HELGOLANDER MEERESUNTERSUCHUNGEN Helgol~nder Meeresunters. 49, 45-56 (1995)
The contribution of marine b i o l o g y to b i o m e d i c a l
research: past, present, future
E. K i n n e - S a f f r a n 1'2 & R. K. H. K i n n e 1'2
1 Max-Planck-Institut ffir molekulare Physiologie, Abteilung Fpithelphysiologie; Postfach 1026 64, D-44026 Dortmund, Germany
and
2Mount Desert Island Biological Laboratory, Salsbury Cove, Maine, USA
"Ask the fish of the sea, and they will declare unto thee" (Job 12:8)
I N T R O D U C T I O N
This c o n t r i b u t i o n focusses on the role m a r i n e biology - or m o r e p r e c i s e l y m a r i n e species or m a r i n e m o d e l systems - h a v e p l a y e d in the e l u c i d a t i o n of basic p h y s i o l o g i c a l processes. M a n y of t h e s e h a v e t u r n e d out to b e of utmost s i g n i f i c a n c e in u n d e r s t a n d i n g the function of the h u m a n b o d y in h e a l t h a n d d i s e a s e or in d e v e l o p i n g t h e r a p e u t i c a l m e a n s to cure h u m a n maladies. T h e particular u s e f u l n e s s of m a r i n e m o d e l s in a v a r i e t y of t h e s e s e m i n a l d i s c o v e r i e s can b e t r a c e d b a c k to t h r e e m a i n reasons: (a) A b u n d a n c e of species, size of b i o l o g i c a l m o d e l system, and r e a d y accessibility l e a d to an e a s e of e x p e r i m e n t a t i o n . (b) In s e v e r a l m a r i n e species, that r e p r e s e n t early steps in m a m m a l i a n evolution, o r g a n structure is often q u i t e simple and o r g a n function is h i g h l y specialized. Such " u n i f u n c t i o n a l i t y " contrasts to the "multifunctionality" usually f o u n d in m a m m a - lian a n d h u m a n tissues and facilitates the i n v e s t i g a t i o n of a particular function in a defined, h o m o g e n e o u s cell population. (c) Most importantly, it has b e c o m e clear, m a i n l y t h r o u g h the r e c e n t a d v a n c e s in m o l e c u l a r biology a n d cell biology, that b i o d i v e r s i t y in cellular function does not r e q u i r e an i n c o m p r e h e n s i b l e n u m b e r of functionally different units: it can b e r e d u c e d to the e x i s t e n c e of a limited n u m b e r of families of closely r e l a t e d m o l e c u l e s that are e m p l o y e d by n a t u r e to p e r f o r m basic cellular functions (Kinne. 1991a). T h e s e similarities allow us to d r a w conclusions from findings on m a r i n e o r g a n i s m s as to the function of a h u m a n o r g a n w i t h e v e n m o r e certainty.
In the following, mainly contributions of m a r i n e m o d e l s to r e n a l p h y s i o l o g y and p a t h o p h y s i o l o g y will b e h i g h l i g h t e d , b u t it should be e m p h a s i z e d t h a t t h e r o l e of m a r i n e models was e q u a l l y i m p o r t a n t in o t h e r areas of cell research. In the a r e a of o o g e n e s i s , s p e r m a t o g e n e s i s , cytokinesis, and r e p r o d u c t i o n , n u m e r o u s studies on t h e e g g s of s e a urchins, s a n d dollars, and snails h a v e laid a firm basis for our u n d e r s t a n d i n g of cell division a n d its t e m p o r a l and spatial o r g a n i z a t i o n (Rappaport, 1991). E l a s m o b r a n c h testes h a v e recently b e e n d i s c o v e r e d to p r o v i d e an i d e a l m o d e l system for the studies of different p h a s e s of s p e r m a t o g e n e s i s a n d the v i v i p a r o u s dogfish is a suitable m o d e l for
46 E. K i n n e - S a f f r a n & R. K. H. K i n n e
