T he biochem ical and genetical analysis o f lactase phlorizin hydrolase:
w ith specific reference to the lactase persistence/ non-persistence
polym orphism in man.
C lare B arbara H arvey
A thesis subm itted for the degree o f D octor o f P hilosophy
in the U niversity of London
June 1994
M R C H um an B iochem ical G enetics U nit,
Gallon L aboratory
ProQuest Number: 10106853
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10106853
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author. All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
ABSTRACT
This thesis describes investigations on the biochemical and genetical properties
of human intestinal lactase-phlorizin hydrolase which were designed to help in the
understanding of the molecular basis of the lactase persistence / non-persistence
polymorphism in man. There were two major objectives, one was to study the
expression of the lactase gene in lactase persistent and non-persistent individuals and
the second was to identify as many polymorphisms as possible within the lactase gene
in order to increase informativeness for linkage analysis and to explore the extent of
allelic association across the gene.
The expression of the lactase gene was studied in a series of samples of adult
intestine. The level of the enzyme activity, the protein and the mRNA was examined in
each sample. Individuals were assigned lactase persistence status on the basis of their
lactase to sucrase activity ratio. The results of this analysis suggest that differences in
the level of transcription of the lactase gene are important in determining the lactase
persistence phenotype. Lymphoblasioid cell lines were established from 32 of these
individuals and used to prepare genomic DNA.
The mapping of the lactase gene to chromosome 2 was confirmed and refined to
band q21 using a panel of somatic cell hybrids and in situ hybridisation.
Using a variety of electrophoretic techniques which are sensitive to the detection
of single base changes, polymorphisms were identified at seven different sites within
the 70kb region comprising the lactase gene. Analysis of these polymorphisms in 50
families revealed that only 3 of the possible 128 haplotypes occur frequently,
suggesting an area of linkage disequilibrium stretching across the whole lactase coding
region. These markers have also been used for linkage analysis and enabled the
The lactase gene polymorphisms are currently being used to study unrelated
individuals as well as families charactensed with respect to their lactase persistence /
Ta b l e o f c o n t e n t s
p a ge number
Abstract. 2
Table of contents 4
List of figures. 10
List of tables. 13
Abbreviations. 15
Acknowledgements. 16
1. IN T R O D U C T IO N . 17
1.1 Ex p r e s s i o n a n d c h a r a c t e r i s t i c s o f t h e e n z y m e
LACTASE PHLORIZIN HYDROLASE. 18
1 .1 .1 CHARACTERISUCS o f t h e ENZYME. 18
1.1.2 E v i d e n c e f o r d i m é r i s a t i o n o f t h e l a c t a s e p r o t e i n. 19
1.1.3 GLYCOSYLATION o f THE LACTASE POLYPEPTIDE. 19
1.1.4. O t h e r INTESTINAL DISACCHARIDASES. 20
1.1.4.1 S u c r a s e - is o m a lta s e . 20
1.1.4.2 M a lta se-g liic o a n iy la se. 21
1.1.4.3 T reh alase. 21
1 .1 .5 VARIATION IN ENZYME ACTIVITIES WITHIN THE SMALL INTESTINE.24
1.1.5.1 Variation in lactase and other enzyme activities within
a villus. 24
1.1.5.2 Variation in lactase and other enzyme activities within
the small intestine in humans. 24
1.1.5.3 Variation in LPH activity within the small intestine in
other animals. 25
1.1.6 D e v e l o p m e n t a l r e c i t a t i o n o f t h e LPH p r o t e i n i n m a n. 26
1.1.6.1 Lactose tolerance tests. 27
1.1.7 Ev id e n c e f o r m e n d e l i a n i n h e r i t a n c e. 32
1.1.7.1 Evidence from family studies. 32
1.1.7.2 Evidence from twin studies. 33
1.1.7.3 Evidence from population studies. 34
1.1.8 Ch a r a c t e r i s t i c s o f t h e l a c t a s e m e s s e n g e r RNA a n d
THE r e s u l t i n g PROTEIN. 38
1.1.8.1 Molecular characteristics o f the protein deduced from
the cDNA sequence. 38
1.1.8.2 Characteristics and tissue specificity o f the
1.1.8.3 Quantification of the levels o f lactase messenger RNA and determination of its cellular localisation compared with the localisation o f lactase protein and activity. 41
1.1.8.3.1 Studies using rabbit intestine. 41
1.1.8.3.2 Studies using rat intestine. 43
1.1.8.3.3 Studies using human intestine. 46
1.1.9 Lo c a l i s a t i o n AND ORGANISATION OF THE LACTASE GENE. 5 0
1.2 E x t e n t o f g e n e t i c v a r i a t i o n i n m a n . 5 2
1 .2 .1 INBORN ERRORS OF METABOLISM. 5 2
1 .2 .2 COMMON ENZYME POLYMORPHISMS. 5 2
1 .2 .3 DNA BASIS FOR PROTEIN VARIATION. 53
1 .2 .4 . Ot h e r o c c u r r e n c e s o f a bn o r m a l p r o d u c t i o n o f a
TEMPORALLY REGULATED PRODUCT. 58
1.2.4.1 Persistence o f fetal haem oglobin. 58
1.2.4.2 Persistence o f fetal alpha-fetoprotein. 59
1.3 DETECTION OF D N A P O L Y M O R P H I S M S . 60
1 .3 .1 D e t e c t i o n m e t iio d s . 61
1 .3 .2 Si n g l e St r a n d Co n f o r m a t i o n An a l y s i s (S S C A ). 63
1 .3 .3 DENATURING GRADIENT GEL ELECTROPHORESIS. 67
1 .4 L IN K A G E STUDIES AND HAPLOTYPE ANALYSIS. 6 9
1.5 A IM S OF THIS PRO.IECT. 7 2
2. M ATERIALS AND METHODS. 73
2.1 An a l y s i s o f i n t e s t i n a l s a m p l e s. 73
2.1.1 Du o d e n a l b i o p s ie s e r o m h u m a n s. 73
2.1.2 INTESTINAI_ SAMPLES FROM ANIMALS. 74
2.1.3 Pr e p a r a t i o n OF SAMPLES. 74
2.1.3.1 Preparation of aqueous and Triton extracts. 74
2.1.3.2 Preparation o f hoinogenates. 75
2 . 1 . 4 IMMUNOHISTOLOGY. 75
2.1.5 De t e r m i n a t i o n OF LACTASE AND SUCRASE ACTIVITIES. 76
2.1.5.1 Routine protocol. 76
2 .1.5.2 M ethods used to evaluate assay protocol. 76
2 . 1 .6 SODIUM DODECYL SULPHATE POLYACRYLAMIDE GEL
ELECTROPHORESIS (SDS-PAGE). 77
2.1.6.1 Preparation o f SDS-PAGE gels. 77
2 .1.6.2 E lectroph oresis of SDS-PAG E gels. 78
2.1.6.4 Immunodetection o f specific proteins. 79
2.1.6.4.1 Estimation of apparent molecular weight. 79
2.2 ESTABLISHM ENT AND MAINTENANCE OF
L Y M P H O B LA SIO ID CELL LINES. 80
2.3 Ge n e r a l DNA m e t h o d s. 81
2.3.1 COMMONLY USED BUFFERS. 81
2.3.2 AGAROSE GEL ELEC TR O PIlO l^SIS. 81
2.3.2.1 Gel purification methods. 82
2.3.2.1.1 Using NA45 paper. 82
2.3.2.1.2 Centrifuging through glass wool. 82
2.3.3 DETERMINATION OF DNA CONCENTRATION. 83
2.3.3.1 S p ectro p h o to m etry . 83
2.3.3.2 Comparison with known standards. 83
2.3.4 ETHANOL PRECIPITATION OF DNA. 84
2.4 Pr e p a r a t i o n o f ( ;e n o m i c DNA a n d p u r i f i c a t i o n
OF CLONED D N A . 84
2 .4 .1 St o c k SOLUTIONS. 8 4
2.4.2 Hu m a n g e n o m i c DNA. 85
2.4.3 PREPARATION OF BACTERIOPHAGE LAMBDA DNA. 85
2.4.4 Pr e p a r a t i o n o f p l a s m id DNA. 87
2.5 POLYMERASE CHAIN REACTION. 93
2 .5 .1 Ol i g o n u c l e o t i d e PRIMERS. 9 3
2 .5 .2 PREPARATION OF NI ICLEOTIDE STOCKS. 9 3
2 .5 .3 REACTION CONDITIONS FOR P C R AMPLIFICATION FROM
GENOMIC D N A . 9 4
2.5.4 PROTOCOL FOR PCR AMPLIFICATION FROM RNA TEMPLATE. 95
2.5.4.1 Preparation o f total RNA. 95
2.5.4.2 Sem i-quantitative RNA-PCR protocol. 95
2 .5 .5 DETECTION OF PRODUCT. 9 6
2.5.6 Su b s e q u e n t m a n i p u l a t i o n s o f PCR p r o d u c t. 96
2.5.6.1 Restriction enzyme digestion of PCR products. 96
2.5.6.2 Subcloning o f PCR products. 97
2.6 M E T H O D S OF DETECTING POLYMORPHISM. 97
2.6.1 Re s t r i c t i o n f i m g m e n t l e n g t h p o l y m o r p h i s m (RFLP). 97
2.6.1.1 Preparation of filters. 98
2.6.1.2 Preparation and ^^P labelling of probe DNA. 98
2.6.1.3 H ybridisation and washing down o f filters. 99
2.6.2 Si nCiLe St r a n dCo n i-o r m a t i o n An a l y s i s (SSCA). 100
2.6.2.1 Preparation of samples for SSCA. 100
2.6.2.2 Preparation of the gel. 101
2.6.2.3 Routine conditions for the analysis of the 5F and
F2 PCR products. 101
2.6.3 DENATURING GRADIENT GEL ELECTROPHORESIS (DGGE). 102
2.6.4 SIMPLE ACRYLAMIDE GEL ELECTROPHORESIS (SAGE). 103
2.7 Si l v e r s t a i n i n g o f a c r y l a m i d e g e l s. 103
2 .8 IN SITU HYBRIDISATION. 104
2.9 Se q u e n c i n g o f PCR p r o d u c t s. 104
RESULTS A N D C O N C LU SIO N S.
