Copyright 0 1990 by the Genetics Society of America
Interacting Genes That Affect Microtubule Function
in
Drosophila melanogaster: Two Classes of Mutation Revert the Failure to
Complement Between
haync2
and Mutations in Tubulin Genes
Cathy
L.Regan' and Margaret
T.Fuller2
Department of Molecular, Cellular and Developmental Biology, University of Colorado, Boulder, Colorado 80309 Manuscript received October 20, 1989
Accepted for publication January 12, 1990
ABSTRACT
T h e recessive male sterile mutation haync2 of Drosophila melanogaster fails to complement certain &- tubulin and a-tubulin mutations, suggesting that the haywire product plays a role in microtubule function, perhaps as a structural component of microtubules. T h e genetic interaction appears to require the presence of the aberrant product encoded by hayncz, which may act as a structural poison. Based on this observation, we have isolated ten new mutations that revert the failure to complement between hay"" and B2t". T h e revertants tested behaved as intragenic mutations of hay in recombination tests, and fell into two phenotypic classes, suggesting two functional domains of the hay gene product. Some revertants were hemizygous viable and less severe than haync2 in their recessive phenotype. These mutations might revert the poison by restoring the aberrant product encoded by the haync2 allele to more wild-type function. Most of the revertants were recessive lethal mutations, indicating that the hay gene product is essential for viability. These more extreme mutations could revert the poison by destroying the ability of the aberrant haywirenc2 product to interact structurally with microtubules. Flies heterozygous for the original hay""' allele and an extreme revertant show defects in both the structure and the function of the male meiotic spindle.
T
HE genetics of the haywire locus (hay) of Drosoph-ila melanogaster suggests that this gene might encode a structural component of microtubules. T h e locus was identified through failure to complement
(REGAN and FULLER 1988) between the hayncz muta-
tion (3-34.4 m.u.) and certain alleles of B2t (3-48.5
m.u.), a gene that encodes a testis-specific @-tubulin
isoform (KEMPHEUS et al. 1980). Male flies heterozy-
gous for the haync2 mutation and a null allele of B2t
(haync2
+/+
B2t") are sterile, even though a wild-type copy of each of the two genes is present. T h e geneticinteraction between haync2 and B2t is allele specific.
T h e hay"c2 mutation fails to complement the null allele
B2t" and amorphic B2t alleles that encode unstable
p2-
tubulin subunits, but complements some of the B2t
alleles that encode partially functional products (RE- G A N and FULLER 1988). T h e hayncz mutation fails to complement a-tubulin as well as @-tubulin mutations.
For some but not all alleles of tubA84B, the gene
encoding the most abundantly expressed a-tubulin of
Drosophila (KALFAYAN and WENSINK 1982; MAT-
THEWS, MILLER and KAUFMAN 1989), males hetero- zygous f o r hay"" and a tubA84B mutation are sterile
(REGAN 1988). haync2 also fails to complement
ton, Indiana 47405.
versity School of Medicine, Stanford, California 94305-5427. Genetics 125: 77-90 (May. 1990)
' Present address: Departnlent of Biology. Indiana University, Blooming- ' Present address: Department of Developmental Biology, Stanford Uni-
whirligignc4, another mutation identified due to failure
to complement tubulin mutations (L. GREEN, N. WOLF
and M. T. FULLER manuscript submitted for publica-
tion). Thus hayncz is a member of a network of inter-
acting mutations. Although hayncz acts as a dominant
enhancer o f tubulin mutations, the hayncz allele is
recessive when in an otherwise wild-type genetic
background ( i e . , hayncz
+/++
males are fertile).Failure to complement between mutations in sepa- rate genes could indicate that the products of the two genes are involved in the same function. In at least two related cases where noncomplementing mutations map to separate genes, the products of the two genes are proteins that interact with each other at the struc-
tural level. Mutations in a-tubulin that show allele-
specific failure to complement with @-tubulin muta-
tions have been identified in both
D.
melanogaster andSaccharomyces cerevisiae (MATTHEWS and KAUFMAN
1987; STEARNS and BOTSTEIN 1988; HAYS et al. 1989).
&-Tubulin and @-tubulin form a heterodimer that
functions as the basic structural subunit of microtu-
bules (LUDUENA, SHOOTER and WILSON 1977). If the
functional relationship implied by the failure t o com-
plement between hayncz and B2t alleles is due to a
78 C. L. Regan and M. T. Fuller
tubulin genes of Drosophila, the hay gene product is
a candidate for a microtubule associated protein
(MAP) or other structural component of microtubule
arrays.
T h e phenotype of males homozygous for hay""' but
wild type for tubulin supports the conclusion that
haywire is important for microtubule function. hay""z/ haync2 males have defects in three microtubule-based processes during spermatogenesis: meiosis, flagellar elongation and shaping of the nucleus by cytoplasmic microtubules (REGAN and FULLER 1988). T h e same three processes (or a subset thereof) are defective in
males homozygous for B2t mutations (KEMPHUES et
al. 1982; FULLER et al. 1987). Male flies heterozygous for both hay"" and B2t mutations (hay""
+/+
B2t) also exhibit defects in microtubule-based processes during spermatogenesis (REGAN and FULLER 1988; RECAN1988).
An important characteristic of the genetic interac-
tion between haywire and the a-tubulin, P-tubulin and
whirligzg genes is that deletions of the hay locus com-
plement the mutations that hayncz fails to complement.
For example, males heterozygous for both a deficiency
of hay and a null allele of B2t
(Df(hay)
+/+
B 2 f ) arefertile (REGAN and FULLER 1988). This result indi-
cates that is not a null mutation, and that the
presence of the defective product encoded by the
hay""2 allele is required for the genetic interaction.
T h e aberrant product encoded by hayncz could act as
a structural poison of microtubule assembly or func- tion.
We have isolated new mutations that eliminate the
ability of the hayncz product to act as a poison by
selecting revertants of the dominant genetic interac- tion between hay""' and B2t". Most of the revertants
behave as intragenic mutations in haywire. The rever-
tants fall into two distinct phenotypic classes, indicat-
ing that reversion of the interaction between hayffcz
and B2t" can occur by two fundamentally different
mechanisms. Some revertants appear to partially re-
store haywire function. These mutants are viable when
heterozygous with a deficiency for hay, and flies het-
erozygous for the original hayffcz allele and a revertant
show less severe defects in spermatogenesis than
hay""/Df. In contrast, many of the revertants appear
to cause more extreme defects in haywire function
than the original allele. These revertants are lethal
when heterozygous with a deficiency of hay. Flies
heterozygous for these new mutations and the original haync2 allele survive to adulthood, but are male sterile with defects during meiosis that are more severe than those of huy""/Df. Defects in male meiosis were as- sayed in three ways. (1) T h e appearance and overall
organization of early postmeiotic cells in testis
squashes was examined by phase contrast microscopy
for abnormalities due to defects in the preceding
meiotic divisions. (2) Chromosome behavior was fol-
lowed by staining of condensed chromatin with aceto-
orcein. (3) Spindle structure was analyzed by immu-
nofluorescence microscopy of cells stained with anti-
tubulin antibodies. Flies heterozygous for haywire mu-
tants have defects in both the structure and the func- tion of the male meiotic spindle.