i n v e s t i g a t i o n s of the h o r m o n a l r e g u l a t i o n of r e p r o d u c t i o n (Callard, 1991; Koob & Callard, 1991). In n e r v e physiology, the g i a n t axon of the squid was - a n d c o n t i n u e s to be - one of the prime m o d e l systems i n which the properties of n u m e r o u s ion c h a n n e l s a n d ion p u m p s have b e e n characterized a n d their role in n e r v e c o n d u c t i o n i d e n t i f i e d (Boron & Knakal, 1992; H o d g k i n & Huxley, 1952). Also the Na-K-ATPase, the p r i m a r y p u m p m a i n t a i n i n g i n t r a c e l l u l a r ion homeostasis a n d cell volume, was first d e s c r i b e d b y Skou in the leg n e r v e s of crabs (Skou, 1957), t h e r e b y p r o v i d i n g the first link of a n A T P - c o n s u m i n g cellular reaction to the translocation of i n o r g a n i c electrolytes across cell m e m b r a n e s (Skou, 1989). This e n z y m e w a s later f o u n d b y S c h a t z m a n n (1967) to b e i n h i b i t e d b y "cardiac glycosides" which are u s e d for the t r e a t m e n t of heart failure. U n t i l today, the s o d i u m chloride-secreting rectal g l a n d of the shark is one of the richest sources of this e n z y m e for b i o c h e m i c a l a n d biophysical studies (Medzihradsky et al., 1967). T h e rectal g l a n d also, very early on, p l a y e d a n i m p o r t a n t role in e l u c i d a t i n g the m e c h a n i s m s of h o r m o n a l r e g u l a t i o n of salt transport a n d i n t r a c e l l u l a r s i g n a l l i n g - a n area still p u r s u e d vigorously u s i n g this o r g a n as m o d e l system (Schofield et al., 1991).
MARINE BIOLOGY AND RENAL PHYSIOLOGY
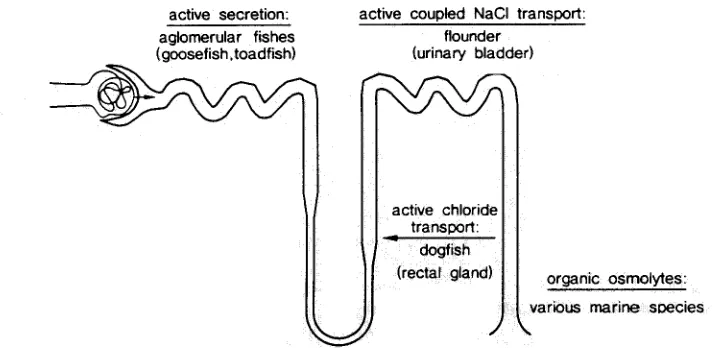
In Figure 1, a s c h e m e of a m a m m a l i a n n e p h r o n is r e p r e s e n t e d with its proximal tubule, H e n l e ' s loop, the distal t u b u l e a n d the collecting duct. In all t h e s e parts of the k i d n e y , studies on m a r i n e o r g a n i s m s have c o n t r i b u t e d essential i n f o r m a t i o n on the function of t h e s e s e g m e n t s a n d the cellular a n d m o l e c u l a r basis for their function.
For the proximal tubule, in 1923, the q u e s t i o n was solved w h e t h e r m a m m a l i a n k i d n e y s h a v e the capability to excrete c o m p o u n d s from the blood into the p r i m a r y urine (Marshall & Vickers, 1923) i n addition to the - at that time - w i d e l y - a c c e p t e d functions of filtration in the g l o m e r u l u m a n d r e a b s o r p t i o n a l o n g the t u b u l e (Cushny, 1917). Definite proof of secretory processes i n this s e g m e n t could b e o b t a i n e d b y the use of t h e goosefish
active secretion: aglomerular fishes (goosefish,toadfish)
active coupled NaCI transport: flounder
(urinary bladder)
active chloride transl~ort: - dogfish
(rectal gland) organic osmolytes:
various marine species
\
Fig. 1. Schematic representation of a mammalian (human) nephron indicating some of the main segments where marine biology has considerably contributed to elucidating the cellular a n d
M a r i n e biology a n d b i o m e d i c a l r e s e a r c h 47
(Lophius piscatorius),
i.e. an a g l o m e r u l a r fish, in w h i c h no filtration occurs, a n d substan- ces transferred from the blood to the urine must h a v e p a s s e d the r e n a l cell in a secretory direction (Marshall & Grafflin, 1928). This process w a s first f o u n d for a variety of organic dyes and later l e d to the d e v e l o p m e n t of contrast m e d i a to trace the urinary tract, or to m e a s u r e r e n a l b l o o d flow. This route is still e m p l o y e d for the t a r g e t i n g of antibiotics in r e n a l infection or of diuretics to their i n t r a t u b u l a r site of action. T h e cellular m e c h a n i s m u n d e r l y i n g t u b u l a r secretion w a s also first u n v e i l e d using fish models, such as the flounder, (Forster & Taggart, 1950; Kinter, 1966). In F i g u r e 2, studies are s h o w n in w h i c hFig 2. 3H-chlorphenol red autoradiograph of proximal tubules dissected from the kidney of the winter flounder
(Pseudopleuronectes americanus)
and incubated Eor 60 rain at 20~ in 10 ~M tritiated chlorphenol red. Individual tubules are shown both in cross and longitudinal sections m a low-magnification dark-field photomicrograph (autographic silver grams appear as white dots). Forfurther details see Kinter, 1975 (reprinted with kind permissionl
48 E. K i n n e - S a f f r a n & R. K. H. Kinne
Lumen
H*,
I
'H§
,,~
%'S- . . .