3. ANALYSIS OF LACTASE E X PR E SSIO N IN
IN T E ST IN A L SPEC IM EN S. 105
3.1 H I S T O L O G Y . 105
3.2 En z y m e a s s a y. 106
3.2.1 e v a l u a t i o n o f ASSAY PROCEDURE. 106
3 .2 .2 En z y m e a s s a y r e s u l ls in i n t e s t i n a l s a m p l e s. 108
3.2.3 ETHNIC ORIGIN OF PERSISI ENT VERSUS NON-PERSISTENT
INDIVIDUALS. 109
3.3 PR O T E IN STUDIES. 114
3.3.1 ANALYSIS OF THE LACTASE AND SUCRASE-ISOMALTASE PROTEINS BY SDS - POLYACRYLAMIDE GEL ELECTROPHORESIS
IN INTESTINAL SAMPLES FROM THE PATIENT SERIES. 114
3.3.2 La c t a s e IMMUNOHISTOLOGY. 115
3.4 M E S SE N G E R RNA S T U D IE S . 115
3 .5 Co l l e c t i o n o f b l o o d s a m p l e s a n d e s t a b l i s h m e n t
OF L YM PH O B LA SIO ID CELL LINES. 119
3.6 FURTHER ATTEMPTS TO INVESTIGATE PUTATIVE
ABNORMAL LACTASE PROTEINS. 119
3.6.1 An a l y s i s o f p a t i e n t m a t e r i a i.. 120
3 .6 .2 An a l y s i s OF SAMPLES FROM ANIMALS. 121
3.6.2.1 Do antibodies against human lactase recognise
lactase in other species? 121
3.6.2.2 Investigation of apparent dimer bands in animals. 122
4. REGIONAL LOCALISATION OF THE
LA CTA SE GENE. 132
4 .1 An a l y s i s o f s o m a t i c a n d m i c r o c e l l h y b r i d s. 132
4 . 2 In s i t u h y b r i d i s a t i o n. 138
4 . 3 Li n k a g e a n a l y s i s i n f a m i l i e s. 138
4 . 4 FURTH ER ANALYSIS OF THE FRAGMENT AND
M IC R O -C E L L HYBRID PANEL. 139
5. DETECTION OF POLYM ORPHISM W ITHIN THE
LA CTA SE GENE. 144
5 .1 INITIAL SEARCH FOR RESTRICTION FRAGMENT
LENGTH POLYMORPHISMS WITHIN THE LACTASE GENE. 145
5 .2 A n a l y s i s o f t h e 5' f l a n k i n g r e g i o n o f t h e
LACTASE GENE (5F PCR P R O D U C T ) . 146
5 .2 .1 Ad a p t a t i o n o f t h e Si n g l e St r a n d Co n f o r m a t i o n
An a l y s i s t e c h n i q u e. 146
5 .2 .2 A n a l y s i s o f t h e 5' f l a n k i n g r e g i o n (5 F ) b y SSCA. 146
5 .2 .3 ANALYSIS OF THE 5 F REGION USING DENATURING GRADIENT
GEL El e c t r o p h o r e s i s (DGGE) a n a l y s i s. 151
5 . 3 SSCA ANALYSIS OF THE FRAGMENT CONTAINING
EXON 2 (F2). 156
5.4 An a l y s i s o f t h e 3' u n t r a n s l a t e d r e g i o n (U T )
USING SIMPLE ACRYLAMIDE GEL ELECTROPHORESIS. 160
5.5 An a l y s i s o f a n M s pI RFLP w i t h e x o n 17
(LCT3 PCR P R O D U C T ) . 160
5 . 6 PRELIM INAR Y S S C A ANALYSIS OF OTHER REGIONS
OF THE LACTASE GENE. 161
5 . 6 .1 M A E II ANALYSIS OF EXONS 8 AND 11. 161
5.6.2 ATTEMPTS AT SSCA OF OTHER REGIONS. 162
5.7 DISCUSSION OF THE TECHNIQUES USED FOR THESE
A N A L Y S E S . 162
6. ANALYSIS OF DATA COLLECTED BY THE
VARIOUS PO LYM O RPH ISM DETECTION
T E C H N I Q U E S . 166
6 .1 L IN K A G E ANALYSIS WITH OTHER MARKERS ON
6 . 2 Ob s e r v e d a l l e l e f r e q u e n c i e s i n t h e
POPULATIONS STUDIED. 17 0
6 . 3 e x p l o r a t i o n o f t h e e x t e n t o f a l l e l e
A S S O C IA T I O N . 175
6 . 3 .1 Re c o m b i n a t i o n w i t h i n t h el a c t a s e GENE. 175
6 . 3 . 2 Pa i r w i s e a n a l y s i s o f a l l e l e a s s o c i a t i o n. 175
6 . 3 . 3 D e t e r m i n a t i o n OF HAPLOTYPES. 176
6 . 3 . 4 Ca l c u l a t i o n o f t h e d e g r e eo f l i n k a g ed i s e q u i l i b r i u m
OBSERVED. 178
6 . 4 As s o c i a t i o n s t u d i e s w i t h l a c t a s e
PERSISTENCE / N O N - P E R S I S T E N C E . 183
7. D I S C U S S I O N . 186
A P P E N D I C E S .
A1 Enzyme activities, histology, protein detection and mRNA
levels for all individuals tested. 200
A2 Assumed genotype results for all the CEPH parents tested. 202 A3 Assumed genotype results for the Italian series of patients
whose lactase persistence status is known. 206
A4 Assumed genotype results for the series of unrelated
individuals tested. 208
A5 Haplotypes observed in each individual of the CEPH
families studied. 212
R E F E R E N C E S . 219
Pe r s o n a l Pu b l i c a t i o n s.
