ESTIMATION OF VARIANCE COMPONENTS OF INTERNODE MIMULUS GUTTATUS
LENGTH IN A CLONED POPULATION OF
W. J. LIBBY2
Department of Genetics, University of California, Berkeley, California
Received February 15, 1962
a plant is vegetatively reproduced, or cloned, so that the same genotype can I;evelop in more than one environment, differences in a given character be- tween the individuals of the clone reflect the effect of the different environments on the expression of the character. If a population of plants is cloned and ran- domly grown in a nonuniform environment, it is possible to estimate environ- mental and total genetic components of variation by evaluating the variation within and between clones.
BURTON and DEVANE (1953) computed environmental and total genetic com-
ponents of variance, and broad-sense heritability, for seven characters using replicated clones of several ecotypes of tall fesque. COOPER (1959) further refined the technique by using parent-off spring regression to estimate narrow-sense (additive) heritability of Lolium ear emergence, and by comparing broad-sense and narrow-sense heritabilities for evidence of nonadditive effects. It is the pur- pose of this study to further explore variance component estimation using clones, and to compare such estimates to independent estimates using more conventional designs.
MATERIALS A N D METHODS
Mimulus guttatus Fisch., the yellow monkey flower commonly found growing on stream banks and seeps in western North America, was chosen for these ex- periments. I t is an outcrossing perennial which clones in nature, and has proven to be an excellent research organism. The logarithm of the length of the six inter- nodes above the first flowering node was the quantitative character chosen for study (Figure 1). This character develops late enough so that differences in the condition of the vegetative parts used for cloning did not strongly influence the development of the character (LIBBY and JUND 1962). Analysis of a 105-plant population sample indicated that the logarithm of this measurement is normally distributed ( x 2 = 3.35, x 2 95 = 15.51 : 8 df), has negligible skewness (t = 0.06,
t 9 5 = 1.96), and slight but nonsignificant kurtosis (t = -0.81, t.95 = -1.96), while analyses of other samples indicated that the variance was uncorrelated with the mean level.
1 Part of a thesis submitted to the Graduate Division, University of California, in partial 2 Present address: School of Forestry, University of California, Berkeley.
fulfillment of the requirements for the degree of Doctor of Philosophy i n Genetics.
p
-i=
FIGURE 1 .-A Mimulus g u f f a f u s plant at the time of measuring. Its 1 1 th flowering node is in bloom. The six internodes which are measured are shown between arrows pointing to the first and seventh flowering nodes.
VARIANCE COMPONENTS 771 length was extended to 16 hours with incandescent lamps. The temperature was maintained above a minimum of 16"C, and high temperatures in the neighbor- hood of 40°C were commonly observed on warm, clear days. The plants were grown singly in four-inch pots, and were assigned positions at random within the space allotted a given experiment in the greenhouse. The soil used in each experiment all came from the same source, and the filled pots were assigned posi- tions at random so that local soil gradients were not associated with local green- house environmental conditions. In order to approximate the natural condition of M . guttatus soils, the pots were placed in shallow trays of water so that the soil in each pot remained saturated throughout each experiment. Differences in growth of the six measured internodes were negligible between trays, and if present, random for any given clone or sibship due to the method of assigning plant position.
During the experiments, damage from slugs and small caterpillars was oc- casionally sustained by young plants, and the older plants were occasionally attacked by mealy bugs and frequently by leaf miners and fungus of the genus Botrytis. High smog levels were typically associated with the appearance of necrotic areas on the leaves of both young and older plants. These various causes did not result in severe damage or death of any plant in the experiments used to estimate variance components, although Botrytis killed several plants in the later selection experiments.
The nodes used for cloning the plants were from basal runners which were, with few exceptions, of comparable development and physiological condition. The dry seeds from crosses were heated at 62°C for 48 hours prior to germination, a practice which reduced the germination time variance by about 50 percent. The seeds generally began germinating about three days after they were placed on moist filter paper, with the majority of the seeds germinating within a three- day period. The seedlings were planted in preassigned pots about one day after germination. Axillary branches were clipped off at weekly intervals. reducing interplant competition for light compared to the situation observed in nature. Equalizing axillary branch growth also probably removed a genetic component of internode elongation variance not associated with competition. Branch removal was necessary: (1 ) As a convenience in handling the plants; (2) To prevent loss of smaller plants due to excessive competition; and (3) As a sanitation measure due to Botrytis infection. After a plant had flowered at ten nodes, it was measured and removed, giving the slowly-developing plants light conditions more com- parable to those of the faster-developing plants.
required about ten weeks, during September through early November, to com- plete development of ten flowering nodes, while the ramets required about 13 weeks, during late September through December, to reach a comparable stage of development. The average growth of the 105 clones was greater in the earlier, or ortet, block (297, log scale) than it was in the later, or ramet, blocks (167 and 196). The data were analyzed as a two-way, mixed model, analysis of variance
(SCHEFFE 1959, Chap. 8) (Table 1 ) .
