. TAYAL AND K.1t. NANnA
are clearly seen. In the axil of each lemma, a flower can also be seen. The initiation of palea. lodicules, stamens and carpels has also taken place in the uppennost flower. The development of the spikelet is completed within 1 days when a distinct ovule in the ovary and pollen mother ceUs in the stamen are formed in the uppermost flower (Fig. 8). The tlower next below it, exhibits a stage of development less advanced than that of the topmost one. The poUen grains are
seen
in the anther lobe . ~t~139 days. The carpel has also ditl'erentiated into ovary, style and stigma by this time (Fig. 9). An 8-nucleate embryo sac within the ovule can also be clearly made out.In SD plants the inflorescence axis elongates appreciably at 60 days and exhibits spikelet formation (Fig. 16). The well-developed glumes and three lemmas can be seen in the uppennost spikelet. The initiation of a tlower in the axil of each lemma has also started. Even the palea can be observed in the uppermost tlower (Fig. 16)•... A similar stage in ND plants is reached in 125 days (Fig. 1). The stamens become well developed in the uppermost tlower and the pollen mother cells are formed at 61 days (Fig. 11). A single ovule within the ovary can be seen. The lodicules have also developed by this time. A second flower of the spikelet can also be observed. It is smaller in size and is at a much earlier stage of development.1"he third tlower is not visible probably because it is suppressed, The stamens and carpel of the uppermost flower mature in 74 days when the pollen grains are seen within the
stamens and an embryo sac within the ovary (Fig. 18).
The changes in the growing apex in LD plants (Figs. 25-29) are similar to those in ND ones (Figs. 5-9). Thus, a well developed glume and lemma can be seen in· the uppermost spikelet of panicle after 130 days (Fig. 25). The flowers in the axils of lemmas have also been initiated. The glumes cover the tloral parts completely' and the lemmas increase in size after 131 days (Fig. 26). The palea and lodicules are also formed. The stamens are elongated and exhibit archesporial stage and the carpel in the topmost flower has a single ovule. Fig. 27 shows the uppennost spikelet after 144 days. The floral parts are all fully developed by this time. The pollen grains within the anther lobes and 2-nucleate embryo sac within the ovule can be clearly observed. As in ND and SD plants, the second flower in the spike lets is not' visible and is probably suppressed. The ovary of the uppermost spikelet shows 'a fully mature embryo-sac after 151 days (Fig. 28). The megagametog~esis is, thus, completed in 35 days in LD plants. In ND plants also this phase is completed in
35 days. In SD plants, on the other hand, it takes longer to coinplete this ph~. The micro-gametogenesis is oompleted artera week and the 2 nuclei in the pollen grains-a generative and tube can be observed (Fig. 29).
Embryogenetic phase : -This phase relates to ontogenic changes occurring from . ' .
EJlFECr OF PHOTOPERIOD oN sHoot ApEX OF pANlcuM MiLtACBUM
if
suspensor is observed 7 days after gametogenesiS in ND and SD plants (Figs. 10 and 19). The embryogenesis is completed in 14 days after the completion of the develop • ment of embryo sac under aU the photoperiods (Figs. 11, 20 and 30 for ND, SD andLD, respectively). The structure of the embryo is typically monocotyledonous. Size of the inflorescence andfloral parts :-Table II shows that the inflorescence of ND plants in the longest and of SD ones the shortest (Table II, column 2). The glumes are more or less of the 'same size in SD and ND plants while those of LD
.
.
plants are smaller (Table II, columns 3 and 4). The lemma and palea lengths do not differ significantly with the photoperiod (Table D, columns 5 and 6). The lodicule, stamen and carpel of LD plants are longer than those of SD and ND ones. The differencess in the latter two are not statistically significant (Table. II. columns 7-9).
Although, the size of floral parts differs under the three photoperiods, that of the embryos does not show significant differences.
Table II. Effect of photoperiod on the length of inflorescence and floral organs
and number of spikeletslear in Panicum miliaceum
Photoperiodic Length (cm) of Number
treatment ~ores- Glume Glume Lemma Palea Lodi- Stamen Carpel Embryo of Spike
renee I II cule ______~___ 1,,~/ear
2 3 4 S 6 7 8 9 10 11
SD 20.3 0.39 0.37 0.30 0.28 0.01 0.30 0.30 0.14 166.4
ND 37.7 0.39 0.37 0.31 0.29 0.05 0.27 0.30 0.15 540.8
LD 32,54 0,36 0.34 0.31 0.28 0.10 0.34 0.34 0.15 461.8
Number of spikelets per ear : -The photoperiod also influences the number of spikelets produced per ear (Table n, column 11). Fewer spikelets are produced in LD and SD than in ND plants, the decrease being more in SO than in LD plants.