organic cations
Lumen
~/S
PAH.~
Blood
NG~,. I IA-
pA~ -~
weak organic acids
NQ§
S0/2-
sulfate
Fig. 3. Transport scheme for the secretion of the weak organic acid p-aminohippurate (PAH) across a proximal tubular cell (upper right panel). Uptake from blood into the cell involves indirect coupling to a sodium-A- cotransport system. (A- stands for glutarate or other dicarboxylic acids). Transfer across the luminal membrane involves an electrogenic transporter driven by the electrical potential difference across the cell membrane. The two other schemes show additional kinds of indirect coupling of sodium cotransport transport systems in the renal secretion of sulfate in winter flounder and organic cations (S) in mammals. For further information see Kinne. 1988a (reprinted with kind
permission)
intracellularly a c c u m u l a t e d g l u t a r a t e w i t h a w e a k o r g a n i c acid, comprises t h e s e q u e n c e of e v e n t s that u l t i m a t e l y l e a d to the i n t r a c e l l u l a r a c c u m u l a t i o n of the w e a k o r g a n i c acid. S u c h a m e c h a n i s m has p r o v e d to b e o p e r a t i n g also for e x a m p l e in the e x c r e t i o n of uric acid in c r u s t a c e a n s (A. N i e s et al., unpubl, obs.) as well as in m a m m a l s (Maxild et al.. 1981).
M a r i n e b i o l o g y a n d b i o m e d i c a l r e s e a r c h 49
Blood Lumen
No-K- i
ATPose K+
T A L H Cell
\
__.2
2CI - /
/ K +
K /
/ No ~"
~
K § NoCI-KCIcotronsport
2Ci- .~- Ba + +
-- NO +
B l o o d Rectal Gland C e l l L u m e n
\
No-K- "/ ~ 1 - 1 - ~ N ~ 2 4 7 ATPose j K+ ~ , ~
NoCI-KCI i
N~ § ~x/~
K + _ ~cotronsport fl 2CI
2No + - - - ~ /
J
2CI -,, \
Fig. 4. Schematic representation of transcellular active chloride secretion in rectal gland cells of elasmobranchs and active chloride reabsorption in the thick ascending limb of Henle's loop in
mammalians (modified after Epstein & Silva, 1985, with kind permission)
d o c u m e n t e d in F i g u r e 5 [ K i n n e & K i n n e - S a f f r a n 1979). T h i s l i n k l e d to a w o r k i n g m o d e l for a c t i v e c h l o r i d e t r a n s p o r t i n t h e m a m m a l i a n t h i c k a s c e n d i n g l i m b - a n d t h e o p e r a t i o n of all t h e a b o v e m e n t i o n e d transport s y s t e m s c o u l d a l s o b e d e m o n s t r a t e d in t h e k i d n e y ( G r e g e r , 1985). It w a s further s h o w n t h a t t h e s e t r a n s p o r t s y s t e m s h a v e s i m i l a r p r o p e r t i e s - as d e p i c t e d for t h e N a - K - 2 C 1 c o t r a n s p o r t e r in T a b l e 1 ( K i n n e , 1 9 8 8 a ) - b u t t h a t their c e l l u l a r l o c a l i z a t i o n is d i f f e r e n t , in o r d e r to e n a b l e t h e m a m m a l i a n c e l l s to r e a b s o r b c h l o r i d e r a t h e r t h a n to s e c r e t e c h l o r i d e as t h e r e c t a l g l a n d c e l l s do. To this e n d a N a - K - 2CI c o t r a n s p o r t e r a n d a p o t a s s i u m c h a n n e l a r e k n o w n to b e t r a n s f e r r e d i n t o the l u m i n a l c e l l m e m b r a n e , w h e r e a s a c h l o r i d e c h a n n e l is in t h e c o n t r a l u m i n a l m e m b r a n e . S u c h t r a n s p o s i t i o n r a i s e s i n t e r e s t i n g q u e s t i o n s on t h e m o l e c u l a r i d e n t i t y of t h e t r a n s p o r t s y s t e m s a n d t h e n a t u r e of t h e s i g n a l s c o n t r o l l i n g their i n t r a c e l l u l a r s o r t i n g a n d t a r g e t i n g ( S i m o n s & Fuller, 1985) - q u e s t i o n s c u r r e n t l y b e i n g i n v e s t i g a t e d - a g a i n u s l n q m a r i n e m o d e l s y s t e m s .