F values for both clones and blocks are highly significant. The clonal com- ponent, represents the amount of variation due to differences between the clones, which may be due to genetic differences, or differences due to a common physiological or morphological condition of the ortet and two ramets at the time of cloning with respect to each clone ("c" effects)
,
or both. Since the 105 clones are a random sample of a population, u f l is an estimate of this population parameter.The blocks component, + h l , represents the amount of variation due to replica-
tions of each clone being grown in different areas of the greenhouse (all three blocks), and at different times of the year (ortet block us. the two ramet blocks). These three blocks cannot be considered as random samples of a realistic popula- tion of environments. Therefore, estimates only an effect of the three environments.
The error component, U:, represents the residual variation not assignable
directly to either the three blocks or 105 clones. Local environmental fluctuations or differences in such things as temperature, humidity, light, pathogens and soil, coupled with differences in the time of reaching a given stage of development, would contribute to this variation. These would be a random sample of a realistic population of environmental causes of variation in the greenhouse, and therefore
U: estimates a population parameter. However, there may be an interaction be-
tween the gross environmental differences of the three blocks and the clones. This would not constitute a sample of a realistic population of such interactions. Since the clones are not replicated within blocks, this effect is not directly estimable, and would be included in the component U,". In this case, however, an analysis of
supplementary data did not detect such interaction, indicating the :U primarily
estimates the variation due to local fluctuations and differences in environment. Assuming that uZ2 represents only genetic variation, broad-sense heritability may be estimated from the data in Table 1 by the relationship h', = U ; J ( U ~ ~
4-
U:) = 0.43 with 95 percent confidence limits of 0.31
<
hi<
0.55 (SCHEFFE 1959,pp. 229-31). A more restricted estimate based on the relationship h', = U",,/
(U:,
+
U:+
+*[) may also be obtained (= 0.32), but has meaning only withrespect to the blocks involved.
Two selection lines were established by all possible crosses (including re- ciprocals) within the seven longest and within the seven shortest clones, forming
A
A
A
A
A A A
A
A A
VARIANCE COMPONENTS
TABLE 1
Log of &-internode length of 105 random clones
773
Mean Expected Estimated
mean square component
Source df square F F.95
A
Blocks 2 486,736 64.52 3.04 a:
+
105Gbl 9,, = 4564.Clones 104 24,637 3.27 1.32 a:+ 30:~ a:L =5697
Error 208 7,545 (12 = 7545
A
A
95 percent confidence intervals
(SCHEFFE 1959, p. 229) 6288
<
ai<
92243788
< <
8480 (BULMER 1957; SCHEFFE 1959, pp. 2 3 - 3 5 )21 long-internode and 21 short-internode families. Such a system of assortative mating substantially increases the accuracy of the parent-off spring regression coefficient (REEVE 1953,1955,1961 ; BREESE 1956). Assuming a linear regression, the regression coefficient ( b ) of the average value of the offspring of one mating on the average value of the parents provides an estimate of additive genetic variance, which is biased if maternal effects or (additive) 11 epistasis are present
(GRIFFING 1960; FALCONER 1960, Chap. 10). In the clonal experiment, b = 0.49, with 95 percent confidence limits 0.40
<
b<
0.59. Since parental values are the means of three members of a clone, this is not a direct estimate of narrow-sense heritability. The additive genetic component (U:) is estimated by b( u,"~+
u,2/3) -buzclU= 4049, where the variance of the clonal averages of the 105-clone sample.Three more point estimates may be obtained using the clonal data. The first is narrow-sense heritability: h: = u:/(u:.~
+
U,') = 0.31. The second is the amount ofnonadditive variance present: u : ~ - U," = 1648. This would include most of the
epistatic variance (except the fractional (additive) components included in the estimate of U,"), the dominance variance, interactions of common genetic systems
with the parental and progeny environments, and "c" effects of cloning. The third is the proportion of u : ~ which is additive: u ~ / u ~ ~ = 0.71.
Estimates bas,cd o n uncloned plants: An assortative mating scheme similar to that described above was carried out with the 319-plant population sample. A midparcnt-off sprinz b of 0.55 was obtained, with 95 percent confidence limits 0 48
<
b<
0.62. Assuming negligible maternal effects and (additive)" epistasis, this is a direct estimate of narrow-sense heritability. The additive genetic com- ponent is estimated by bui = 6177, where U; is the phenotypic variance of the319-plant sample. The 95 percent confidence limits for the two population sample variances are 6316
<
u:~,'<
10,993 and 9617<
U:<
13,222. Multiplying theselimits by their respective b point estimates gives the overlapping ranges 3114
<
AA A A
A A
-
A A A A
A A
A
A A
U:
<
5419 and 5280<
U;<
7259 for the clonal and uncloned regression estimatesof U,:, with an unspecified probability less than 95 percent that such a range
includes the parametsr U:.