1 Regional localization of the lactase-phlorizin hydrolase gene, LCT, to chromosome 2q21.
LIST OF FIGURES.
page number 1.1.5.1.1 Distribution of maltase, sucrase and lactase activities along
the length of the small intestine. 23
1.1.7.3.1 Histograms demonstrating a tri-modal distribution o f lactase
activity relative to another disacchaiidase of the small intestine. 36 1.1.9.1 Diagrammatic representation of the genomic an angement of the
lactase gene. 48
1.1.9.2 The distribution of the introns and positions of the active sites
within the repeat units of the lactase gene. 49 1.2.3.1 The genomic arrangement of the globin gene clusters. 56 1.2.4.1.1 Diagrammatic representation of the proportion of globin chains of
each type produced at different stages of development. 57
2.5.1.2 Diagrammatic representation of the position of the PCR products
within the lactase gene. 90
3.1 Diagram showing the range of tests performed on each biopsy and the blood sample where available and the treatment of the
sample in each case. 110
3.2.2.2 Tlie distribution of the lactase to sucrase ratios seen in each type
of biopsy shown in histogram format. 112
3.2.2.3 Distribution of the ratios of sucrase to lactase activity in the full
series of patients tested. 113
3.3.1.1 SDS- polyacrylamide gel electophoresis and immunoblotting of
13 representative samples with normal histology. 116 3.3.1.2 SDS-PAGE analysis of homogenates of patient samples showing
detection of an abnormal lactase pattern in individual 46. 117
3.3.2.1 Examples of lactase immunohistology results. 117 3.4.1 RNA-PCR of the 10 lactase non-persistent (N) samples tested and
8 representative lactase persistent (P) samples in comparison with the enzyme activities measured in the same individuals. 118 3.6.1 Detection of the putative dimers using modified SDS-PAGE
electrophoresis conditions. 123
3.6.2.1.2 Detection of lactase protein in animal intestinal samples using the
antibody mlac 6. 125
3.6.2.1.3 Detection of lactase protein in animal intestinal samples using
3.6.2.1.4 Detection of a protein in sheep using the antibody HBB3/705/6O which recognises the isomaltase subunit
of human sucrase-isomaltase. 126
3.6.2.2.1 Analysis of denatured and undenatured lactase protein in adult
and pre-weaned rats. 127
4.1.1 Photograph of a gel showing typical results of the PCR analysis of LC T in the fragment hybrids with a 160 base pair fragment in
some samples. 134
4.2.1 A metaphase spread showing fluorescent in situ hybridisation
using Xhchrlac7. 137
4.3.1 A photograph of an example of the results obtaining using
D2S44 to probe MspI digested genomic DNA. 141 4.4.1 Summary of the localisations determined by in situ hybridisation
for LCT, DPP4, GCG and D2S44. 142
5.1 Diagrammatic representation of the lactase gene showing the sites of previously described variation and the position of the
three known Alu elements. 143
5.2.2.1 Diagram of the 5F PCR product showing the site of the A vail
recognition site and the sizes of the digestion products. 149 5.2.2.2. SSCA of the Avail digested 5F PCR product in the presence and
absence of glycerol in the gel mix. 149
5.2.2.3 Photograph of a gel showing SSCA analysis of the A vail
digested 5F PCR product in samples from CEPH family 12. 150 5.2.3.1 Melting profile of a) the larger Avail digestion product of the
5F fragment and b) the smaller fragment. 154 5.2.3.2 Photograph of a gel showing DGGE analysis of the
A vail digested 5F PCR product in samples from the
CEPH family 13291. 155
5.3.1 Photograph of a gel showing SSCA analysis of the F2 PCR
product in samples from CEPH family 17. 157
5.4.1 Photograph of a gel showing the three phenotypes detectable for the lA polymorphism in the UT PCR product of unrelated
individuals by simple acrylamide gel electrophoresis. 158 5.5.1 Photograph of an autoradiograph produced when a filter
carrying MspI digested genomic DNA from unrelated individuals
was probed with the PCR product LCT3. 159
6.3.2.2 Photograph of a, DGGE and b, SSCA analysis of the CEPH
family 1447. 174
6.3.4.1 Diagrammatic representation of the degree of linkage disequilibrium observed across the coding region of the
lactase gene. 182
7.1 Results of the analysis of RNA and DNA in the same individuals in order toidentify whether the lactase persistence polymorphism is controlled by a cis- or a trans- acting factor 198 7.2 Positions of the samples which demonstrate loss o f an allele 199
LIST OF TABLES.
page number
1.1.7.1 Table of the frequency of lactase non-persistence in a few selected ethnic groups as determined by lactose tolerance tests
in different studies. 29
1.1.7.1.1 Review of the world literature on the inheritance of the ability
to digest lactose. 30
1.1.7.1.2 Summaty of the total numbers of progeny of different lactose
digestor phenotype. 31
1.1.7.2.1 The observed frequencies of the lactase persistence phenotypes in dizygotic twins and the expected frequencies calculated from the population frequency of lactase non-persistence in the general population of Budapest, Hungary; assuming a
monogenetic model. 35
1.1.7.3.2 Table showing the observed numbers of individuals in the three 'activity' groups, together with the numbers that would have been expected for a population in Hardy-Weinberg equilibrium. 37 1.3.2.1 Examples of the genes where SSCA has been successfully used
to identify mutations/vaiiations. 66
2.5.1.1 Sequence and position of the oligonucleotide primers within the
lactase gene. 89
2.5.1.3 Oligonucleotide primer sequences used for amplification of genes
other than lactase. 91
2.5.3.1 The expected size, in base pairs, of each lactase PCR product, and the annealing conditions used in the PCR reaction and the methods used to conl’irm the identity of the product. 92
2.6.2.3.1. Table of the routine conditions used for the analysis of the
5F and F2 PCR products. 101
3.2.1.1 Lactase and p-galaciosidase activities in the presence and
absence of the inhibitor PCMB. I l l
3.2.2.1 Means (±1SD) and the ranges of values obtained for pinch and
crosby capsule biopsies. 112
3.6.2.1.1 Summary of the cross reaction of the anti human lactase and anti human sucrase-isomaltase antibodies with denatured protein
3.7.1 The cross reaction of anti human lactase antibodies with native lactase from other species as judged by enzyme immunobinding assay and immunoprécipitation expenments. 131
4.1.2 Segregation of LCT in a series of somatic cell hybrids. 135 4.1.3 Segregation of LCT, DPP4 and other chromosome 2 markers
in the micro cell and fragment hybrids. 136
5.7.1 Table of the alleles at different sites within the LC T gene and whether they were detected by RFLP analysis, SSCA,
DGGE or SAGE. 165
6.1.1 Lod scores at different values of the recombination fraction 0 for
LC T and D2S44. 168
6.1.2 Pairwise lodscores at maximum likelihood recombination fraction 0 in females and males for LC T with other
chromosome 2 markers on the CEPH database, version 6. 169 6.2.1 Frequencies of the various alleles detected within the LCT gene. 171 6.3.2.1 Tables of the associations between alleles in the four fragments
of the lactase gene that were analysed. 172/3 6.3.3.1 Tabular representation of the haplotypes observed in the whole
CEPH population. 179
6.3.3.2 Observed numbers and frequencies of the haplotypes observed
in the two major sub-populations of the CEPH series. 180 6.3.3.3 Frequencies of the haplotypes observed in the whole CEPH
series in comparison with those expected from random
assortment of the alleles. 181
6.4.1 Tables of the observed ocun ence of each phenotype in
Ar r r e v ï a t ï o n s
SDS-PAGE sodium dodecyl sulphate - polyacrylamide gel electrophoresis
PMSF phenylmethylsulphonyl tluoride
TPCK L-1- tosylamide-2-phenylethyl-chloromethylketone
TLCK N a p tosyl L lysine chloromethyl ketone
PCMB p- chloromercuribenzoic acid
PCMS p- chloromercurisulphonic acid
IgG immunoglobulin gamma
EDTA ethylenediamineteira-acetic acid
SSCA single strand conformation analysis
DGGE denaturing gradient gel electrophoresis
CDGE constant denaturing gel electrophoresis
SAGE simple acrylamide gel electrophoresis
RFLP restriction fragment length polymorphism
HOT hydroxylamine / osmium tetroxide (chemical cleavage)
r.p.m. revolutions per minute
g unit of centiifugal force (gravity)
Mr relative molar mass
SI sucrase-isomaltase
MG maltase-glucoamylase
LPH lactase phlorizin hydrolase
PCR polymerase chain reaction
Tin melting temperature
LCT lactase gene
DPP4 gene encoding DPPIV (dipeptidyl peptidase IV)
GCG glucagon gene
A CKNOWI.EDGEMENTS
I would like to express my appreciation of my supervisor Dallas Swallow, for
her advice, guidance, encouragement and friendship throughout my time in the MRC
HBGU. I am also indebted to Prof. E.B. Robson for her encouragement and for
organising the funding which enabled me to undertake this work.
