The Log-Linear Relationship Between Sexual Isolation
and
Sequence Divergence
in
Bacillus Transformation is Robust
Piotr Zawadzki,' Michael
S.
Roberts2 and
Frederick
M. Cohan
Department of Biology, Wesleyan University, Middletown, Connecticut 06459-01 70 Manuscript received September 15, 1994
Accepted for publication April 8, 1995
ABSTRACT
The relationship between sexual isolation and sequence divergence in Bacillus transformation was
previously shown to be log linear. In the present study, we have shown that this relationship is robust with respect to naturally occurring genetic variation among recipient strains of Bacillus subtilis and B. mojuvensis. Naturally occurring restriction endonuclease activity was shown not to affect this relationship. Also, seven out of eight recombination mutants tested for their sensitivity to sequence divergence have shown the same relationship between sequence divergence and sexual isolation; a mutant for recH was more sensitive to sequence divergence, suggesting that the product of this gene may be involved in resolution of mismatches in heterogamic transformation. We have also shown that the relationship between sexual isolation and sequence divergence is robust with respect to variation in the conditions
of transformation, including variation in the length of donor DNA, the concentration of donor DNA, and intracellular competition between donorderived and recipient-derived DNA. The robustness of the relationship between sexual isolation and sequence divergence among naturally occurring strains and across transformation conditions allows us to predict the eventual outcome of sequence divergence among B. subtilis and its closest relatives.
G
ENETIC exchange in bacteria is much more pro- miscuous than that in the eukaryotes. In the sex- ual eukaryotes, populations that are separated by only2%
sequence divergence are frequently unable to ex- change genes, while bacteria may exchange genes with organisms that are much more divergent in DNA se- quence (YOUNG and LEVIN 1992; COHAN 1994a). For example, transformation between Bacillus species that are 5% divergent is reduced only threefold compared with transformation within species (ROBERTS and CO-HAN 1993).
The promiscuous nature of genetic exchange in bac- teria leads to two possible outcomes of neutral sequence divergence among bacterial species (COW 1995). First, interspecies recombination may be so rare that the species are free to diverge without bound in neutral characters. In this case, the species will diverge eventu- ally at every nucleotide site where a neutral substitution is possible. Alternatively, interspecies recombination may be frequent enough so that neutral sequence diver- gence between species is held to an equilibrium level.
COW (1995) has presented a coalescence model to predict the eventual outcome of neutral sequence
CorreSpondzng authur Frederick M. Cohan, Department of Biology, Wesleyan University, Middletown, CT 06459-0170.
E-mail: [email protected]
'Present address: School of Biological Sciences, University of Sussex, Falmer, Brighton BNl 9QG, United Kingdom.
'Present address: JRDC/MSU Joint Microbial Evolution Project, Mi- crobiology Laboratoly, RIKEN Institute, 2-1 Hirosawa, Wakoshi, Sai- tama 351-01, Japan.
Genetics 140 917-932 (July, 1995)
divergence among species from several estimable popu- lation genetic parameters. The model takes into ac- count a positive feedback loop between sexual isolation and sequence divergence. In this loop, increased sexual isolation leads to greater sequence divergence, because sexual isolation opposes the homogenizing effect of gene flow (COHAN 1994b). Reciprocally, increased se- quence divergence among species causes greater sexual isolation (ROBERTS and COHAN 1993). This is because mismatch in donor and recipient sequences interferes with recombination (TE RIELE and VENEMA 1982a,b, 1984). The degree to which sexual isolation is affected by sequence divergence can determine the outcome of the positive feedback loop: if sexual isolation is highly sensitive to sequence divergence, the feedback loop runs away to unbounded sequence divergence; with lower levels of sensitivity, the feedback loop can lead to an equilibrium level of divergence ( COHAN 1995). The present study addresses the sensitivity of sexual isolation to sequence divergence in Bacillus transformation in order to predict the eventual outcome of neutral se- quence divergence among Bacillus species.