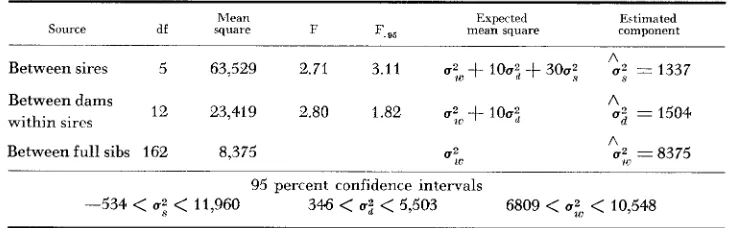
An independent sib analysis employing ten offspring per dam, three dams per sire, and six sires was used (FALCONER 1960, p. 173 and 183). The 24 parents were randomly collected from the Sierra population in June, 1960, and randomly assigned male or female status and mating groups. Two of the plants were nearly male-sterile, and therefore nonrandomly designated females. The analysis is presented in Table 2.
The additive genetic component is estimated by 4 2 = 5348 with little bias and poor accuracy, or by 4 ~ : = 6016 with an improvement in accuracy, but also an increase in bias dependent on the magnitude of nonadditive genetic components and maternal effects included in U:. The nonadditive components in U:, except
the fractional (additive)" components also contained in U', are estimated by U: - U: = 167. a value of low precision, but indicating minor nonadditive and
maternal effects variance on the basis of this one point estimate. Assuming only additive genetic variance, narrow sense heritability is estimated by the ratio 2 ( ~ :
+
u ~ ) / u : , ~ ~ = 0.51 with 95 percent confidence limits 0.21<
h:,<
1.06 (GRAY-BILL, MARTIN and GODFREY 1956), and environmental variance is estimated by The various estimates. with confidence limits where appropriate. are compiled for comparison in Table 3. These estimates are presented as applying only to the principal component in the estimate, assuming for the present that the various possible biases discussed above are nonoperative. While several of the point estimates do not closely agree, or are irreconcilable (such as h:
>
h i ) , when confidence limits are considered all estimates can be worked into a single system. Three generations of selection for long and short internodes, based on indi- vidual plant performance, developed a cumulative selection differential of 1254, and a cumulative response of 438. Using the ratio of response to selection differ- entia1 as an estimate of narrow-sense heritability, hi = 0.35, a value agreeingTABLE 2
Log of six-internode length of seedling progeny A
A A
A A A
A A A A
U?" - U ; - U ; = U ; = 5534.
A A
A
Mean Expected Ectimated
Source df square B F. 95 mean square component
A
Between sires 5 63,529 2.71 3.11 u : ~ + 100;
-t
300; U: == 1337Between dams
within sircs I 2 23,419 2.80 1.82 u : ~ ~ + 1002,
A
02
= 1504 ABetween full sibs 162 8,375
e,.
ut, = 837595 percent confidence intervals
VARIANCE COMPONENTS 775
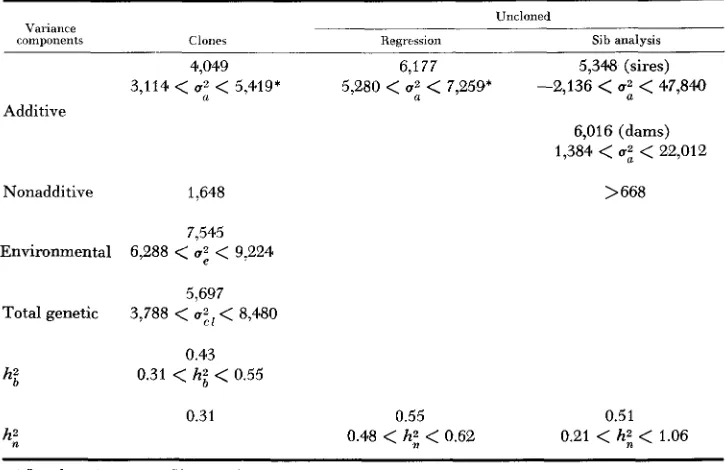
TABLE 3
Point estimates and 95 percent confidence limits of variance components and heritabilities
Variance
components Claries
4,049 3,114
<
U:<
5,419*Additive
Nonadditive 1.648
7,545 Environmental 6,288
<
17;<
9,2245,697 Total genetic 3,788
<
<
8,4800.43 0.31
<
hi<
0.550.31 hi
h i
Uncloned
Regressrun Sib analysis
6,177 5,348 (sires)
5,280
<
17:<
7,259' -2,136<
17:<
47,8406,016 (dams) 1,384
<
U:<
22,012>
6680.55 0.51
0.48
<
hi<
0.62 0.21<
hi<
1.06* Less than 95 percent confidence lerel
more closely with the clonal estimate than with that of the uncloned designs. DLSCUSSION A N D CONCLUSIONS
With respect to the relative usefulness of these analyses, the clonal analysis provides good estimates of the environmental component of variance, the total genetic component of variance, and broad-sense heritability, with confidence limits available for all three. By combining the clonal analysis with a parent- off spring regression, estimates of the additive genetic and nonadditive genetic components can be obtained. Narrow-sense heritability may be estimated directly, with confidence limits available, using a regression of off spring on single members of random clones. This procedure is not valid for selected parent clones, however, and narrow-sense heritability must be estimated by the ratio of the estimated additive and total variances.