A n o t h e r g r o u p of d i u r e t i c s t h e t h i o z i a z i d e s , are k n o w n to i n h i b i t salt t r a n s p o r t in the l a t e d i s t a l t u b u l e [ s e e F i g u r e 6; [ E l l i s o n e t al,, 1987]), a s e g m e n t of v e r y h i g h cell h e t e r o g e n e i t y i n t h e m a m m a l s . H e r e t h e d i s c o v e r y of a s i m i l a r t r a n s p o r t s y s t e m in f l o u n d e r u r i n a r y b l a d d e r ( T a b l e 2; [ S t o k e s e t al,, 1984]), a r a t h e r s i m p l e e p i t h e l i u m , will u n d o u b t e d l y l e a d to a d e t a i l e d c h a r a c t e r i z a t i o n of t h e t r a n s p o r t s y s t e m a n d to a m u c h b e t t e r u n d e r s t a n d i n g of t h e m e c h a n i s m of a c t i o n o f t h e s e d r u g s .
G, Table 1. Properties of the Na-K-2 C1 cotransporter in secretory and absorptive epithelia (Reprinted with permission from Kinne, 1988a, Pergamon Press Ltdl Rabbit TALH Rectal gland Number per cell Turnover rate Sodium binding site
Affinity Specificity
Potassium
binding
site
Affinity Specificity
Chloride
binding
site
1
Affinity Specificity
Chloride
binding
site
2
Affinity Specificity
Interaction with loop diuretics Affinity (bumetanide) Specificity Apparent molecular weight {radiation inactivation} 5 X 104 6000/S 1.3 mM Na > Li>> NH4 >> K 0.3 mM K > NH4 > Cs >> Na >> choline 1.0 mM Br = CI >> NOa = SCN > 15raM Br > Cl > NO3 = SCN 2.5 x 10 -6 M bumetanide > piretanide > furosemide 80 000-90 000 17 • 104 1000/s 4,3 mM Na > Li = NH4 15 mM K-> NH4 = K = Rb = choline > Cs overall 75 mM Br = CI>>NO3 Br > CI > NO3 0.5 x 10 -5 M bumetanide > piretanide > furosemide 43 000-50 0(30 {affinity chromatography)
M a r i n e b i o l o g y a n d b i o m e d i c a l r e s e a r c h 51
0 U
0
0
0
0
"-- oO c"
c ~ E r~ o
0 C ~
>, E
rn ~ LL CO
c" O
fL
E (D
o
LL
t ~
T M! !