MATERIALS AND METHODS
Fly strains and culture: Flies were raised at 25" in humidified incubators on standard cornmeal agar media. Visible mutations and balancers are described in LINDSLEY and GRELL 1968; LINDSLEY and ZIMM (1985, 1987); and vi in ANXOLEBERE and PERIQUET 1973. Df(?L)E(r)lRP deletes 67E3,4-67F3,4 (R. JONES and W. GELBART, personal com- munication). B2tn, which is described in FULLER (1 986) and FULLER et al. (1988), was originally isolated as K M 2 4 by K. MATTHEWS.
T h e B2t" mutation is recessive at 25" but becomes a dominant male sterile mutation at 18 O . Based on this obser- vation, ri B2t" e/TM? flies were maintained in stock at 25", and new cultures were set up at 18" to serve as a source of virgin females. In crosses where these virgins were used, B2tn virgins represents a mixed population of ri B2tn e/ri
B2t" e and ri B2t" e/TM? females.
Selection of revertants: ru h hayncz vi/TM? males muta- genized with 25 mM ethyl methanesulfonate (EMS) follow- ing the protocol of LEWIS and BACHER (1968) were mated to B2t" virgin females, and male progeny of the genotype
haync2
+/+
B2t" were mated en masse (ca. 20/vial) to B2tn virgin females to test fertility (Figure 1). Progeny were produced only if an F I male carried a new mutation (*) that reverted the failure of haync2 to complement B2t". T h e new mutation was retested by crossing the resulting FZ haynr2*+/
+
B2t" males to B2t"/TM? virgins. Balanced third chromo- some stocks were made by mating the resulting Fs ru h hayncz* vi/TM? males with their virgin female siblings of the same genotype. In addition, the new mutations were kept under selection by crossing to B2t" females and selecting fertile haync2*+/+
B2t" male progeny every generation. By maintaining the mutation under selection, w e should have been able to recover any dominant extragenic revertants that mapped to chromosomes other than I I I . Except where stated otherwise, for all data reported in this paper the revertants were carried on the original ru h haync2 vi chro- mosome.Recombination mapping, complementation tests, and brooding tests: Localization of revertants was determined by recombination mapping between the revertant-bearing chromosome, ru h haynCzN vi, and a third chromosome car- rying the visible markers cur rs th st @ cu sr ca. Reciprocal recombinants between cur (3-30.0) and rs (3-35.0), or between h (3-26.0) and th (3-43.0) were picked. Segrega- tion of three phenotypes was scored: male sterility with haync2, lethality with Df(JL)E(z)lRZ, and reversion of the failure to complement the B2t"' allele. Fertility was scored by dissection of the seminal vesicles of five flies for each recombinant line.
Reverts of haywire"" 79
counting progeny through the 18th day after mating. At least 100 progeny flies were examined for each cross. T h e mutant combination was scored as lethal if no flies of the test class eclosed. Brooding tests were performed as de- scribed in RECAN and FULLER (1 988).
Determination of effective lethal phase: Crosses were set up for one night in bottles containing standard cornmeal- agar media, then transferred to bottles with no food, but fitted with a IO-mm Petri dish containing molasses-agar medium and a drop of yeast-water paste. T h e lids were removed and replaced periodically for egg collections. T h e eggs were counted and transferred to another cap contain- ing molasses-agar medium. After 36-48 hr, the number of eggs remaining unhatched was counted. Fertilized but un- hatched us. unfertilized eggs were distinguished as described in WIESCHAUS and NUSSLEIN-VOLHARD (1 986). After hatch- ing, additional yeast paste was added to provide food for the larvae, and the caps put into empty bottles to develop. T h e number of larvae that pupated was scored 7 days later.
Cytology and immunofluorescence microscopy:
Squashes of unfixed testes from young adult males were prepared for phase contrast microscopy as described in KEMPHUES et al. (1980), except that testes were dissected and squashed i n T B l buffer (7 mM K2HPO4, 7 mM KHnH04, 80 mM KCI, 16 mM NaCI, 5 mM MgCIz, 1% PEG 6000). Preparation of orcein-stained squashes of fixed testes for examination of nuclei containing condensed chromatin followed the procedure of LIFSHYTZ and HARAVEN (1977).
Testes were prepared for immunofluorescence micros- copy by a modification of the protocol of J. CASAL, G. GONZALEZ and P. RIPOLL (manuscript submitted for publi- cation). Testes were removed from males in buffer (1 0 mM Tris-HCI (pH 6.8), 47 mM NaCI, 183 mM KCI), transferred to a drop of the same buffer containing 10 PM tax01 on a slide, opened, and gently squashed under a coverslip. Im- mediately after squashing, the slides were frozen by immer- sion in liquid N2 and the coverslip removed with a razor blade, following which the slides were incubated in 95% EtOH at room temperature for 10 min to 2 hr. T h e squashed testes were then fixed in 3.7% formaldehyde in PBS for 10 min, rinsed twice in PBS, preincubated 10-30 min in fetal calf serum, then incubated overnight at 4" in a 1:20 dilution of monoclonal anti-a-tubulin antibody 3/A5 (PIPERNO and FULLER 1985). After washing twice for 10 min at room temperature in PBS, the squashed preparations were incubated for 1 hr at room temperature in a 1:2000 dilution of rhodamine-conjugated goat anti-mouse IgG (Cappel) secondary antibody. T h e samples were then incu- bated for 5 min in PBS containing 0.5 Pg/ml DAPI, rinsed twice in PBS, and mounted in gelutol.