I would like to thank all my colleagues, in the MRC HBGU past and present, for
their help, advice and friendship. In particular I thank the colleagues whose
collaborative work I have included: Yangxi Wang for the RNA-PCR work, M argaret
Fox for the fluorescent in situ hybridisation, Lynn Hughes for the lactase
immunohistology, Wendy Pratt for sequencing and Dalila Darmoul the work on DPP4. I am also grateful for the assistance of Steve Jeremiah who performed the GCG PCR
and many of the DNA extractions, and Ira Islam who performed a very large number of
PCRs and enzyme digestions and also analysed the UT product. I thank David
W hitehouse for useful and thought provoking discussions on linkage disequilibrium. I
also appreciated the help of Phil Johnson in the establishment of the conditions for the
DGGE analysis, especially with respect to the computer programs and to John Attwood
for aiding my use of the CEPH/EUROGEM computing facilities. I am also grateful to
Lynne Sargent for proof reading this manuscript, although none of the remaining
mistakes are her fault!
I wish to acknowledge the collaboration of Dr. Martin Sarner, Dr. Virginia
Sams and their colleagues in the work described in Chapter 3, Professor S. Auricchio
and co-workers who have provided samples from Italians of known lactase persistence
phenotype and Dr. T. Iqbal and Dr. B. Cooper, Dudley Road Hospital, Birmingham
have provided samples from the Indian population of West Birmingham.
I also acknowledge CEPH and Eurogem for the family DNAs, and the MRC
Human Genome Mapping Project for providing the studentship as well as other
1. IN T R O D U C T IO N .
This thesis is concerned with the biochemical and genetical analysis of the
disacchaiidase lactase phlorizin hydrolase (LPH). Lactase is present in mammalian small
intestine where it is responsible for digesting lactose, the sugar found in milk.
In the first pait of the introduction I shall review the current published information
available as to the pattern of expression of LPH activity along the length o f the intestine
and during development in a vai iety of mammalian species. Comparable information for
other disaccharidases of the small intestine, namely sucrase-isomaltase, maltase-
glucoamylase and trehalase will be considered where relevant. In humans, lactase
activity persists into adult life in some people but not in others. The evidence that this is a
genetically deteimined trait will be reviewed, as will the indirect methods used to
determine lactase persistence status in population studies. The gene which codes for the
lactase enzyme has been cloned and the nature of the sequence and the deduced properties
of the protein produced are discussed.
In tlie second pait of the introduction variations in other proteins and the
underlying nucleotide changes which cause them aie considered. In particular, other
examples of abnoimal expression of temporally regulated products are discussed.
Thirdly, the variety of techniques available for the detection of single base
changes is reviewed.
The final section of the introduction describes the principles and uses of linkage
1.1. EXPRESSIO N AND CHARACTERISTICS OF TH E E N Z Y M E LACTASE PHLORIZIN HYDROLASE.
1.1.1. CHARACTERISTICS OF THE ENZYME.
Lactase-phlorizin hydrolase (LPH) (E.C. 3.2.1.23/62) is one o f a group of
hydrolases found characteristically in the brush border membrane of small intestinal
enterocytes in a number of mammalian species. LPH catalyses the hydrolysis of the (3-
galactoside lactose to glucose and galactose; it also has p- glucosidase activity and
hydrolyses phlorizin (4,6-dihydroxy-2(p D glucosido)-p(p-hydroxyphenyl)
propiophenone) at a different active site. The existence of two active sites was suggested
by the differential inhibition of the two activities by Tris (Skovbjerg etaL , 1981), by obseiwation that lactose hydrolysis is inhibited by phlorizin whereas phlorizin hydrolase
activity is unaffected by lactose (Skovbjerg et al., 1981; Lau, 1987) and by the differing thermal stabilities of the two activities (Skovbjerg et at., 1981; Lau, 1987). Direct evidence of the existence of two active sites was obtained from inhibitor studies using
^H-conduritol-B-epoxide (see section 1.1.8. l.)(W acker et at., 1992). The notion that the two active sites were located on the same polypeptide was supported by the failure to
separate these two activities under all conditions investigated (Bolton et al., 1983; Potter et al., 1985). This has been confinned more recently by transfection studies (see section 1.1.8.1.)(Naim et al., 1991).
The lactase phlorizin hydrolase polypeptide has been shown by metabolic
labelling studies to be synthesised as a single chain precursor forni of apparent
200,000 - 215,000, which is subsequently Cleaved by intra-cellular proteases to give the
mature forni found in the brush border, of apparent M^. 140,000 -160,000 (Naim et al., 1987). There is some evidence that this cleavage occurs in the Golgi apparatus (Naim et a i, 1987; Lottaz et al., 1992; reviewed by Naim, 1993; in pig - Danielsen et al., 1984). However the location, within the cell, of this cleavage is more controversial or possibly
1.1.2. EVIDENCE FOR DIMERISATION OF THE LACTASE PROTEIN.
It is thought that lactase may be present as a dimer in the bioish border membrane.
A significant difference in mobility consistent with a 2-foId difference in size was
observed when lactase was analysed under nondenaturing conditions (Potter et a l, 1985). There is also some evidence of the existence o f dimers from SDS-PAGE studies
following crosslinking using dimethyl adipimidate (Danielson, 1990). The amino
terminal sequence of the putative dimer identified after immunoprécipitation was found to
be identical to the sequence of the mature protein, which suppoits the notion that this
component is a dimer rather than a precursor foiTu (Sterchi et al., 1990). This suggestion of lactase being present as a dimer is also supported by the obseiwation of two adjacent,
identical stmctures in electron microscopic studies of a purified lactase preparation
(Skovberg et al., 1981).
1.1.3. GLYCOSYLATION OF THE LACTASE POLYPEPTIDE.
A number of lines of evidence have shown LPH to be a glycoprotein both in man
and other species. For example purified or immunoprecipitated human lactase protein can
be detected with the lectin concanavalin A indicating the presence of a-mannose residues
(Bolton e t a l , 1985) and terminal ABH and Lewis blood group structures are present according to the blood group of the individual (Triadou et al., 1983; Green et al., 1988). Naim etal. (1987) obtained evidence using metabolic labelling and enzyme treatments for the precursor foi*m of LPH canying high mannose glycan units and the mature form
showing complex glycosylation. Subsequently, the presence of two differently
glycosylated mature foiTns of LPH - an N-glycosylated foiTn and an N -/0- glycosylated
form have demonstrated by lectin binding studies (Naim & Lentze, 1992). These forms
could be separated by Helix pomatia lectin column chromatography. Kinetic studies showed that the two foiTns have the same Kj„s but the N -/0 - glycosylated form has a
Vujax approximately four times that of the N-glycosylated form. The O-glycosylation
was said to vary between different individuals (Naim & Lentze, 1992) but how this
related to their blood group was unclear.
1.1.4. OTHER INTESTINAL DISACCHARIDASES.
Several other disaccharidases with different substrate specificities are present on
the brush border membrane of the small intestine and are considered below. O f these,
sucrase-isomaltase (SI) will be considered here in most detail since this enzyme has been
analysed in comparison with lactase in many of the studies described within this thesis
and also in other studies in the literature.
1.1 .4 .1 . S u c r a s e - is o m a lta s e .
Sucrase -isomaltase (EC.3.2.1.10 and 3.2.1.48)(SI) catalyses the hydrolysis of
sucrose to glucose and fructose and also of isomaltose to two glucose moieties. The SI
protein has been shown to be synthesised as a single, enzymically active polypeptide
chain of apparent M,. 220,000 (Sjostrom 6'rr//., 1980). This precursor, pro-sucrase-
isomaltase, caiiies two distinct active sites, one for sucrose and one for isomaltose. The
SI precursor has been shown to be cleaved by pancreatic proteases into two subunits of
slightly different sizes apparent M ,-130,000 - 160,000 (Sjostrom et al., 1980; Hauri et a l, 1980). These subunits stay associated in the mature protein but can be dissociated by the action of agents such as sodium dodecyl sulphate. The larger (M,. 140,000) subunit
has been identified as possessing the isomaltase activity and the smaller (M^ 120,000) the
sucrase activity (Conklin et at., 1975; Brunner et a l, 1979). The active sites of these activities have been localised to aspaitic acid residues at amino acids 505 (isomaltase) and
1394 (sucrase) (Hunziker et al., 1986; W acker et a l, 1992). Analysis using enzyme treatments have shown that SI is inserted in the membrane in a Nj„ orientation
been shown to be glycosylated (Ghersa et al., 1986) and like lactase carries ABH blood group structures (Green et al., 1988).