Fig. 5. Effects of v a r i o u s "loop d i u r e t i c s " o n t h e r a t e of fluid s e c r e t i o n (i.e: s o d i u m chloride) in "in situ p e r f u s e d " rectal g l a n d s of S c i l l i o r r h m u s c a n i c u l u s (L.). Loop d i u r e t i c s are k n o w n to i n h i b i t c h l o r i d e r e a b s o r p t i o n in h u m a n k i d n e y at t h e t h i c k a s c e n d i n g l i m b of H e n t e ' s loop. S e c r e t i o n at v a r i o u s c o n c e n t r a t i o n s of t h e d i u r e t i c s is g i v e n in p e r c e n t of control. For f u r t h e r details s e e K i n n e & K i n n e -
Saffran, 1979 ( r e p r i n t e d w i t h k i n d p e r m i s s i o n )
":1= I=
C
E
0
E
i z
600
400
200
.200
Sodium transport
Chloride transport
800
\
\
. . . . ~ . . o
~ o
r
E
O
u 0
-200
P < .001
\
%
P < . 0 0 1
-400 -4OO
A I F A I F * C T Z A I F A I F + C T Z
Fig. 6. Effect of c h l o r o t h i a z i d e (CTZ) o n s a l t r e a b s o r p u o n m d i s t a l t u b u l e s of r a b b i t k i d n e y . N o t e t h a t s o d i u m a n d c h l o r i d e t r a n s p o r t a r e i n h i b i t e d to t h e s a m e e x t e n t s u g g e s t i n g t h e p r e s e n c e of a n e l e c t r o n e u t r a l NaC1 c o t r a n s p o r t in t h i s r e n a l s e g m e n t . For f u r t h e r i n f o r m a t i o n s e e E l l i s o n et al. 1987
52 E. K i n n e - S a f f r a n & R. K. H. K i n n e
Table 2. Effect of hydrochlerothiazide on simultaneously determined sodium and chloride tracer fluxes in the urinary bladder of the winter flounder
{Pseudopleuronectes americanus).
Mucosa-to- serosa denotes the fluxes m e a s u r e d w h e n tracer ions w e r e present at the surface r e p r e s e n t i n g the lumen of the bladder; serosa-to-mucosa depicts fluxes w h e n tracer ions were p r e s e n t at the outside of the bladder. Net fluxes r e p r e s e n t the difference b e t w e e n the fluxes found in the mucosa-to-serosa direction and those observed in the serosa-to-mucosa direction. Note the similarity of sodium and chloride net fluxes and the parallel inhibition by hydrochlorothiazide. Modified after Stokes et al.,1984, with kind permission
Flux
Mucosa-to-serosa (n = 8) Serosa-to-mucosa (n = 7) N e t fluxes
JNa Jcl JNa Jcl JNa Jcl
~M/cm 2 9 h ~M/cm 2 - h
Control 1.70 2.45 0.52 1.34 1.18 1.11
• 0.25 • 0.30 • 0.07 • 0.12
HCTZ 0.40 0.69 0,23 0.29 0.17 0.40
(0.1 mM) _ 0.05 • 0.24 • 0.02 • 0.04
P < 0.005 < 0.005 < 0.002 < 0.001 - -
JNd ~ sodium flux; Jcl = chloride flux
o s m o l y t e s in t h e v o l u m e r e g u l a t i o n of t h e s e c e l l s ( G a r c i a - P e r e z & Burg, 1991). D u r i n g t h e f o r m a t i o n of c o n c e n t r a t e d u r i n e , t h e s e c e l l s a r e e x p o s e d to s a l i n i t i e s w h i c h a r e s i m i l a r to or e x c e e d t h o s e e n c o u n t e r e d b y m a r i n e o r g a n i s m s . T h e s e o r g a n i c o s m o l y t e s h a v e for a l o n g t i m e a l r e a d y b e e n i d e n t i f i e d i n m a r i n e o r g a m s m s / s e e T a b l e 3: [ Y a n c e y e t al., 1982]) a n d a r e a l s o f o u n d in t h e m a m m a l i a n p a p i l l a ( T a b l e 4). T h e r e g u l a t i o n of t h e i r i n t r a c e l l u - l a r c o n c e n t r a t i o n s a c c o r d i n g to t h e e x t e r n a l o s m o l a r i t y h a s b e e n e x t e n s i v e l y s t u d i e d for e x a m p l e i n s k a t e e r y t h r o c y t e s ( G o l d s t e i n & Brill 19911. A g a i n n u m e r o u s s i m i l a r i t i e s h a v e b e c o m e a p p a r e n t , a n d it is to b e e x p e c t e d t h a t t h e c r o s s - t a l k b e t w e e n d i s c i p l i n e s - a s m t h e o t h e r e x a m p l e s m e n t i o n e d a b o v e - will y i e l d fruitful a n d i m p o r t a n t i n s i g h t s i n t o g e n e r a l p h y s i o l o g i c a l m e c h a n i s m s .