RESULTS
Isolation of mutations that revert the failure to complement between haync2 and B2t": Although hayncZ
fails to complement B2tn, a deficiency of hay comple-
ments B2t" (REGAN and FULLER 1988). This result
indicates that the dominant genetic interaction be-
tween hay"" and mutant alleles of the B2t locus re- quires the presence of the defective product encoded
by the hay"" mutation. Based on this observation, we
designed a scheme to select revertants of the failure to complement between hay"" and B2t" (Figure 1). Three general types of revertants might be expected from this selection protocol: (1) intragenic revertants
EMS
P
F1
F2
\
r u h n c 2 v i TM3
d
Inc2vi
d
x
n B2t"emass mate
/
n B 2 t " e
\Q
ri B2t
"
e$!
select
ru h nc2*vi
n B 2 t n e
cf
x
TM3 n B2t"eFIGURE 1 .--Selection for revertants of the Failure to complement between hay"" and B2t". (P) Mutagenized TU h hay"" ui/TMr3 males
were mass mated to ri B2t" e virgin fenlales. F,) ru h hay"" ui/ri B2t"
e progeny males were tested for fertility by mass mating to ri B2t" e
virgin females. Only those males that carry a new mutation (*) that reverts the Failure to complement w i l l be fertile. (F2) Revertants were retested by mating ru h hay""' ui/ri B2t" e males to ri B2t" e /
Til.13 females, and balanced stocks constructed from the resulting progeny.
that reduce expression or function of the mutant haync*
product and thus act more like a deficiency, (2) intra-
genic revertants that either fully or partially correct the original hay"" defect so that the hay gene product no longer acts as a poison but more like the wild-type
product, and (3) dominant extragenic suppressors of
the interaction that could identify other components
of the complex in which the hay gene product and ,&-
tubulin function.
To select revertants, male flies bearing hayncz were
mutagenized, crossed to B2t" females, and their haync2
+/+
B2t" sons mass mated to virgin females. Fl malescarrying new mutations that interfere with the ability
of the hayncz product to act as a poison were selected
as fertile males in a background of sterile hayncz
+/+
B2t" heterozygotes (Figure l ) , and the new mutations recovered through the resulting progeny. Ten inde-
pendent revertants were recovered from 10,800 F l
males tested. One of the revertants was lost. As shown
in Table 1, all revertants tested are male fertile when
heterozygous with B2t", but male sterile when heter-
ozygous with the original hay"" mutation, indicating
80 C. L. Regan and M. T. Fuller
TABLE 1
Complementation tests for male fertility between hayncz'" and mutations that affect microtubule function
Revertant hay"" 821" B2t'" t ~ b A 8 4 B " " ~ wrl"''
haynr2 S S S S S
haync2 S F F F F
haynr2"' S F F F F
haynczm3 S F F F F
haynrZm4 S F F F F
haynr2"' S F F F F
haynrzm7 S F F F F
haynrZNX S F F F F
haynrzmY S F
S F F F
For each mutation listed in the column headings, mutantlTM3 flies were crossed to hayflcZW/TM3 flies and 2 5 mutant/haynC2"' male progeny tested for fertility by mating as described in MATERIALS AND METHODS. As controls, haynC2"/TM3 and/or mutantlTM3 sib- lings were also tested for fertility. In all cases, control males were fully fertile. Revertants 2 and 8 were also tested with B2t7, and the heterozygous combination was male fertile (data not shown). F =
male fertile; S = male sterile. Genotypes left blank were not tested.
nine revertants further characterized were all linked
to the third chromosome. We have designated the
revertants hay"c2""-haync2""Y to indicate that they were
induced on a chromosome carrying the hay"" muta-
tion and are thus likely to be doubly mutant. Mutations selected as revertants .of the failure to
complement between huync2 and B2t" also revert other
genetic interactions in which haync2 participates. T h e haync2 mutation fails to complement the B2t" and B2t7
alleles (RECAN and FULLER 1988), which encode sta-
ble, partially functional &-tubulin subunits (FULLER et
al. 1988). haync2 also fails to complement the a-tubulin
missense mutation tubA84B""' and the second-site
noncomplementing mutation wrZnc4 (Table l), both of
which themselves fail to complement B2t alleles (HAYS
et al. 1989; L. GREEN, N. WOLF, K. MCDONALD and M. FULLER, submitted for publication). In all cases
tested, the revertants complement these mutations.
Males heterozygous for a revertant and either B2t1*,
tubA84Bnc3j, or wrlnC4 were fully fertile (Table 1).
Most of the revertants are recessive lethal muta- tions: T h e observation that haync2 homozygotes have reduced viability when grown at 28" (REGAN and
FULLER 1988) suggested that haywire gene function
might be essential for viability. Revertants that lower
the level or reduce the function of the haync2 gene
product should show a more extreme phenotype than
the original haync2 allele in a wild-type tubulin back-
ground, and might be lethal. T o examine this possi- bility, we tested the viability of flies heterozygous for the revertants and a deletion of the hay region (Table
2). Although flies heterozygous for the original haync2
mutation and a deficiency of hay are viable at 25", seven of the nine revertants were lethal in combina-
tion with the deficiency (Table 2, column 1). This
result suggests that some of the revertants are more
extreme alleles of haywire, and that function of haywire is required for viability.
For most of the revertants, haync2""/Dfanimals sur-
vived embryogenesis but died during the larval
period. Flies carrying the revertant mutations were
first outcrossed to Ore-R so that there would be no
contribution of a balancer/balancer class to embryonic
lethality, then hayncZ2"/0re-R females were mated to
Df(?L)E(z)IR2/0re-R males. T o test for embryonic
lethality, eggs layed over a 2-hr period were collected,
counted, allowed to develop, and the number of fer-
tilized eggs that had not hatched by 36-48 hr after
oviposition was determined. For five of the revertants,
greater than 97% of the fertilized eggs laid by rever-
tant/Ore-R females hatched (Table 3). Similar results
were obtained from a control where Ore-R females
were crossed to Df(3L)E(z)IR2/0re-R males. T h e per-
cent of fertilized eggs that developed to pupation was determined in the same experiment. Although 95% of the embryos from the control cross pupated, for
revertants I , 2 , ?, 4 and 7, only approximately 2/3 of
the embryos survived to pupate (Table 3). Periodic
examination of the developing larvae suggested that
death occurred during midlarval development. T o
confirm that h ~ y " ~ ~ ~ / D f ( ? L ) E ( z ) l R 2 animals die as larvae, we examined the progeny of hayncZ2"/TM6B,Tb
females crossed to Df(?L)E(z)IR2/TMGB,Tb males for
revertants 1, 2, 3, 4 and 7. T h e presence on the
balancer chromosome of the dominant mutation Tb
allowed us to differentiate second and third instar
haynCZ2"/Dflarvae from their siblings heterozygous for
the balancer. N o Tbf "wandering" third instar larvae
were found for any of the revertants, indicating that all
hayncZnr/Df
animals die prior to late third instar.However, some medium-sized Tbf animals were pres-
ent, indicating that the lethal phase for haynczm/Dfcan
be as late as second to early third larval instar. TbC
second instar larvae that had become quiescent were transferred to a fresh plate of medium and observed. By the next day, they had ceased movement, become flaccid, clear and amorphous, and degenerated.