The cDNA encoding SI (in rabbit -Hunziker et al., 1986; in humans - Green et a l, 1987; Chan tret et al., 1992) and the 5' genomic structure have been characterised (in humans - Chantret et al., 1992; Wu et al., 1992). The human 81 mRNA is 6 kb long (Green et al., 1987) and encodes a protein of moleculai* weight (M,. ) 209,402 comprising 1827 amino acids. There is no cleaved signal sequence in the rabbit (Hunziker et a l, 1986). It is noteworthy that the sequence shows two fold internal homology with
isomaltase encoded by the N-tenninal repeat and sucrase by the second (Hunziker etal., 1986; Chantret eta l., 1992).
1.1.4.2. M alta se-g lu co a m y la se.
Maltase-glucoamylase (EC.3.2.1.20) catalyses the hydrolysis o f maltose and
other oligosaccharides with a 1 -> 4 glucopyranosidic bonds. In pigs the enzyme has
been shown to be synthesised as a single polypeptide chain of apparent molecular weight
240,000, which is subsequently cleaved by pancreatic proteases into two subunits
(Sorensen et al., 1982). Two distinct active sites which differ in heat stability were observed and an N-terminal anchor identified (Noren et a i, 1986). In contrast,
experiments on human maltase-glucoamylase identified a protein of appaient molecular
weight 335,000 which was not proteolytically processed (Naim et al., 1988; Hauri eta l., 1985a). This protein was shown to be glycosylated in humans (Naim et al., 1988).
1 .1 .4 .3 . T re h a la se .
Trehalase (EC. 3.2.1.28) catalyses the hydrolysis of a a trehalose (1-0-a D
glucopyranosyl-a D glucopyranoside), which is found mainly in insects and
mushrooms. Deficiency of this enzyme activity has been reported in a family and was
The enzyme is a minor component of the small intestine and renal brush border
membranes (Galand, 1984). It has been shown by standard SDS-polyacrylamide gel
electrophoresis to have apparent molecular weight 75,000, and to be a glycoprotein
(Galand, 1984). The cDNA coding for trehalase has been cloned in rabbit, and was
found to encode a protein of M,. 65,516 within a mRNA of 1.8 kb (Ruf et al., 1990). Thus both the mRNA and the protein are much smaller than those for the other intestinal
hydrolases. Indeed, there is no internal homology within trehalase (Ruf et a l , 1990) unlike both sucrase-isomaltase (section 1.1.4.1.) and lactase-phlorizin hydrolase (section
1.1.8.1.). Expression of the rabbit trehalase mRNA in Xenopus laevis oocytes was observed after micro injection of the capped mRNA. The results of experiments
involving solubilization of the enzyme from the oocyte surface membrane by treatment
with phosphatidylinositol-specific phospholipase C from Bacillus thuringiensis suggest that trehalase is anchored in the membrane via glycosylphosphatidylinositol (Ruf et ah, 1990). This is again in contrast to SI, LPH and maltase-glucoamylase which are
anchored via peptide regions which span the plasma membrane (sections 1.1.4.1 (SI),
— M 0110 s e
Sucrose
“ » L o c f o s e
4 8
4 2
3 9
I B O
- L___
200 2 20 2 4 0 2 6 0 2 8 0
U t 140
D u o d e n u m
l
::
j
I____
. C mp o s t
1,0
C m f r o m t e e t h t o c o p s u l e
\
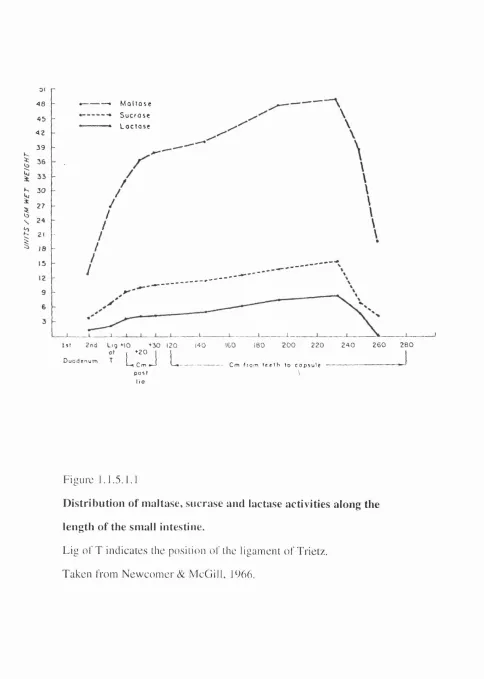
F i g u r e 1.1.5.1.1
D is tr ib u tio n o f m a lta s e , s u c r a s e a n d la cta se a c tiv itie s a lo n g the l e n g th o f the sm a ll intestine.
L i g o f T i n d i c a t e s t he p os i t i o n o f t h e l i g a m e n t o f T r i e t z .
T a k e n f r o m N e w c o m e r & M c G i l l , 1966.
1.1.5. VARIATION IN ENZYME ACTIVITIES WITHIN THE SMALL INTESTINE.
Lactase protein is believed to be found exclusively in the small intestine, although
the evidence for this in man is somewhat limited. It is expressed in the enterocytic cells
of the intestinal villus. Indeed, it been shown by histochemistry and immunohistology to
be specifically localised on the brush border (Lojda & Kraml, 1971; Maiuri e ta l., 1991). The patterns of expression of lactase and other enzyme activities vary along the length of
the intestine as well as along the vertical axis of a single villus in humans and other
animals.
1.1.5.1. Variation in lactase and other enzvme activities within a villus.
The pattern of expression of the disaccharidases lactase, maltase, sucrase,
trehalase and also of alkaline phosphatase activity along the crypt-villus axis in humans
was examined in a study by Nordstrom & Dahlqvist (1973). All the disaccharidase
activities except lactase showed a similar pattern of expression with little activity in the
crypt, a peak of activity mid-way up the villus and a decline to the villus tip in both
duodenum and proximal Jejunum. Lactase activity was examined only in the proximal
jejunum, but was detemnined both in individuals with high lactase activity and in an
individual with low lactase activity. The peak of lactase activity was slightly more apical
than that of the other disaccharidases but was identical in both cases. The peak of
alkaline phosphatase activity was distinctly more apical than the peak of maltase, sucrase
and trehalase activities in duodenum and jejunum.
1.1.5.2. V ariation in lactase and other enzvme activities within the small intestine in humans.
The pattern of variation in level of LPH activity along the length of the small
intestine was first described by Newcomer & McGill (1966)(Figure 1.1.5.1.1.). The
and greatest in the jejunum. This same pattern of variation of specific activity along the
intestine was also seen for two other small intestinal disaccharidases, sucrase- isomaltase
and maltase (Figure 1.1.5.1.1; Newcomer & McGill, 1966).
1.1.5.3. Variation in LPH activity within the small intestine in other
a n im als.
Lactase activity is found in the intestine of most mammals but not in birds and
amphibia (Plimmer, 1906; Parsons & Pritchard, 1965; Keriy, 1969; reviewed by
Semenza, 1967). However, in mammals that do not possess lactose in their milk, that is
sealions (group Pinnipedia)(Pilson & Kelly, 1962), no typical lactase activity was found
even in neonates (Sunshine & Kretchmer, 1964; Crisp et a i, 1987).
In all mammals which have lactase activity in the small intestine, the pattern of
expression of lactase along the length of the intestine was similar to that found in
humans, with maximal activity in the mid intestine and lower levels at the proximal and
distal ends. Mammals studied include rat (reviewed in Henning, 1985), rabbit (Keller et a l, 1992; Semenza et a l, 1993), baboon (Welsh et a l, 1974), galago monkey (Wen et a l, 1973), dog (Welsh & Walker, 1965), cattle (Le Huerou et a l, 1992) and pig (A. Collins, personal communication).
It was first reported some 90 years ago that the level of lactase activity is higher in
the young animal than in the adult (for example Plimmer, 1906; Koldovsky et a l, 1966; Hore & Messer, 1968). The most detailed studies of the change in lactase activity during
development have been performed on samples from rat. In rat maximal lactase activity is
found in the neonate, such that the specific activity of LPH in the adult is 20% of that
found in 7 day old rats (Nudell et a l, 1993). It has been shown that this decline in activity happens at or around the time of weaning (for example Welsh & Walker. 1965;
1.1.6. DEVELOPMENTAL REGULATION OF THE LPH PROTEIN IN MAN.
In humans, as in other mammals (discussed in section 1.1.5.3), lactase activity is
high after birth until around the time of weaning, and declines to a lower level in adult life
in most of the world's population (reviewed in Flatz, 1987 and section 1.1.7).