Bacillus subtilis laboratory strain 1A2 and test donors is closely predicted as a log-linear function of the DNA sequence divergence between donor and recipient (ROBERTS and COHAN 1993). Over three orders of mag- nitude of sexual isolation, the following formula pre- dicted sexual isolation from sequence divergence with a coefficient of determination of 96%:
l o g d p ) = #JT
+
a (1)where p = sexual isolation, T = sequence divergence,
4
= sensitivity of sexual isolation to sequence diver- gence, and a = the intercept. The sensitivity parameter4
was estimated at 18.87. Assuming a log-linear relation- ship between sexual isolation and sequence divergence, with a slope4
of 18.87 ? 1.72 (SE), COHAN (1995) predicted that B. subtilis and its closest relatives [ B. moja- vensis (recently described by ROBERTS et al. 1994), B. a t r o p h m , B. amyloliquefaciens, and B. lichenijimis] are destined to diverge without bound in neutral charac- ters.In this study, we investigate whether the log-linear form of the relationship between sequence divergence and sexual isolation, as well as the estimated value of
4,
are sufficiently robust to allow predictions of evolu- tionary divergence. We investigate first whether all re- cipient genotypes yield the same relationship between sexual isolation ( p ) and sequence divergence ( T ) , andsecond, whether the relationship between p and T is
robust with respect to the conditions of transformation. We assayed the relationship between sexual isolation and sequence divergence for a series of recipient geno- types by following the method of ROBERTS and COHAN
(1993). Our approach was to transform a given recipi- ent toward rifampicin resistance, a trait coded by rifR alleles of ?OB, using donor DNA extracted from rifR mutants of related strains. Sexual isolation between the recipient and a test donor was quantified, as in the earlier study (ROBERTS and COHAN 1993), as the ratio of the homogamic to heterogamic transformation fre- quencies. For each recipient, we estimated the sensitiv- ity
( 4 )
of sexual isolation to sequence divergence asthe slope of the log-linear function in Equation 1 (using sexual isolation and sequence divergence data for a series of test donors), and we assayed the coefficient of determination of the log-linear function.
We first investigated how the sensitivity of sexual isola- tion to sequence divergence may be affected by muta- tions in the four recombination pathways used in B.
subtilis transformation (pathways characterized by
ALONSO et al. 1991, 1992). ALONSO and coworkers
(1992) have constructed a series of isogenic mutant strains that are deficient in each of the recombination pathways. We have compared the sensitivity parameter of each mutant with that of the isogenic wild-type strain. We tested two mutants of each of the four recombina- tion pathways: mutants for the recR and recL genes of the a pathway [corresponding, respectively, to recR (ALONSO et al. 1990) and rec0 (J. C. ALoNSO, personal
communication] of Escherichia coli), mutants for the addA and addB genes of the
P
pathway [corresponding to recBCD of E. coli (KOOISTRA and VENEMA 1991)1,
mu- tants for the recH and recP genes of the y pathway, and mutants for the recB and recG genes of the E pathway (whose E. coli counterparts are unknown) (J. C. ALONSO, personal communication).We next investigated the effect of restriction endonu- clease activity on the relationship between sexual isola- tion and sequence divergence. This work was motivated by our earlier result that some Bacillus strains isolated from nature have restriction endonuclease activity and that restriction activity in naturally occurring strains re- duces the rate of transformation by unmodified donors (COHAN et al. 1991; but see TRAUTNER et al. 1974 and IKAWA et al. 1980).
We have also assayed for variation in sensitivity to sequence divergence among naturally occurring, non- restricting strains of B. subtilis and B. mojavensis. This work was motivated by several considerations. First, this approach allowed us to address whether genetically based factors other than restriction activity and recom- bination enzymes may affect the sensitivity of recombi- nation to sequence divergence. Second, the variation in the sensitivity parameter #J that is most relevant to prediction of future sequence divergence is the varia- tion occurring in nature. Finally, the model for pre- dicting future sequence divergence among species re- quires that recombination in each species is equally sensitive to sequence divergence (COHAN 1995). There- fore, to predict divergence among B. subtilis and closely related species, we had to test that wild-type strains of these species follow the same relationship between sex- ual isolation and sequence divergence.