Two of the revertants, rv5 and ru6, that were lethal
when heterozygous with a deletion of the haywire
region (Table 2) also appear to be associated with
dominant maternal effect embryonic semilethal mu- tations. When either rv5/0re-R or rv6/0re-R females
were crossed to either Df(3L)E(z)lRZ/Ore-R or
Ore-R males, more than half of the resulting fertil-
ized embryos failed to hatch (Table 4). In contrast,
good embryonic viability was observed in crosses
where the chromosome carrying the revertant was
contributed by the male. The dominant maternal ef-
fect lethality associated with rv5 and rv6 does not ap-
pear to be due to haploinsufficiency of haywire. When Df (?L)E(z)lR2/0re-R females were crossed to Ore-R
R e v e r t s of haywire""
TABLE 2
Most of the revertants are recessive lethal mutations that form a single complementation group
81
Df" rv2 rv3 rv4 rv5 rv6 rv7 rv8
nc2 1?1/4?3'
rv2 O/ I 9 6
rv3 0/286 0 / I YOd
rv4 0/205 0/221 0/605
ru 5 0/500 0/22? 0/?22 0/?47
rv6 0/454 0/116 0 / 4 I 2 0/62?
rv8 67/182 70/380 15/113 I3/365 28/39 7 58/?93 64/?02
W l ' 0/?09
ru7 0/326 0/785 0/269 0/151 0 / 5 4 3 0/1044
r u Y 16/128 ms' 12/114 5 / 9 2 78/753 52/797 22/145
" Df was Df(3L)B(z)lR2. rul and rv7 were also tested with an independently derived deletion, Df(3L)lxd6, and were lethal.
'
Data presented as # test class progeny/total # of progeny recovered. ru h hayncz" vi/TM? flies were crossed to Df(3L)E(z)lR2/TM6B,D,so 2.5% of the progeny were expected to be the hay"""/Df genotype.
haynr2"" was not tested inter se with the other revertants because it was induced on a ru h haync2 red e chromosome that is not viable when heteroyygous with ru h hay"" vi. All other mutations were induced on a ru h hay"" vi background.
All inter se crosses were hayn""/TM3 X haynC2"/TM3, so that 33% of the progeny should be the double heterozygote class. However, the background ru h hay"" vi chromosome on which most of the revertants were induced exhibited reduced viability not due to the hay"" nutation; i n control crosses only 10% of the progeny of ru h hay"" vi/TM? parents were homozygous for the ru h haync2 vi chromosome. Therefore, in the znter se crosses, 10% representation of the double heterozygous class indicates no reduction in viability over the original
C ~ ~ ~ I I I O S O I I I ~ .
flies were recovered, but progeny counts were not made. nzs = male sterile. Genotypes left blank were not tested. All I hay??Czn'z/hay"rz"%
crosses done at 2 5 " .
TABLE 3
Lethal phase of hay"''"lDf
Genotype o f
females" nh hatched Percent pupated' Percent
r v l / + 1665 99.7 71.3
m2/+ 745 97.1 59.5
rv?/+ 1143 98.3 68.1
ru4/+ 689 99.1 65.7
ru 7/+ 774 99.4 61.7
+/+ 925 97.7 94.9
a In all cases.
+
stands for the Ore-R wild-type strain. ru/+ or +/+
females were crossed to Df(?L)R(z)lR2/0re-R males.'
n = total number of fertilized eggs scored. T h e small number of eggs that remained white after 36 h r were not included in this number.Percent of fertiliyed eggs that developed to pupation.
(Table 4). Thus rv5 and 7-716 are not likely to be simple
loss of function mutations at haywire. Tests with re-
combinant chromosomes generated for rv5 indicated
that the mutation responsible for the dominant ma-
ternal effect lethality is located on the left arm of
chromosome
ZIZ
distal to th, consistent with the posi-tion of haywire. Although many of the progeny of ru5/
Ore-R mothers did not survive embryogenesis, mouth
hooks and denticle belts were visible in many of the
unhatched embryos, indicating that they developed
nearly to hatching.
The lethal revertants form a single complemen- tation group: T h e revertants that were lethal when
heterozygous with a deficiency of the hay region were
also lethal in heterozygous inter se combinations
(Table 2), indicating that they belong to a single
complementation group. Although this result is most consistent with the conclusion that the lethal rever-
TABLE 4
Two of the revertants are associated with dominant maternal effect embryonic lethal mutations
Females Males n hatched
Percent
r v Y / + 792 35.7
ru5/+ +/+ 923 43.3 Of /+ m5/+ 889 89.9
+/+ rv5/+ 786 97.3
rv6/+ Df /+ 1455 13.5
m6/+ +/+ 1371 34.1
Of/+ rv6/+ 1432 95.2
+/+ rv6/+ 989 97.5
Df /+ +I+ 769 96.6
+/+ Df /+ 876 99.8
Symbols as in Table 3.
* For all experiments in this table, the chromosome carrying rv5
was a recombinant marked with TU h cu sr ca, in which the right arm of the original mutagenized third chromosome was exchanged by recombination with a marked third chronlosome (Table 5). T h e dominant maternal effect lethal mutation was also carried on a recombinant 7715 chromosome marked with ru h st pp cu sr ca, but not on a recombinant from which the revertant, the hay"" mutation, and the region of chromosome 3L from ru to vi had been crossed off.
'
In all cases, Of was Df(3L)E(z)lR2.tants are all alleles of haywire, in our system failure to complement alone may be an insufficient criterion to place two mutations in the same gene. As described below, we have shown that several of the revertants
cosegregate with the haywire locus in recombination
tests.
Some revertants appear to partially restore hay- wire function toward wild type: T w o of the rever-
tants, haynCZNR and haynC2NY, were viable as hemizygotes
82 C . L. Regan and M. T. Fuller
ized. In addition to reverting the failure to comple-
ment B2t mutations (Table l), haynCzN8 also partially
reverts several aspects of the hay"" recessive pheno-
type. hay""/Df males produce no progeny, females
have greatly reduced fertility, and adults of both sexes have deranged scutellar bristle patterns (REGAN and
FULLER 1988). In contrast, haynCZN8/Df males were
weakly fertile. Of 50 huyncZru8/Df males tested for
fertility by individual mating tests, 30 produced no
progeny, and the remaining 20 were slightly fertile,
producing fewer than 20 progeny each. The pheno-
type of haync2ruR/haynr2 is more severe than the pheno-
type of haync2NR/Df; males heterozygous for haynczN8
and the original haync2 allele were completely sterile
(data not shown). Although huynrZm8/Df females were
not as fertile as wild type, they were more fertile than
hayn"/Df females. Most haynC2NX/Df females produced
low numbers of progeny in individual mating tests. In
addition, bristles were normal in haynCzNR/Dfflies. T h e
revertants that were viable when hemizygous were
also viable with the other revertants tested (Table 2),
but failed to complement them for male fertility,
possibly due to the presence of the original hay"" mutation on the revertant chromosomes.