However, in most Caucasians and in certain other populations the level of lactase activity
remains high throughout adult life. Initially it was thought that possessing lactase activity
in adulthood was the normal situation, with the first case of "alactasia" being reported by
Holzel et al. in 1959. Gradually it became clear that a low level of lactase activity was not a rare condition but nonnal for adults of most races. This condition may be called lactose
intolerance, since when the enzyme activity is low the individual is unable digest lactose,
and this can lead to symptoms of varying intensity, noiTnally abdominal distension or
discomfort, flatus and diarrhoea on consumption of lactose containing foodstuffs.
Because intolerance of lactose and lactase deficiency can have various causes, specific
lactase deficiency manifesting in the adult is more specifically referred to as lactase non
persistence. The term adult-type hypolactasia has also been used to indicate this
situation.
Early studies using density gradient centrifugation showed that the lactase activity
in adults and in infants gave identical sedimentation patterns (Gray & Santiago, 1969).
The kinetic properties of the lactase enzyme from adults with persistent lactase activity,
from adults non-persistent for lactase activity and from infants were also similar
(Lebenthal et at., 1974). Immunotitration and crossed Immunoelectrophoresis
experiments suggested an absence of any immunological difference between the adult and
infant enzyme and between enzyme from adults of different race (Potter et at., 1985). These results thus suggested that only one lactase enzyme was involved rather than there
being two different enzymes, one of which was expressed in adults and the other in
1.1.6.1. L a c to se to le ra n c e te sts.
The most definitive test for lactase persistence status is to assay the lactase activity
in a biopsy taken from the jejunum. However, to allow for the variability in activity due
to regional differences within the intestine (see section 1.1.5.2) as well as to control for
the quality of the sample, lactase activity is usually expressed as a ratio of lactase activity
to the activity of another intestinal disaccharidase (Welsh etaL, 1978; Ho etal.y 1982; Flatz, 1984). Furthermore, it is important to monitor for villous damage in pathological
specimens since such damage may cause secondary disacchaiidase deficiency (as seen for
example in coeliac disease). It is noteworthy that lactase activity is often lost before the
other disaccharidase activities.
Invasive tests such as taking a biopsy of the small intestine are not suitable for
population studies so various fomis of oral test for lactose tolerance have been
developed. In general, a lactose challenge of 5()g is given to an adult after an overnight
fast and then measurements are made of some parameter at different time points.
Symptoms experienced by the subject are also noted.
The products of hydrolysis of lactose, namely glucose and galactose, may be
detected in the blood or urine from an individual. Direct detemiination of the rise in the
level of plasma glucose can be made using either venous or capillaiy blood. In a study in
which these two sampling methods were compared the authors claimed that the use of
capillaiy blood gave a more accurate diagnosis of lactase persistence status (McGill &
Newcomer, 1967). Alternatively, the rise in plasma galactose can be measured (for
example Fischer & Zapf, 1965). More recently, a strip test for measuring urinary
galactose has been developed (Ai'ola et a l, 1987) which requires the administration of alcohol with the test to block the removal of galactose by the liver. Thus the galactose
absorbed across the brush border is excreted into the urine where the level can be
assayed. This test has the advantage of being non-invasive but the need for
is peiTnitted. An indirect method for detecting lactase persistence/non-persistence status
depends on the femientation of the undigested lactose by the gut flora in the colon and
measurement of the expired hydrogen gas produced by this bacterial action (this is known
as the breath hydrogen test). Administration of a mixture of glucose and galactose in
place of the lactose may be used as a control. This identifies individuals who are able to
digest the lactose but in whom the constituent monosaccharides are not absorbed.
The sensitivity and specificity of these detection methods have been evaluated by
a number of investigators. Even though the best discrimination and accuracy was
obtained using the galactose/ethanol test, in two of the studies breath hydrogen
determination was recommended since it is the least invasive technique. However, all
authors who have used the breath hydrogen technique emphasise that it must be
administered with care (Newcomer et al., 1975; Metneki et a i, 1984). In general, lactose tolerance was found to agree well with the enzyme assay results obtained from biopsy
Ethnic group (incl country of
test) number of individuals tested % non-persistent Reference Ugandan Ankole
24 25 Cook & Kajubi,
(1966)
Ugandan Bantu 35 89 Cook & Kajubi,
(1966)
S. African Bantu
22 95 Jersky et al.,
(1967)
Thais 100 96 Flatz &
Saengudom, (1969)
Nigerian Yomba
41 98 Kretchmer
et a /.,1971 Inuit of
Greenland
219 55 Gudmand-Hoyer
e ra /. (1973)
Finns 159 17 Sahi et al.
(1973)
Gernian 65 14 Flatz
(1984)
N. American Blacks
41 83 W elsh et al.
(1978)
N. American Whites
222 28 W elsh et al.
(1978)
Whites from Birmingham
67 3 Iqbal et al.,
(1993)
Indians in Birmingham
55 51 Iqbal et a l,
(1993)
Table 1.1.7.1
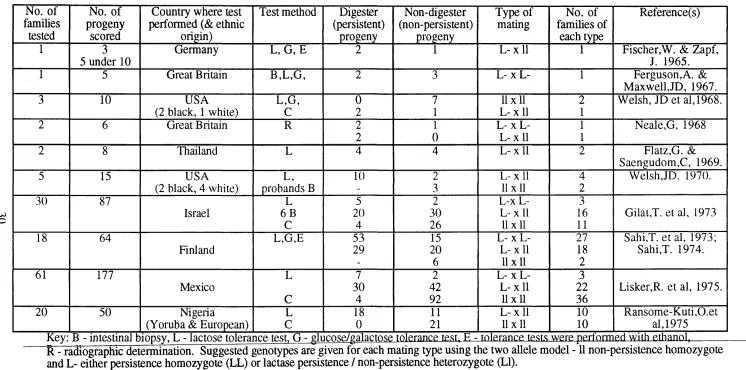
No. of families
tested
No. of progeny
scored
Country where test performed (& ethnic
origin)
Test method Digester (persistent) progeny Non-digester (non-persistent) progeny Type of mating
No. of families of
each type
Reference(s)
1 3 Germany L, G ,E 2 1 L - x ll 1 Fischer,W. & Zapf,
5 under 10 J. 1965.
1 5 Great Britain B ,L,G , 2 3 L- X L- 1 Ferguson,A. &
Maxwell,JD, 1967.
3 10 USA L,G , 0 7 11x11 2 Welsh, JD et al, 1968.
(2 black, 1 white) C 2 1 L - x ll 1
2 6 Great Britain R 2 1 L- X L- 1 Neale,G, 1968
2 0 L - x ll 1
2 8 Thailand L 4 4 L - x ll 2 Flatz,G. &
Saengudom,C, 1969.
5 15 USA L, 10 2 L - x ll 4 Welsh,JD. 1970.
(2 black, 4 white) probands B - 3 11x11 2
30 87 L 5 2 L-x L- 3
Israel 6 B 20 30 L - x ll 16 Gilat,T. et al, 1973
C 4 26 11x11 11
18 64 L,G,E 53 15 L- X L- 27 Sahi,T. et al, 1973;
Finland 29 20 L - x ll 18 Sahi,T. 1974.
- 6 11x11 2
61 177 L 7 2 L- X L- 3
Mexico 30 42 L - x ll 22 Lisker,R. et al, 1975.
C 4 92 11x11 36
20 50 Nigeria L 18 11 L - x ll 10 Ransom e-K uti,0.et
(Yoruba & European) C 0 21 11x11 10 al, 1975
Key: B - intestinal biopsv. L - lactose tolerance test. G - glucose/galactose tolerance test, E - tolerance tests were perfniTned with ethanol,
R - radiographic determination. Suggested genotypes are given for each mating type using the two allele model -11 non-persistence homozygote and L- either persistence homozygote (LL) or lactase persistence / non-persistence heterozygote (LI).
Table 1.1.7.1.1.