We next addressed the effect of the conditions of transformation on the relationship between sexual iso- lation and sequence divergence. We first investigated whether the relationship may be affected by the length of donor DNA fragments presented to the recipient cell, as suggested by ROBERTS and COHAN (1993). A
duplexes in favor of the recipient strain. If mismatch repair plays a role in limiting heterogamic transforma- tion in B. subtilis, longer donor fragments may cause sexual isolation to be less sensitive to sequence diver- gence.
We investigated the effect of donor fragment size on the sensitivity to sequence divergence by using, as do- nor, PCR-amplified DNA of different lengths as well as much longer segments of genomic DNA. The present approach departs from the work of ROBERTS and CO-
HAN (1993) in that the earlier work contrasted the effect of only one length of PCR-amplified DNA with that of genomic DNA.
We have also investigated the effect of donor DNA concentration on the relationship between sexual isola- tion and sequence divergence. Just as longer donor fragments may potentially saturate the mismatch repair system, very high concentrations of donor DNA may also potentially saturate mismatch repair. This is be- cause multiple donor molecules may independently en- ter the recipient cell under high donor DNA concentra- tions (the process of “congression”) (DUBNAU 1982); this would be expected to increase the challenge im- posed upon mismatch repair enzymes during hetero- gamic transformation.
Finally, we have investigated how the relationship be- tween sexual isolation and sequence divergence may be affected by intracellular competition between exoge- nous recipientderived and donorderived DNA. B. subti- lis appears to grow in nature as colonies on soil particles (LORENZ et al. 1988). Therefore, when a competent cell in nature takes up exogenous DNA from a potential donor strain (other than its clonemates in the colony), the cell may simultaneously take up exogenous DNA from its clonemates. The simultaneous uptake of DNA from recipient and donor strains may lead to competi- tion between exogenous, homologous DNA segments (derived from different strains) for the one homolo- gous site on the chromosome. We have tested whether this intracellular competition between exogenous DNA from donor and recipient strains might exaggerate the effect of sequence divergence on sexual isolation.
MATERIALS AND METHODS
Strains B. subtilis strains 1A2 (a derivative of strain 168) and 2A2 (a derivative of strain W23) were obtained from the Bacillus Genetic Stock Center (BGSC). The type strains of B. amylolaquefaciens (ATCC 23350) and B. lichenifonis (ATCC 14580) were obtained from American Type Culture Collec- tion (ATCC). The type strain of B. atrophaeus (NRRL NRS 213) was obtained from the Agricultural Research Service Culture Collection at the National Center for Agricultural Utilization Research. All B. subtilis recombination mutants and the isogenic wild-type strain YB886 were kindly provided by JUAN C. ALONSO. All naturally occumng strains of B. subtilis and B. mojavmis were isolated from soil as described by Co- rn and coworkers (1991). All strains of B. subtilis used in this study belong to one of two sequence-similarity groups: the “168 group” and the “W23 group” (ROBERTS and COHAN
1995), as listed in Table 1. All strains used in this study and their genotypes are listed in Table 1.
Screening for endonuclease restriction activity in naturally occurring strains of B. subtiilk: Ten randomly chosen strains of B. subtilis isolated from Death Valley, CA (DV3-G2, DV3-
D-1, DV2-F-1, and DV2-&2) were screened for endonuclease restriction activity following the method of COW et al.
Isolation of rifampicin-resistant mutants: For each strain to be used as a donor in transformation, rifampicin-resistant mutantS were isolated as described by ROBERTS and COW
(1993).
Purification of genomic DNA Genomic DNA was ex- tracted and purified according to COW et al. (1991). The concentration of DNA was determined by absorption at 260 nm. The purity of DNA was estimated by the ratio of absorp tion at 260 nm to that at 280 nm. Only DNA preparations with a ratio >1.6 were used for transformation.