At least seven revertants map to the region of
chromosome 3L that contains haywire: T o determine whether the revertants were likely to be new muta- tions at the hay locus, the mutations responsible for the following three phenotypes were tested for cose- gregation in recombination tests: male sterility when
heterozygous with haync', lethality when hemizygous
with a deletion of the haywire region, and reversion of
the failure to complement mutant B2t alleles. Females
heterozygous for TU h haynrZyuvi and a third chromo-
some marked with the visible mutations cur rs th s t
Pp
cu sr cu were crossed to males bearing appropriate
visible mutations. Based on inheritance of the visible
markers spaced along the length of the third chro-
mosome, progeny carrying recombinant chromo-
somes resulting from a single cross-over that occurred
between cur (3-30) and rs (3-35.0) or between h (3-
26.0) and th (3-43.2) were picked. Reciprocal recom-
binants that had either crossed off the left arm of the
chromosome distal to cur or h or the left arm proximal
to rs or th along with the entire right arm of the third chromosome were tested for each revertant. In each case, the revertant segregated as if in the h-th or cur- rs interval. T o test inheritance of the hay locus within the interval, recombinants were crossed to flies bear-
ing hay"" and the recombinant/haynC2 progeny assayed
for male fertility. T o test inheritance of the lethal
mutation, recombinants were crossed to flies bearing
Df(3L)E(z)lR2 and the progeny scored for viability
of the recombinant/Df class.
For r u l , m2, rv3, ru4, m5 and rv7, the male sterility
of ru/haynC2 and the lethality of rvlDfcosegregated in
TABLE 5
The seven revertants tested map to the hay interval and cosegregate with haynCZ in recombination tests
Flanking
ru/haynr2 sterile rulhay"" fertile No. of re- markers Revertant rulDf lethal rulllf viable combinants (m.u.)"
ru 1 5(f) 2 7 h-th (17)
ru2 7(f) 4 12 h-th ( 1 7)
rv4 4(f) 6 10 h-th ( 1 7)
ru5 5(f) 3 8 h-th (17)
rv 7 3(f) 4 7 h-th (17)
rulhay"" ru/hay"'*
sterile fertile
No. Flanking
rv3 3(f) 3 6 cur-rs (5)
markers ru8 4(f) 6 10 cur-th ( 1 3)
Data presented as number of recombinants showing the indicated phenotype. Reciprocal recombinants were tested for each interval.
T h e size of the interval to which the mutation mapped is indicated in parentheses. (f) All recombinants that were sterile when heterozygous with hay"" were fertile when heterozygous with B2tn,
indicating that the revertant cosegregated with the haywire locus.
all recombinants tested (Table 5 ) , indicating that the
lethal mutation is tightly linked to the hay locus. Those recombinants that were sterile with haynC', and so had
inherited a mutation at the hay locus, were tested for
inheritance of the mutation responsible for reversion
of the genetic interaction with B2t by crossing them to flies bearing the B2t" allele. In all cases, recombi- nantjB2t" males were fertile, indicating that the mu- tation responsible for reversion was also tightly linked to haywire. With the exception of the tests for lethality,
the same experiments were done with haync2N8. T h e
four recombinants that were male sterile when het-
erozygous with hay"" were male fertile when hetero-
zygous with B2t", indicating that the revertant seg- regated with the hay locus. Although the number of recombinants tested was small and therefore the res-
olution of the cosegregation analysis is low, taken
together with the observation that the lethal rever- tants fall into the same complementation group and fail to complement a small deletion of the haywire
region, the results indicate that the revertants are
likely to be intragenic mutations in hay.
It remains formally possible that the more severe revertants (see below) are mutations that alter the function of a neighboring gene in addition to haywire.
T h e closest known gene to haywire is E ( z ) (R. JONES
and W. GELBART, personal communication). T h e
haywire""' allele and revertants m2, rv3, ru4, ru5 and 7-717 are all viable and fertile when heterozygous with
recessive lethal alleles of E ( z ) . These results suggest
that haywire and E ( % ) are separate loci and that the
revertants tested are not lethal due to lesions in E ( z ) .
T h e phenotypes of the revertants suggest that they fall into two mechanistically different classes. Rever- tants I , 2, 3 , 4 , 5 and 7 could be intragenic mutations
that remove the poison effect of haync2 by making
Reverts of haywire"" 83
contrast, could eliminate the poison effect of
haync2 by introducing a compensatory mutation in cis
that renders the aberrant haynr2 product more wild
type in function.
The lethal revertants are more severe than a de- ficiency in their effects on spermatogenesis: T h e revertants that are hemizygous lethal could be null
alleles that eliminate the poison effect of haync2 by
eliminating the altered gene product. If so, the phe-
notype of h~y"'~'"/huy""~ flies should be indistinguish-
able from that of Df(huy)/huync2. Like
Df(huy)/huynC2
and h ~ y " ' ~ / h a y " ' ~ (REGAN and FULLER 1988), males heterozygous for huy""2"/huy""2 are sterile and have defects in meiosis, flagellar elongation and nuclear shaping. However, for the seven revertants character- ized, in no case were the phenotypic effects on sper-
matogenesis in huync2m/huynCz identical to those ob-
served in
Df(huy)huynr2
males. For the lethal revertantsrv2-rv7, the defects in meiosis and flagellar elongation in h~y"'~"'/huy"'~ males were more severe than in Of/ haync2, indicating that the revertant mutations are
probably still antimorphic (Table 6). For rv8, the
revertant that appears to partially correct the defects
associated with the original haynr2 allele, the defects in
spermatogenesis appeared less severe in huync2NX/huync2
than in
Df/huy""
males (Table 6).For the above phenotypic comparisons, the effects of the different genotypes on meiosis were the most readily quantifiable. T h e severity of the defects in meiosis was judged by examination of the appearance and overall organization of early postmeiotic sperma- tid cysts in testis squashes by phase contrast micros- copy (see Figure 3, Table 6). A brief review of male meiosis in wild type is provided below to aid interpre- tation of the effects on the appearance of early post- meiotic spermatid cysts in the mutant animals. During spermatogenesis a single spermatogonial cell under-
goes four mitotic divisions to produce a cyst of 16
primary spermatocytes which proceed through
meiosis in synchrony, giving rise to 64 postmeiotic
spermatids (LINDSLEY and TOKUYASU, 1980). Cysts of
cells at the different stages of meiosis can be readily
identified in squashed preparations of unfixed testes based on changes in arrangement of cytoplasmic com-
ponents before and during meiosis (Figure 2). Follow-
ing meiosis, the mitochondria in each cell aggregate to form a derivative consisting of wrapped layers of
membrane, resembling an onion in cross-section. In
onion-stage early postmeiotic spermatids, the mito- chondrial derivative appears as a dark circle adjacent to the white nucleus in each cell (Figure 2a). If the preceding meiotic divisions were normal, all nuclei (arrowhead) in an early spermatid cyst are the same
size, all mitochondrial derivatives (arrow) are the same
size, and each mitochondrial derivative is associated
with a single nucleus.