CO> Effect of photoperiod on the period elapsing between various ontogenic eve1lts in the development of Table lIf. growing apex and floral axis in Panicum miliaceum
Photo- periodic treat ment Period of
.
panicle, initiation
PerIod of of game togenesis Period of embryo- genesis Glume Period (Days) from panicle initiation to the formation of Lemma Palea Stamen carpel' Micro Oameto---am:bryoge and spore genesis nesis lodicule
Period from gameto
::c
(Days) (Days) (Days) Mega Micro genesis toembryo- genesis (Days) ~
;!
i< [!!;SO ND LD 2 53
102
.
116
3 7S 173 151
4
89 151 16S 5 7 7 7
6 7
14 10 7 8
21 14
8
10 21 18 9 10 .
21 18 10 22 35 28
11 22 3S 35
12 29 3S 42 13 3S 49 49 14 13 14 14
~.
\111,.. .
:Z:'>
~,>
F·SPiJii ~#~(j!-.... • ... '" fl ••
1iIIW-,,j;
_
IIo..-+~' . -, ...'_h
,-' " , .•. JI' ... # .... M t .. ___I
.._,...,.
r . -
. .
,
,
JlIEECT oP PHOTopmUol> ON SHoot APEX OF PANICUM MtLIACEUM 9
r
I DISCUSSION
l
110
The results presented in this paper clearly demonstrate that while photoperiod ttas a pronounced effect on the formative and gametogenetic phases, it hardly affects the rate of events during the embryogenetic phase. The period from the completion of gametogenesis to embryogenesis, thus, did not change with the photoperiod. The shortening of the length of vegetative period under SD conditions is .. therefore, due to the snortening of' the formative and gametogenetic phases. The decrease in the longth of the vegetative period in September sowing as compared with that in earlier sowings reported in a previous communication (Tayal and Nanda, 1977) may, there fore be ascribed to the shorter days that prevailed during the growth of plants of that sowing.
Another interesting point that emerges from this investigation is that the photo period affects not only the rate with which events proceed during different ontogenetic phases but also the magnitude of these events. Thus, in plants exposed to short day conditions the floral axis was short with fewer protrusions of panicle branch initials and spikelets. The decrease in the size of panicle and the number of spikelets takes place in spite of the opitmum level of nutrition in the soil, thus, confirming the view put forth earlier that the rates of uptake of mineral nutrients are governed by the same regula\ory. mechanism which determines the developmental rythm of the plant (Chinoy ~na Nanda, 1950, 1952, Sirohi et al., 1959). This is supported by the fact that similar hastening of ontogeny and reduction in the size of the inflorescence and number ofspikelets in long day plants, wheat (Garg, 1%0) and Oat (Tayal and Nanda, 1962) are brought about when they are exposed to long day conditions.
R.:EFim.ENCES
C'l!noy, I.J. and K.. K.. Nlnd:t.. 1950). Photop~riodic treltmant and nitrogen uptake in wheat.
cu".
Sci•• 19 : 24-25.- - _...._ -•• and • (1952). Effect of vernalization and photoperjodic treatment on
growth and development of crop plants. IV. Uptake of nitrogen, phosphorus· and potassium by wheat under varying photoinductive and post photOinductive treatments.
Phys/o/. Plant., 5: 11·32.
Garg, O.P. (1960). A morphological study of the growing apex with special reference to its influence upon growth and development of some crop. plants. Ph. D. thesis, Delhi University. Sirobi, G.S., K..L. Sawhney. J. J. Chinoy and K.K. Nanda. (1959). Growth and phasic development
' o f wheat. HI. Dry rna tter production and water content. Indian
J:
PI. PhYsio/••' 1. : 83. Tayal. M.S. M.P. Kaushik and ,KK.. Nanda. (1961). Growth and phasic development of oat I. A :,developmental study of the shoot apex in two varieties of Avena sativa under differential
photoperiodic treatments. Indian J. PI. Physial. 4 : 13249. .
- - : " - -•• and K.K Nanda (1962). Growth and phasic development of oat
n.
Development" of the inflorescence and 80m! organs in two varities under two photoperiods. Phylan., 19 : . 14]-153.- - - " - . ' J and . (1971), Etf(lCt of time of sowing on the development of shoot