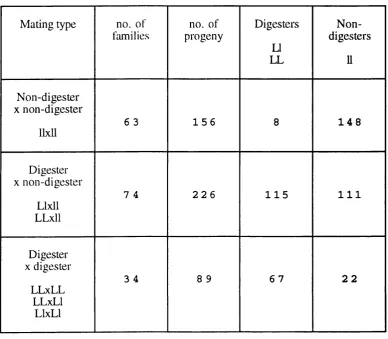
M a t i n g t y p e n o . o f f a m ilie s
n o . o f p r o g e n y
D i g e s t e r s
U L L
N o n d i g e s t e r s
U
N o n - d i g e s t e r X n o n - d i g e s t e r
11x11
6 3 1 5 6 8 1 4 8
D i g e s t e r X n o n - d i g e s t e r
L lx ll L L x l l
7 4 2 2 6 1 1 5 1 1 1
D i g e s t e r X d i g e s t e r
L L x L L L L x L l
L l x L l
3 4 8 9 6 7 2 2
Table 1.1.7.1.2
Sum m ary o f the total num bers o f progeny o f different lactose digester
p h e n o ty p e .
The information is taken from the family studies in Table 1.1.7.1.1. Suggested
genotypes as in Table 1.1.7.1.1, using 2 allele model are shown LL - persistence
homozygote, LI - lactase persistence / non-persistence heterozygote and 11 non-persistent
1.1.7. EVIDENCE FOR MENDELIAN INHERITANCE.
The frequency of lactase non-persistence has been determined in a large number
of studies in different populations of the world. The frequency has been shown to vary
between populations, ranging from almost 100% in Thais to below 10% in certain
Northern European populations. These data have been reviewed by Simoons (1978) and
Flatz (1987). A selection of the populations that have been studied are shown in
1.1.7.1.
Table
Various studies have been perfoimed to investigate whether lactose tolerance is an
inherited characteristic. They fall into three categories, namely family studies, twin
studies and population studies. The results of these studies are discussed in the
following sections (also reviewed in Swallow & Harvey, 1993; see appendix).
1.1.7.1. E vid en ce from fam ily studies.
The evidence that lactase persistence is a genetically deteimined polymorphism
comes mainly from family studies. The results of these studies are summarised in Table
1.1.7.1.1. A few of these studies have involved direct determination of lactase activity in
the intestine e.g. Ferguson & Maxwell, 1967. However, the majority of families studied
from a wide variety of geographic regions of the world have been examined using an
indirect lactose tolerance test (discussed section 1.1.6.1). These include progeny of
mixed marriages (Flatz & Rotthauwe, 1971).
Three types of family have been studied: families in which both parents are lactase
persistent (i.e. lactose digestors); families in which one parent is persistent and the other
non-persistent and families in which both parents are non-persistent. The results of all
the family studies detailed in Table 1.1.7.1.1 are summarised in Table 1.1.7.1.2 for each
of the available mating types. The balance of evidence from these family studies is
single gene locus where lactase persistence is dominant to non-persistence, which is
recessive. The only results which conflict with this model come from the occunence of a
few individuals in two of the studies who are lactase persistent despite having two non-
persistent parents. In both the study of Gilat, et ciL, 1973 (4 individuals) and that of Lisker et a!., 1975 in Mexico (4 individuals), the possibility that one or other o f the parents had a secondaiy deficiency of lactase was not excluded, nor was the possibility of
non-patemity. Lisker et al. also suggest that these exceptions may be due to errors in the glucose detection method used (95% accuracy estimated, Lisker et al. 1974). It should be noted that the number of lactose non-digester (intolerant or hypolactasic) progeny is
greater in each case than would have been expected for each type of mating. However
this can readily be explained by ascertainment bias(s) since in almost all the studies
families were ascertained through non-persistent individuals. Sahi, (1974) showed, in
his family study, that the numbers of individuals of each phenotype agreed reasonably
well with expectation when this bias was allowed for.
1.1.7.2. E vidence from twin studies.
Another line of evidence in support of the single gene mode of inheritance was
provided by twin studies (Metneki et al., 1984). In this study o f 102 pairs of Hungarian twins it was found that monozygotic twins showed 100% concordance of lactase
persistence phenotype. In the dizygotic twins the proportions of lactase persistence and
non-persistence phenotype were consistent with expected results (shown in Table
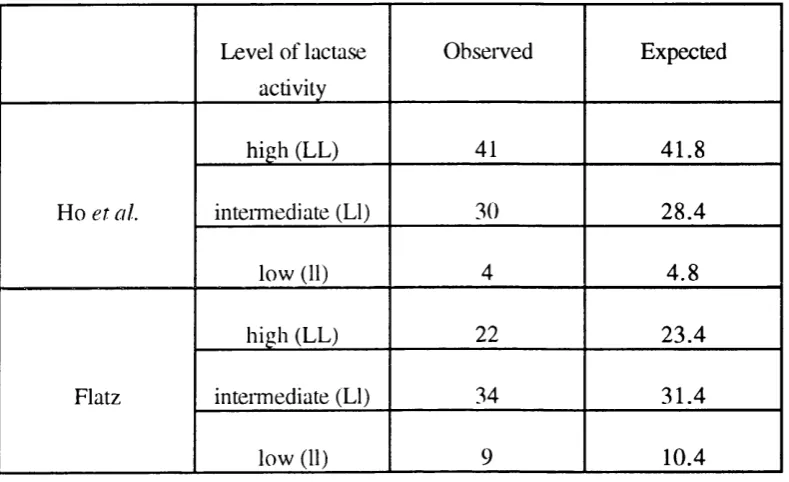
1.1.7.3. E v id en c e fro m p o p u la tio n stu d ie s.
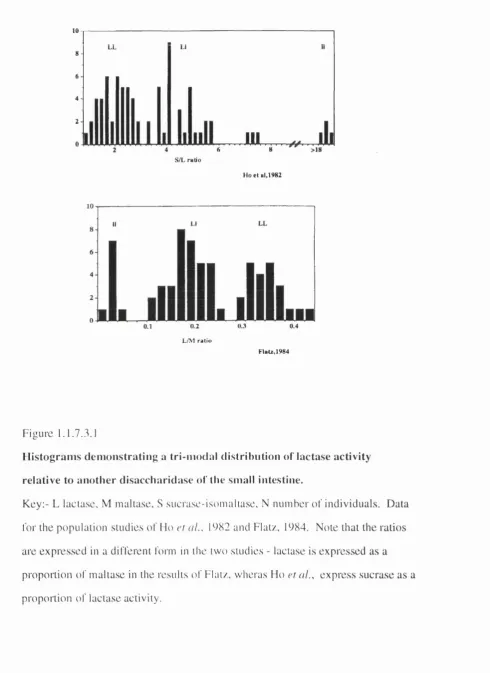
Supporting evidence for monogenetic inheritance is also provided by
population studies in which lactase activities were measured directly on intestinal
material. A trimodal distribution of activities was obseiwed in adult intestinal samples,
when sucrase/lactase ratios in post-moitem tissue (Ho et al., 1982) or lactase/maltase ratios in healthy volunteers (Flatz, 1984) were determined (Figure 1.1.7.3.1). In both
these studies, which in each case involved native Europeans, the frequencies of the
individuals within each of these groups were entirely consistent with the expected
frequency for the two types of homozygotes and a group of heterozygotes (see Table
1.1.7.3.2). The level of activity observed in the putative heterozygotes was
approximately half that of the persistent homozygotes, although both of the groups
concordant discordant
persistent / persistent
non-persistent / non-persistent
persistent / non-persistent
Observed 27 11 12
Expected 26.Ü 11.1 12.9
Table 1.1.7.2.1.
The observed frequencies o f the lactase persistence phenotypes in dizygotic tw ins and the expected frequencies calculated from the
population frequency o f lactase non-persistence in the general population o f B udapest, H ungary; assum ing a m onogenetic m odel.
Similai’ results were obtained using the I’requencies detemnined in the two separate
10
LL
8
6
4
2
0
>18 S/L ratio
Ho e t al,1982
L/M ratio
FlaU,1984
F i g u r e 1.1.7.3.1
H i s t o g r a m s d e m o n s t r a t i n g a tr i- m o d a l d is tr ib u tio n o f la c ta s e a c tiv ity r e la tiv e to a n o t h e r d is a c c h a r id a s e o f tiie s m a ll intestin e.