Flies heterozygous for haynC2 and the revertant mu-
tations exhibit defects in early spermatid cysts indica- tive of defects in meiosis. Representative fields of the
types of defects observed and scored are shown in
Figure 3, panels a-f. T h e relative frequencies of these
defects among the different genotypes are given in
Table 6. For comparison, Figure 3a shows normal
onion stage spermatids from wild
type.
In the Ore-Rstrain tested, 93% of the cysts had completely normal
early spermatids, and
7%
of the cysts contained oneor a few cells with minor defects such as an abnormally
large mitochondrial derivative.
In haync2 homozygotes or huy""/Df(huy) heterozy-
gotes the most common (80-85%) onion stage phe-
notype was cysts of spermatids in which the nuclei
(arrowhead) and mitochondrial derivatives (arrow)
were of different sizes and often not correctly paired
(Figure 3c). These defects indicate that meiosis and cytokinesis occurred in most cells, but that many cells
did not receive an equal amount of chromatin or
mitochondria. Often, onion stage nuclei were hard to see by phase contrast microscopy.
Revertant huync2""*, which is viable when heterozy- gous with the deficiency, appeared to show less severe
meiotic defects. In huync2m8/huync2 males, 33% of the
onion stage cysts were intermediate between the char-
acteristic
buy""
phenotype and wild type (Figure 3b),and showed only mild defects. Flagellar elongation
was also more like wild type in huync2"'8/huync2 males
than in haync2 homozygotes. However, spermatid nu-
clei still failed to elongate.
Revertants rv2, ru3, ru4, ru5, ru6 and rv7, all of
which are lethal when heterozygous with a deficiency,
appeared to be more severe than either huync2 or
Df(huy)
in their effects on meiosis. In males hetero-zygous for the original haynr2 allele and any of these
latter revertants we frequently observed early sper-
matid cysts with 16 cells, each with a large mitochon-
drial derivative (Figure 3d), a phenotype most likely to arise from failure of cytokinesis after both meiosis
I and 11. Although no nuclei were visible in such cysts
by phase contrast microscopy, staining with DAPI revealed the presence of micronuclei next to the mi- tochondrial derivatives (data not shown, but see also
Figure 4, c and d). We also often observed cysts of 16
cells with nuclei as large as those of the premeiotic primary spermatocytes, in which cytoplasmic compo- nents had aggregated to one side of the nucleus (Fig- ure 3e). However, the aggregates did not resemble the well-organized mitochondrial derivatives charac- teristic of postmeiotic spermatids. It is possible that
cysts like those shown in Figure 3e failed to go through
meiosis successfully, arrested at an early meiotic stage,
84 C . L. Regan and M. T. Fuller
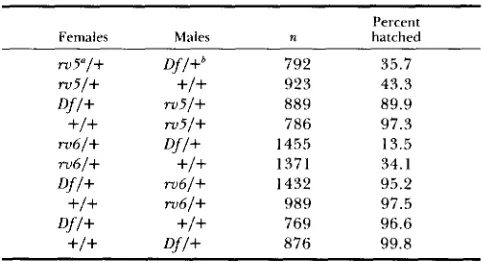
FIGURE 2.--Phase contrast light micrographs of testis squashes showing male meiosis in wild-type D . melanoguster. (a) Cyst of 16
spermatocytes i n pron~etaphase (1') to nletaphase (M) of meiosis I. The spindle and chromosomes are in the clear region in the center of each cell, and are outlined by phase dense layers of membrane and mitochondria (m) aligned along the periphery of the spindle. (b) Cyst of 16 cells i n metaphase (M). anaphase (A) and telophase (T) of meiosis I . (c) Cyst of 32 secondary spermatocytes in interphase between meiosis I
and 11. l h e nuclei (arrowhead) have reformed and cytoplasmic components (arrow) are clustered around them. (d) Cyst of 32 cells in meiosis
11. As i n meiosis I , membranes and mitochondria surround the the clear region that contains the spindle. The two meiotic divisions are distinguishable from one another by the size and number of cells in a cyst. (e) Cyst of early spermatids immediately after the completion of meiosis 11. Mitochondria (arrow) have begin to aggregate to one side o f each nucleus. (f) Cyst of onion stage early spermatids. The mitochondria in each cell have aggregated to form the mitochondrial derivative, a structure consisting of wrapped layers of membrane. The mitochondrial derivarives (arrow) appear a s dark circles adjacent to the white nuclei (arrowhead).
in both chromosome segregation and cytokinesis, re-
sulting in cysts of 16 cells with 4N nuclei, and in
addition had defects in formation of the mitochondrial
derivative. In males heterozygous for the original
haync2 allele and either 735 or ru6 we sometimes saw
aberrant cysts with no visible nuclei and "knotty"
mitochondrial derivatives that appeared to have large
holes (Figure 3f). ru5 and ru6 were the revertants
associated with dominant maternal effect embryonic lethality. I t is possible that both the embryonic phe- notype and the "knotty" spermatid cysts result from
unusual neomorphic or antimorphic properties of
these two alleles.
T h e percentage of aberrant cysts resembling panels
d, e, and f varied from one revertant to another. For
example, for ru3, the majority of cysts with a pheno-
type more extreme than haync2 homozygotes fell into
category e, while for ru4, the majority of the more
extreme cysts fell into category d. In general, for each
of the onion stage phenotypes described above, all
cells within a given cyst had the same phenotype. For example, we never saw a cell with a large nucleus and amorphous mitochondrial material (panel e) in a cyst
with cells that had bonufide mitochondrial derivatives.
For ru2, ru3, ru4, rv5, ru6 and
ru7,
flagellar elon-gation was also perturbed to a greater extent in
Reverts of haywire"" 85
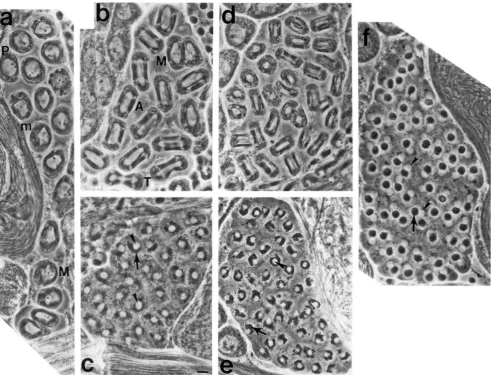
~ . ~ ; v R E :<."l'hcnotypes o f ' cysts o f cells that have attempted meiosis from ha~"""'/hn~"'' ;Ind from wild-type nl;lles. (a) M'ilcl-typc onion
stage early sperrn;ltids. ( I d ) Characteristic phenotypes of early post-meiotic cysts or cysts that appear to have attempted meiosis described in
text. The frequencies of these phenotypic classes in the different genotypes studied are shown in Table 6. (Arrowheads)-nuclei. (Small arrows
i n a ;Ind l>)-nornwl tnitocl1ondri;ll derivatives. (Large arrows in b-d and f)-abnormally large mitochondrial derivatives. (Large arrow in e)- cytoplasnlic ;~ggreg;~tes. (P)-primary spermatocyte. Uar, 10 p n .