The parent-offspring regression using uncloned plants gives a good estimate of narrow-sense heritability, with confidence limits available, and an estimate of the additive genetic component with about the same accuracy and bias as the clonal estimate. No statements can be made about the relative values of environ- mental, total genetic and nonadditive components, or broad-sense heritability from this analysis alone.
the nonadditive component and an approximation of the environmental com- ponent can be estimated with much less confidence. No estimate of broad-sense heritability is possible from this analysis alone. The relatively poor accuracy of estimates from the sib analysis in Table 3 is due to the smaller number of plants used in this experiment, and the lower efficiency of sib analyses in this range of heritabilities (ROBERTSON 1959).
Two objections may be raised with respect to a clonal scheme on biologica1 grounds. First, a plant originating as a vegetative propagule may not be com- parable to a plant of seedling origin. This objection may be avoided, in part, by cloning the seedlings, so that a parent-offspring regression is based only on vege- tatively-propagated plants. However, the applicability of estimates from such a cloned experiment to seedling populations would be uncertain. The second objec- tion concerns the probability that ramets of the same ortet will be similar not only due to a common genotype, but also due to environmental modifications of the physiology and morphology of the ortet which are more or less shared by the ramets. This is similar to maternal effects, and would appear as nonadditive genetic variance in the above scheme of clonal-regression analysis. It is possible to circumvent this problem by combining a random population of seedlings and clones, or by a two-stage cloning scheme (LIBBY and JUND 1962). Fortunately, these disturbances are small, if present at all. with respect to the six internodes measured in these experiments.
SUMMARY
Environmental, total genetic, additive genetic, and nonadditive genetic com- ponents of variance, and broad-sense and narrow-sense heritability, are estimated for internode length by analyzing a cloned population of Mimulus guttatus and progeny from crosses within this population. Independent estimates of some of these parameters, in general compatible with the clonal estimates, are obtained using uncloned parent-off spring regression and an uncloned sib analysis. The ratio of response to selection differential from a selection experiment conforms better to the clonal estimate of heritability than to the estimates from the more conventional designs. The relative usefulness of these different schemes is dis- cussed.
ACKNOWLEDGMENTS
VARIANCE C O M P O N E N T S 777 L I T E R A T U R E C I T E D
BREESE, E. L., 1956 BULMER, M. G., 1957
44: 159-167.
BURTON, G . W., and E. H. DEVANE, 1953
COOPER, J. P., 1959
FALCONER, D. S., 1960
GRAYBILL, F. A., F. MARTIN, and G. GODFREY, 1956
GRIFFING, B., 1960
LIBBY, W. J., and E. JUND, 1962 REEVE, E. C. R., 1953
The genetical consequences of assortative mating. Heredity 10: 323-343. Approximate confidence limits for components of variance. Biometrika
Estimating heritability in tall fesque (Festucu
Selection and population structure in Lolium. 11. Genetic control of date arundinacea) from replicated clonal material. Agron. J. 45 : 478-481.
of ear emergence. Heredity 13: 445-459.
Zntroduction to Quantitative Genetics. Oliver and Boyd. London. Confidence intervals for variance ratios
Theoretical consequences of truncation selection based on the individual specifying genetic heritability. Biometrics 12 : 99-109.
phenotype. Australian J. Biol. Sci. 13: 307-343.
Variance associated with cloning. Heredity (In press.) Studies in quantitative inheritance. 111. Heritability and genetic corre- (Contribution to the correlations between relatives in random mating populations- A note on non-random mating in progeny tests. Genet. Res. 2: 195-203.
lation in progeny tests using different mating schemes. J. Genet. 51 : 520-542. discussion.) Cold Spring Harbor Symposia Quant. Biol. 20: 76-78.
1955 1961
ROBERTSON, A., 1959 15: 219-226. SCHEFFE, H., 1959
Experimental design in the evaluation of genetic parameters. Biometrics