C O N C L U D I N G R E M A R K S
In t h e f u t u r e , m a r i n e b i o l o g i c a l m o d e l s w i l l c o n t i n u e to f o s t e r p r o g r e s s in t h e u n d e r s t a n d i n g of m o l e c u l a r , c e l l u l a r a n d s y s t e m i c p r o c e s s e s in b i o m e d i c i n e . U n d o u b - t e d l y , a m a j o r i m p a c t will b e m a d e b y s t u d i e s o n t h e m o l e c u l a r b i o l o g y of b a s i c p h y s i o l o g i c a l a n d p a t h o p h y s i o l o g i c a l e v e n t s . A t t h e s a m e t i m e h o w e v e r , a n e q u a l l y s t r o n g e m p h a s i s s h o u l d b e p l a c e d o n i n t e g r a t i v e a s p e c t s in m a r i n e b i o l o g y a n d b i o m e d i c i n e alike: o n l y t h e i n t e g r a t i o n of k n o w l e d g e o b t a i n e d a t v a r i o u s l e v e l s of c o m p l e x i t y a n d i n v a r i o u s a r e a s of r e s e a r c h w i l l l e a d to a t h o r o u g h u n d e r s t a n d i n g of life o n t h i s p l a n e t a n d to t h e d e v e l o p m e n t of s t r a t e g i e s to p r o t e c t it.
M a r i n e b i o l o g y a n d b i o m e d i c a l r e s e a r c h 53 Table 3. Organic osmolytes in marine species. T a k e n from Yancey et al., 1982; copyright 1982 by the
AAAS, with kind permission. For references see Yancey et al. 1982
Osmolyte system Principal osmolytes
(occurrences)
C y a n o b a c t e r i a
Synechococcus sp. F u n g i
Saccharomyces rouxii Asteromyces cruciatus
Lichens
Lichina p y g m e a e
Unicellular algae
Dunaliella spp.
Chlorella pyrenoidosa Ochromonas maihamensis
Multicellular algae
Fucus spp. Vascular plants
Gossypium hirsutum L.
Insects ~freeze-tolerant or -resistant)
Eurosta solidaginis (Diptera)
Bracon cephi (Hymenopteral C r u s t a c e a n s
Artemia salina (emerging larvae/ Vertebrates
A. Polyhydric a l c o h o l s - p o l y o l s
glucosylglycerol arabitol
arabitol, glycerol, mannitol m a n n o s i d o m a n n i t o l glycerol
sucrose isofloridoside mannitol
glucose, fructose sucrose glycerol, sorbito] glycerol
glycerol, trehalose
Eubacteria
Klebsiella aerogenes Salmonella oranienburg Streptococcus faecalis
Protozoa
1Vfiemiensis avidus
Vascular plants
Spartina townsendii Atriplex spongzosa Aster tripolium
M e s e m b r y a n t h e m u m nodiflorum
I n v e r t e b r a t e s
All phyla of m a r i n e v e r t e b r a t e s
Balanus nubilus (barnacle)
Eriocheir sinensis (crab)
Parastichopus sp. (echinoderm)
Sepia officinalis (mollusk)
Cyclostomes
A/fyxine glutinosa (hagfish)
A m p h i b i a
Bufo marmus
glutamic acid proline glutamic acid proline y-aminobutyric acid, prolin~ glycine, alanine, proline b e t a i n e
b e t a i n e proline proline
v a n o u s amino acids various amino acids various amino acids various amino acids various amino acids
v a n o u s amino acids
Hyla versicola glycerol
54
Osmolyte system (occurrences)
E. K i n n e - S a f f f a n & R. K. H. K i n n e Table 3 (continued)
Principal osmolytes
C. Urea and methylamines
Cartilaginous fishes {elasmobranchs) Vertebrates
Squalus acanthias (dogfish) Dasyatis americana
(ray)
Raja erinacea
(ray)
urea, trimethylamine-N- oxide urea, t n m e t h y l a m i n e - N-oxide urea, amino acids
D. Urea: estivating forms
Mollusks
Bulimulus dealbatus
Lungfishes: African a n d South A m e r i c a n A m p h i b i a n s
Scaphiopus couchi (spadefoot toad)
E. Inorganic ions
A r c h a e b a c t e r i a
Halobacterium spp. K-
Table 4 0 s m o l y t e s in renal i n n e r medulla of antidiuretic animals. Units are m m o l / k g protein or m m o l / k g wet w e i g h t (in brackets). GPC = glycerophosphorylcholine. Taken from Garcia-Perez &
Burg, 1991. with kind permission. For references see Garcia-Perez & Burg, 1991
Species Urea Sodium Sorbitol Inositot GPC Betaine
Rabbit [7] [ 11] [ 13] [56]
[269] [21] [21] [35]
[346] [279] [80] [16] [41] [42]
1.017 1.890 221 97 195 235
[21]
[20l
Rat [49]
[380] [248] 854 331 2.498 132
4.405 178 517 214
145 214 443
Vole [349] [I71] [26] [12] [43] [19]
Deer m o u s e [695] [278] [19] [14] [65] [21]
Pocket m o u s e [1.129] [350] [53] [14] [71] [50]
Sheep
[54]Dog [67]
[376]
[304]
[14]
M a r i n e b i o l o g y a n d b i o m e d i c a l r e s e a r c h 55
L I T E R A T U R E C I T E D
Boron, W. F. & Knakal, R. C., 1992. N a l + L d e p e n d e n t C1-HCO3 e x c h a n g e in the s q u i d axon. - J . gen. Physiol. 99, 817-837.