. ..
TABLE 6
Frequency of meiotic and postmeiotic spermatid phenotypes in males heterozygous for different haywire mutations
G r l l o q " . ;I b c d e f Number
Ore-R 93 7 0 0 0 0 32
nr2/nr2 0 2 80 16 2 0 45 Qf(hayy/nr2 0 4 85 2 9 0 47 rv2/nc2 0 0 28 3 1 41 0 87 rv3/nr2 0 0 62 12 26 0 50
n 4 / n r 2 0 0 65 29 2 0 48
nv5lnr2 0 0 24 51 15 1 1 59 rv6/nr2 0 0 33 36 12 19 75 ~ 7 / n r 2 0 0 44 28 30 0 79 nIXlnc2 0 33 59 0 7 0 27
Columns a-f refer to the phenotypic classes shown in Figure 3. For e;tcll grnotvpc listed, both testes froln ten flies were examined for the presence ofcysts of cells that fell into one of the six categories shown i n Figure 3. Cysts of cells that were actually in meiosis could not be ;~ccur;~tely counted because the spindles were often so aberrant a s to be unrecogniz;lble i n squashed preparations viewed
by phase contr;tst microscopy. Data presented a s the percentage of cysts th;lt h;td a given phenotype out of the total number of cysts countetl for a 1 1 ten flies (n).
.,Df(hq) \vas D~(~L)E(Z)'"~.
fects in flagellar elongation could be a secondary effect
of the more extreme defects in meiosis in these males.
Spermatid nuclei failed to elongate in all h~y""~"/huy"'~ flies examined.
T h e last column in Table 6 shows the total number
of early spermatid cysts scored in ten flies of each
genotype. This number was higher than in wild-type
in the genotypes that had more severe meiotic defects.
In the severely defective males progress through the early postmeiotic stages could be blocked or proceed
more slowly. T h e less severe revertant, huynCZNX, had
a total number of postmeiotic cysts close to that of
wild type.
The revertants are recessive in their effects on spermatogenesis: For six of the revertants, defects in
TABLE 7
Brooding tests of hay"rzrv/+
Mur;lnt No. of progeny"
Nl 175 f 48.3
N 2 351.1 -t 38.9
N 3 418.3 f 82.1
N 4 392.1 2 68.7
rv5 306.4 f 54.8
~6 224 & 32.1 N 7 457.2 f 69.5
N R 405.8 2 59. I Ore-R 261.1 f 22.4
a Mean number of progeny produced per male. Ten rv/Ore-R
males were mated individually to three fresh virgin 821""" females every 3 days for 9 days. Progeny were counted through the 18th day after each mating.
spermatogenesis were more severe in huynCZN/huynC2
than in
Df/huyncZ
males. This observation indicatesthat these mutations are probably still antimorphic in nature. T o determine whether these mutations had dominant effects on spermatogenesis, males hetero- zygous for each revertant and wild type were tested for reduction in fertility as measured by the number of progeny produced by a single male in brooding tests. In all cases, the fertility of huynrZ"/0re-R was within the range of the fertility of the wild-type con-
trol males (Table
7),
indicating that the revertants arenot dominant in their effects on spermatogenesis, at least at the organismal level.
Both the structure and the function of the male meiotic spindle are defective in haywire mutants
Homologs fail to separate in meiosis I in
hayncZ males: Phenotypic analysis of early postmeiotic spermatid cysts by phase contrast microscopy indi- cated extreme defects in meiosis in h~y"'~"/huy"'~ ani- mals. T o examine the effects of this mutant combi-
86 C . L. Regan and M. T. Fuller
TABLE 8
Frequency of cysts in meiosis
I+sr d i v i ~ i w
1'-51 51-A Inc-I' 51 SI-A per nlalc
St*rond di-
visioll 1:requencv of
~ cyscs in meiosis
Orr-R 3 8 6 2 5 1 .R
nt2/nc2 1 X 1 0 0 0 0.95
7'rstrs f'ron~ 20 tnales of each genotype were squashed and stainrtl w i t h aceto-orcein, es;lnlined for cysts of cells i n meiosis, and t11r tlunlhcr of cysts at eacll stage i n meiosis determined. Only cysts cle;~rly in meiosis b;~secI on cellu1;tr nlorphologv were scored. Met- aphase ; I n d ;tn;lph;lse I were clistinguislled from metaphase and
;tn;tphasc I 1 hy the appear;lnce of the chromosomes ;IS bivalents at tnetq~llase I and uniwlents at met;~ph;tse I 1 (LIFSCHYTZ and HAR-
EVEN I 9 i i ) . I~cc;tusc meiosis occurs i n a wave across the cyst. a sitlglr cyst o f ~ c n rontained some cells i n metaphase and others in an;lph;lse. P-\I = cysts i n promet;lph;lse I and/or metaphase 1. M-A
= cysts with bo111 met;~ph;tse ant1 anaph;tse figures or cysts wholly i n ;In;lph;lsc. Int-P = it1terph;lsc and protnetaphase 11. M = meta- phase I l.
we prepared squashes of orcein stained testes from
huynr2rv2/huynr2 males. Testes from 20 wild-type flies and 20 huy"r2n'2/huynr2 flies were systematically scanned
and scored for cells in any stage of meiosis I, inter-
phase and meiosis I 1 (Table 8). In the wild-type males
scored, 50% of the meiotic cysts observed had a
meiosis I chromosome configuration, with the remain-
ing 50% either in interphase between the two divisions
o r in meiosis I1 (Table 8).