K e y ; - L lacta.se, M m a l t a s e , S s u c r a s e - i s o m a l i a s e , N n u m h e r o f i n d i v i d u a l s . D a t a
l o r t h e p o p u l a t i o n s t u d i e s o f H o et ///., 1982 a n d Flat/., 1984. N o t e t h a t t h e r a t i o s
a r e e x p r e s s e d in a d i f f e r e n t fo rm in th e t w o s t u d i e s - la c t a s e is e x p r e s s e d a s a
p r o p o r t i o n o f m a l t a s e in th e r e s u l t s o f F l a t / , w h e r a s H o et al . , e x p r e s s s u c r a s e a s a
p r o p o r t i o n o f l a c t a s e a c tiv ity .
Level of lactase activity
Observed Expected
Ho et al.
high (LL) 41 41.8
intemiediate (LI) 30 28.4
low (11) 4 4.8
Flatz
high (LL) 22 23.4
intemiediate (LI) 34 31.4
low (11) 9 10.4
Table 1.1.7.3.2.
Table show ing the observed num bers o f individuals in the three 'activity' groups, together with the numbers that would have been expected for a population in H ardy-W einberg equilibrium .
The gene frequencies were calculated assuming that the groups had genotypes LL (high
activity), LI (intennediate activity) and 11 (low activity). The data are taken from the
population studies of Ho et al. (1982) and Flatz (1984) (data derived from tables in these papers). Note that the ratios in these two studies are inverted in relation to each other,
1.1.8. CHARACTERISTICS OF THE LACTASE MESSENGER RNA AND THE
RESULTING PROTEIN.
1.1.8.1. M olecular characteristics o f the protein deduced from the cDNA
s e q u e n c e .
The complete cDNA sequence coding for human LPH was found to be 6274
nucleotides long and contained an open reading frame encoding a polypeptide o f 1927
amino acids (Mantei et a l, 1988). This sequence was deduced to comprise a cleavable signal sequence, a C terminal hydrophobic sequence and the remaining sequence
exhibited a weak four fold internal homology. These repeats have been thought to
suggest duplication events in the evolutionary past. Indeed, the repeat unit has been
found to be a member of a family of sequences which includes the bacterial enzyme p-
glucosidase bglA of Clostridium thermocellum (Grabnitz e t a l , 1991).
The LPH protein appears to be translated, in humans, as a 1927 amino acid pre-
pro-fomi of calculated molecular weight 218,6(K). There is predicted to be a cleaved
signal sequence of 19 amino acids at the N-temninal end. The glycosylated pro-LPH
migrates on SDS-PAGE gels with an apparent molecular weight 215,000. In turn pro-
LPH is processed to a mature forni which is comprised of amino acids 867-1926 of the
pre- pro-foiTn. The mature protein has an apparent molecular weight 160,000,
corresponding to a calculated polypeptide molecular weight of 121,324.
The cloning of the lactase cDNA also provided direct evidence of the presence of
both active sites on the single lactase polypeptide. This was demonstrated by transfection
of a full length cDNA, in an appropriate vector, into COS cells and the detection of both
activities (Naim et a l, 1991). Direct evidence of the existence of two distinct active sites was obtained from the protein by inhibitor studies using ^H-conduritol-B-epoxide
and 1747. These sites correspond to nucleotides 3828/30 in exon 8 and nucleotides
5256/8 in exon 15 (nucleotide positions are those in cDNA sequence). Residue 1271
was assigned to lactase activity and 1747 to phlorizin-hydrolase as a result of calculations
from both the degree of inactivation of the enzyme and the amount o f radioactivity bound
to each site. These sites are similar to the site V-I-S-E-N-G found in C thermocellum p- glucosidase A (Grabnitz et a l , 1991), and indeed there appears to be a family of
glycosidases with Glu in the active site (Wacker et a l , 1992).
The cleaved pro-portion (residues 20 - 866) comprising the first two repeat units
has no known function and may be rapidly degraded. Although it was initially suggested
that this portion might be a soluble, cytosolic 8-glycosidase because of its homology with
the mature lactase polypeptide (Mantei et a l , 1988), no evidence has been obtained to support this. Attempts to raise antibodies to this pro-portion have not been successful,
resulting in the opinion that it was rapidly degraded. Indeed, the active site motif was
shown not to be conseiwed in the first two repeats which foim the pro-portion o f the
molecule, which suggested that this portion does not function as a glycosidase (this
region was not present at all in repeat one, and the sequence in repeat two was Y-L-A-G-
N-G)(Wacker et a l, 1992). Naim et a l, (1991) demonstrated that the full length lactase cDNA when transfected into COS cells in an appropriate vector produced an enzyme with
the same activity per mole as the mature form of lactase, either isolated from the brush
border or produced by trypsin treatment of the transfected pro-LPH, when either lactase
or phlorizin was used as substrate. Expression studies using partial and complete cDNA
sequences in an appropriate vector transfected into COS cells demonstrated that the pro
sequence is required for insertion of the protein in the plasma membrane (Oberholzer et a l , 1993). Thus the first two repeats of the lactase sequence appear to have no
independent physiological function and are most probably only involved in the targeting
of the mature LPH polypeptide to the brush border.
Initial sequence analysis had suggested that LPH would have a C-in N-out
portion which is only long enough to cross the lipid bilayer once (Mantei et aL, 1988). Wacker et al. 1992, have shown using the hydrophobic photolabel TID that the mature lactase polypeptide is indeed anchored into the membrane by this region (amino acid
1883-1901 in rabbit) and that the C-teiminal region of the polypeptide is cytosolic. This
is unlike the situation in sucrase-isomaltase where the protein is anchored via an amino-
terminal non-cleaved signal (Wacker et at., 1992 and section 1.1.4.1) or in trehalase which is attached to the membrane via a phosphatidylinositol anchor (Ruf et a i , 1990 and section 1.1.4.3).
1.1.8.2. C haracteristics and tissue specificity o f the m essenger RNA.
Northern blot analysis of the lactase message using the rabbit cDNA probe
revealed a message of approximately 6 kb in the small intestine of rabbits which was not
present in the lung, kidney or liver (Mantei et al., 1988). Studies in rats have shown that the lactase mRNA is found in the small intestine, but not in heart, liver, lung, kidney,
testes, spleen or brain, the message was assessed to be 6.8 kb (Buller etaL , 1990).
In humans, SI-mapping and primer extension experiments localised the 5' end of
the LPH mRNA to 11 nucleotides upstream of the start of translation (ATG) in both
persistent and non-persistent individuals (Boll et al., 1991). Indeed, no qualitative difference was found between the mRNA in non-persistent or persistent individuals by
PCR, sequencing or SI-protection assays nor was any difference seen in the band pattern
on Southern blot analysis (Boll et al., 1991). However the SI protection assays only studied the region 5' to nucleotide 5456 such that differences in 3' processing of the
1.1.8.3. Q uantitation o f the levels o f lactase m essenger RNA and
determ ination o f its cellular localisation com pared with the localisation o f lactase protein and activity.
Northern blots, slot blots and SI mapping have been used to attempt to quantify
the levels of lactase mRNA (LPH mRNA) at different stages of development and at
different positions in the intestine. The method of expressing the results obtained varies;
LPH mRNA may be quantified with respect to the amount of protein produced, total
mRNA, or relative to another intestinal or housekeeping mRNA.
The cellular localisation of the lactase message has been investigated by the
technique of in situ hybridisation to tissue sections. The majority of experiments have
been performed using rat intestine, although some work has also been done in rabbit, pig
and using human tissue from persistent and non-persistent individuals. In all cases the
lactase mRNA was found in the enterocytic cells of the villus and not in the crypt (for
example in rat (Rings et al., 1992b), in rabbit (Freeman et al., 1993) and human (Maiuri et a l, 1994)). This is in agreement with the localisation of lactase activity along the ciypt-villus axis (see section 1.1.5.1) and with immunohistological detection of the LPH
protein (Maiuri et a l, 1992 - rabbit and rat, 1993a - human).
I shall review the studies on each species separately, in the following sections.
1.1.8.3.1. Stu dies using ra b b it in testin e.
In rabbits there is a decline in the level of lactase activity in the intestine at the time
of weaning. Sebastio et al. (1989) observed a parallel decrease in LPH mRNA level at this time. However, these authors also demonstrated a higher level of LPH mRNA in 5
adult animals than in post-weaned animals. In their study the level of LPH mRNA was