Callard, G. V., 1991. Reproduction in m a l e e l a s m o b r a n c h fishes. - C o m p . Physiol. 10, 104-154. C u s h n y , A. R., 1917. T h e secretion of urine. L o n g m a n s G r e e n , London, 288 pp.
Ellison, D. H., Vel~zquez, H. & Wright, F. S., 1987. T h i a z i d e - s e n s i t i v e s o d i u m chloride c o t r a n s p o r t in early distal t u b u l e . - Am. J. Physiol. 253, F546-F554.
Epstein, F. H. & Silva, P., 1985. Na-K-C1 c o t r a n s p o r t in c h l o r i d e - t r a n s p o r t i n g epithelia. - A n n . N. Y. Acad. Sci. 456, 187-197.
Eveloff, J., Kinne, R. & Kinter, W., 1979. p - A m i n o h i p p u r i c acid t r a n s p o r t into b r u s h b o r d e r vesicles isolated f r o m f l o u n d e r kidney. - Am. J. Physiol. 237, F291-F298.
Forster, R. P. & T a g g a r t , J. V., 1950. Use of isolated r e n a l t u b u l e s for the e x a m i n a t i o n of m e t a b o l i c p r o c e s s e s a s s o c i a t e d with active cellular t r a n s p o r t . - J. cell. comp. Physiol. 36, 251-270. Garcia-Perez, A. & Burg, M.B., 1991. Renal m e d u l l a r y o r g a n i c osmolytes. - Physiol. Rev. 71,
1081-1115.
Goldstein, L. & Brill S. R., 1991. V o l u m e - a c t i v a t e d t a u r i n e efflux from skate erythrocytes: p o s s i b l e b a n d 3 i n v o l v e m e n t . - Am. J. Physiol. 260, R1014-R1020.
G r e g e r , R., 1985. Ion t r a n s p o r t m e c h a n i s m s in thick a s c e n d i n g limb of H e n l e ' s loop of m a m m a l i a n n e p h r o n . - Physiol. Rev. 65. 760-797.
H o d g k i n . A. L. & Huxley, A. F.. 1952. A q u a n t i t a t i v e description of m e m b r a n e c u r r e n t a n d its application to c o n d u c t i o n and excitation in nerve. - J Physiol. I 17. 500-544.
K i n n e R. K. H.. 1988a. S o d i u m c o t r a n s p o r t s y s t e m s in epithelial secretion. - C o m p . Biochem Physiol 90A, 721-726.
Kinne, R., 1988b T h e N a / K / C l - c o t r a n s p o r t e r in epithelia: a p e r s o n a l r e m i n i s c e n c e . - Bull. Mt D e s e r t [sl. biol. Lab. 51-56.
Kinne. R. K.H. I991a. F r o m diversity to similarity in biological transporL - I s s u e s Biomed. 15, 69-94 Kinne R . K . H . . 1991b. Molecular p r o p e r t i e s of the Na-K-C1 c o t r a n s p o r t e r . In: N e p h r o l o g y . Ed. by M
Hatano. S p r i n g e r , Tokyo, 2. 1095-1102.