In contrast, homologous chromosomes apparently
b i l to separate in huynr2N2/huynr2 males. All of the cells
in meiosis observed in huynr2rv2/huynr2 were scored as
meiosis I based on the number and morphology of
the chromosomes (Table 8). In most meiotic cells in
bivalents, and sometimes the dotlike fourth chromo-
some bivalent. Most (74%) of the meiotic figures
resembled prometaphase, with the three bivalents
scattered (Figure 4a). A few cells had clustered biva-
lents resembling metaphase of meiosis I. Only once
did we observe chromosomes in apparent anaphase. T h e number of cysts scored as in meiosis based on cellular morphology was approximately the same in
spermatocytes are not arrested in meiosis in the mu- tant. Although aberrant, cytoplasmic rearrangements and other aspects of the meiotic cell cycle occur, many cells go on to the postmeiotic stages shown in Figure
3. However, the chromosomes appear to remain ar-
rested as unresolved bivalents even into the post-
meiotic stages in huynr2N2/huync2 males. In orcein
stained preparations where postmeiotic cells with mi- tochondrial derivatives were seen, we often observed
condensed chromatin adjacent to the mitochondrial
derivative (Figure 4d). In some cases, what appeared
to be the three large condensed bivalents occupied a
small nucleus adjacent to the mitochondrial derivative (Figure 4c). Chromosome arms were observed in some huync2n~2/huynr2 males we observed three large apparent
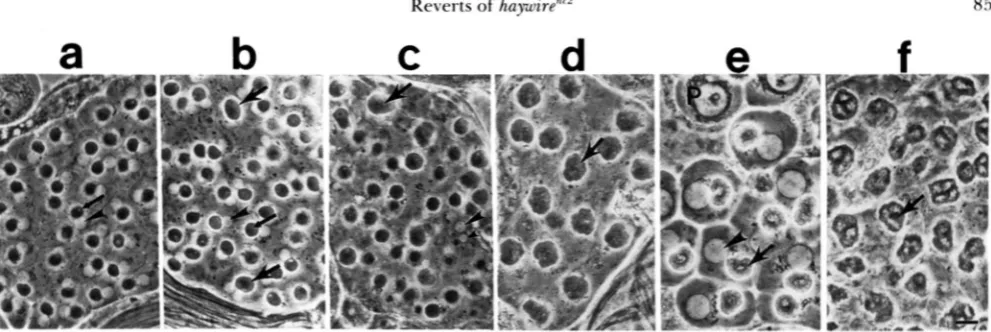
haynr2n12/huynr2 as in wild type (Table 8), indicating that
F l C U R E 4.-Bivalents form but homologs f i i l to separate in
hay"'2"'z/hay"r2 males. Light micrographs of testis squashes showing chromosomes stained w i t h orcein. (a) Meiotic cells in hay"'z"'z/hay"'z.
The majority of the chromosomes in meiosis in hay"'2"z/hay"'z males were in this prometaphase configuration (Table 7 ) . Note the three large condensed bivalents in each cell. (Arrow) bivalent with a hole in the center. (b) Meiosis I i n Ore-R wild-type male, showing cell in metaphase (arrowhead) and cells with chromosomes separating in anaphase I (short arrows). (c and d) Onion stage early spermatids from hay"'z"'2/hay"'z males. The nuclei are much smaller than the mitochondrial derivatives (arrows), and contain condensed chro- matin (arrowheads). Three clusters that may represent the three bivalents are visible in panel c. Note in panel d the apparent holes in the condensed chromatin, resembling the bivalents with holes in panel a. Bar, 10 pm.
spermatid nuclei, and in some cases the chromatin eventually appeared to decondense. T h e presence of highly condensed chromatin at the onion stage, re-
sulting in very small nuclei, may explain why nuclei
are often difficult to see in the type of cysts repre-
sented in Figure 3, c, d and f.
Although many mutations that affect meiosis also
affect mitosis (GATTI et ul. 1983), the mechanisms
involved in the two processes must be to some extent different, especially with respect to the pairing and
segregation of homologs during meiosis I. Even
though the meiotic chromosome cycle appears to ar-
rest in huync2"'2/huynr2, the premeiotic mitoses seem to
Reverts of haywirenr2 87
F I G U R E .5.-\lciotic spindles are ; ~ l m o r n l ; ~ l i n hay""""//~oy"'' males. Inllnullofluor~se~~ce microscopy of cells i n meiosis i n squashed preparations of testes stained with antibody to tubulin (a. c and e) and w i t h DAPl (b, d and
f).
(a and b) Wild type. Arrow point to spindleand corresponding nletaph;lse chron~oson~es. Note that the bivalents are clustered into a single bright spot at the metaphase plate i n each cell i n b. ( c and (I) hay"z""/hay"'2 cyst in prometaphase I. Note that the three bivalents in each cell are still separate in d. (e and f ) hay"'2""'z/
hay"" cvst a t :I later stage i n meiosis than i n c . Note that the chronloson~es i n each cell in f a r e clustered into a single bright spot, :IS i n b.
sence of gross defects in the preceding germ line
mitotic divisions (data not shown). We also found no evidence for gross mitotic defects in dividing cells in
larval brains from six haynr2n'2/haync2 animals. I t is
possible that the chromosome cycle defect observed
in h ~ y " ~ ~ ~ ' ~ / h a y " ~ ~ males is specific to meiosis.
Asters form but full metaphase meiotic spindles were not observed in haync2rv2/haync2 males: Muta-
tions in haywire affect the structure of the male meiotic
spindle as well as its function. Meiotic spindle mor- phology was examined by immunofluorescence stain- ing of testes from h ~ y " ~ ~ " " ~ / h a y " ~ ~ males with antibody against a-tubulin. Small microtubule asters were ob-
served at the positions expected for spindle poles
(Figure 5c) in cysts of spermatocytes that could be
judged as in prometaphase based on the appearance
of their DAPI-stained chromosomes (Figure 5d).
However, the fully organized metaphase spindles typ- ical of wild-type (Figure 5a) were never observed in the mutant, even in cysts where the chromosomes had
clustered together as if in metaphase. In some cysts
containing cells with clustered chromosomes typical
of metaphase I (Figure 5f), we observed cells that
appeared to have four asters, as if in meiosis I1 sub-
sequent to failure of both chromosome segregation
and cytokinesis (Figure 5e).
DISCUSSION
T h e model that the failure to complement between
haync2 and B2t mutations is based on a structural
interaction between the products of the two genes,
and that the aberrant gene product encoded by the
haynr2 allele acts as a poison when incorporated into
microtubule arrays (REGAN and FULLER 1988), pre-
dicts that the genetic interaction should be reverted by eliminating the ability of the aberrant hay"" prod- uct to act as a poison. We have identified ten new
mutations that revert the failure to complement be-
tween hay"" and tubulin mutations. A t least seven of
the revertants appear likely to be intragenic mutations
and will be considered as new alleles of hay for the
purposes of this discussion. These revertants localize
to the region of the chromosome that contains hay,
cosegregate with the hay"" mutation in recombination
tests, and in most cases are lethal in heterozygous
combination with a deletion of the haywire region.
In all cases tested, mutations isolated because they
reverted the failure to complement between hay""
and B2t" also reverted the failure to complement between hay"" and other B2t alleles, between hay"'2
and alleles of the tubA84B a-tubulin gene, and be-
tween haync2 and the wrZnr4 mutation. T h e observation
that each of these interactions is reverted by the same
mutation suggests that the genetic interactions be-
tween hay and the P-tubulin, a-tubulin, and whirligig
genes all share a common mechanism based on t h e
action of the poison product encoded by the hay""
allele.