Kinne R. & Kinne-Saffran. E.. 1979. Effect of 'loop diuretics' on the salt secretion in s h a r k rectal gland. - Mar. Ecol. Prog. Ser. 1. 129-132.
Kinter. W. B.. 1966. C h l o r p h e n o l red influx a n d efflux: m i c r o s p e c t r o p h o t o m e t r y of f l o u n d e r k i d n e y t u b u l e s . - Am. J. Physiol. 211, 1152-1164.
Kinter. W. B. 1975. Structure a n d f u n c t i o n of r e n a l t u b u l e s isolated from fish kidneys. - Fortschr. ZooI. 23. 223-231.
Koob T. J. & C a l l a r d [. P., 1991. R e p r o d u c t i o n in f e m a l e e l a s m o b r a n c h s , - C o m p . Physiol. I0, 155-209.
Marshall. E. K. & Grafflin. A. L. 1928. T h e s t r u c t u r e a n d f u n c t i o n of the k i d n e y of L o p h i u s piscatorius. - Bull. J o h n s H o p k i n s Hosp. 43. 205-235.
Marshall. E. K. & Vickers J. L.. 1923. T h e m e c h a n i s m of the elimination of p h e n o s u l p h o n e o p h t h a - Iein b y the k i d n e y - a proof of s e c r e t i o n b y the c o n v o l u t e d t u b u l e s . - t~ull. J o h n s H o p k i n s Hosp. 34. 1-16.
Maxild. J., Moiler, J. V. & Sheikh, M. I.. 1981. A n e n e r g y - d e p e n d e n t , s o d i u m - i n d e p e n d e n t c o m p o - n e n t of active p - a m i n o h i p p u r a t e t r a n s p o r t in r a b b i t r e n a l cortex. - J. Physiol., Lond. 310. 2 7 3 - 2 8 3
M e d z i h r a d s k y , F., K.line M. H. & H o k i n L. E., 1967. S t u d i e s on the c h a r a c t e r i z a t i o n of t h e s o d i u m - p o t a s s i u m t r a n s p o r t a d e n o s i n e t r i p h o s p h a t a s e . I.: Solubilization, stabilization, a n d e s t i m a t i o n of a p p a r e n t m o l e c u l a r weight. - A r c h s Biochem. Biophys. 121, 311-316.
Pritchard. J. B.. 1990. Rat r e n a l cortical slices d e m o n s t r a t e p - a m i n o h i p p u r a t e / g l u t a r a t e e x c h a n g e a n d s o d i u m / g l u t a r a t e c o u p l e d p - a m i n o h i p p u r a t e t r a n s p o r t . - ft. PharmacoL exp. Thor. 255. 969-975
R a p p a p o r t . R., 1991. Cytokinesis. - C o m p . Physiol. I0. 1-36.
S c h a t z m a n n , H. J., 1967. Effect of cardiac g l y c o s i d e s o n active N a - K - t r a n s p o r t . - P r o t o p l a s m a 63. 136-142.
56 E. K i n n e - S a f f r a n & R. K. H~ K i n n e
S h i m a d a , H., M o e w e s , B. & Burckhardt, G., 1987. Indirect c o u p l i n g to N a + of p - a m i n o h i p p u r i c acid u p t a k e into rat r e n a l b a s o l a t e r a l m e m b r a n e vesicles. - Am. J. Physiol. 253, F 7 9 5 - F 8 0 1 . Simons, K. & Fuller, S.D., 1985. Cell s u r f a c e polarity in epithelia. - A. Rev. Cell Biol. I, 243-288~ Skou, J. C., 1957. T h e i n f l u e n c e of s o m e cations on the a d e n o s i n e t r i p h o s p h a t a s e from p e r i p h e r a l
nerves. - Biochim. b i o p h y s . Acta 23, 394-401.
Skou, J. C., 1989. T h e identification of the s o d i u m - p u m p as the m e m b r a n e - b o u n d N a §
a c o m m e n t a r y on ' T h e i n f l u e n c e of s o m e cations on an a d e n o s i n e t r i p h o s p h a t a s e from p e r i p h e r a l n e r v e s ' . - Biochim. b i o p h y s . Acta I000, 435-438.
Stokes, J. B., Lee, I. & D'Amico, M., 1984. S o d i u m chloride a b s o r p t i o n by the u r i n a r y b l a d d e r of the w i n t e r flounder. - J. clin. Invest. 74, 7-16.