PCR amplification of the ?OB gene: Segments of the rpoB gene were PCR-amplified from some donor strains for the purpose of being used as donor molecules in transformation. From each such donor, a 2351- and a 3367-bp fragment of the ?OB gene were amplified using genomic DNA as a template. Primers were based on the sequence of the ?OB gene of strain 168 (sequence kindly provided by KATHRYN BOOR, MARIAN
DUNCAN, and CHESTER PRICE). For the 3367-bp fragment, primers extended from 1102-1 125 bp (5”AGGTCAACTAGT- TCAGTATGG ACG3’) for the leading strand and from 4468- 4445 bp (5’-AACACCTGGTTCAGGAACATTGTC-3‘) for the lagging strand. For the 2351-bp fragment, the primers ex- tended from 1102- 1125 bp (the same primer as for the 3367- bp fragment) for the leading strand and from 3452-3435 bp (5’-CATCATCCTTCACTAGGC-3‘) for the lagging strand. Primers were synthesized by Oligos, etc. (Guilford, CT).
PCR was performed according to the GeneAmp protocol (Perkin-Elmer-Cetus) in 100 pl volume reactions of PCR buffer (50 mM KC], 10 mM Tris-HC1, 2 mM MgC12, and 0.1 mg/ml bactogelatin, at pH 8.3), with the following final con- centration of reagents: 200 p~ each dNTP, 1 mM each primer, 2.5 units AmpliTaq DNA polymerase (Cetus), and 0.01 pg template DNA. The PCR conditions involved denaturation at 94” for 30 sec, annealing at 52” for 45 sec, and extension at 70” for 3.5 min for the 3367-bp fragment or 2.5 min for the 2351-bp fragment. PCR was run for 40 cycles. It was previously shown that genomic DNA used as a template in PCR may contribute a very small amount to transformation when PCR- amplified DNA is used as donor (ROBERTS and COHAN 1993). To reduce further the contribution of genomic DNA in trans- formation, we used the PCR-amplified ?OB fragment as a template for a second PCR reaction. The DNA from this sec- ond round of PCR was used as donor DNA for transformation.
Restriction-digest analysis of sequence divergence at +OB:
The 3367-bp segment of rpoB that was PCR-amplified from laboratory strain YB886 was digested by four 4cutter restric- tion endonucleases (DpnII, H i n f f , MspI, and IzsaI), and the digests were assayed by electrophoresis as described by ROE ERTS and COHAN (1993). Restriction digest patterns from other strains were taken from data reported by ROBERTS and COW (1993, 1995). We inferred the presence and absence of restriction sites from restriction digest patterns using the computer algorithm of COHAN et al. (1991). Sequence diver- gence between pairs of strains was inferred from the number of shared and nonshared restriction sites, following equation 5.50 of NEI (1987).
Transformation: Recipient strains of B. subtilis and B. moja- vensis were induced to become competent and were trans- formed toward rifampicin resistance with 10 pg/ml of g e n e mic DNA extracted from rifR mutants of several strains, El, DV6-A-2-3, DV7-B-4, DV4D-4, DV4”2, DV4A-5, DV2-
TABLE 1
Strains used in this s t u d y
Strain" Origin
~~
Restriction
Genotype activity
B. subtilis 168 group 1A2 YB886 BG101" BG119" BG123" BG117" BG126" BG125" BG128" BG107" R0-J-2g
R S - G 4 h
W E - 2 RO-A4 RO-NN-1 RO-00-2 RO-GG2 RO-CG1 W23 group RO-HH-1 RO-DD-2 2A2 RO-E-2 DV3-G2' DV3-B-1 DV6A-2-3 DV7-B4 DV4D-4 DV4C-2 DV4A-5 DV2-D-1 DV2-F-1 DV2-B-2
B. mojavensis IM-A-3 12' IM-G45 IM-F-1 RO-H-1 RO-QQ2 RO-C-2
B. atrqbhaeus NRRL N W 2 1 3 B. amyloliquefaciens
ATCC 23350
B. lichenijbrmis ATCC 14580
BGSC~
J.
c.
ALONSOJ. C. h o ~ s o J.
c.
ALONSOJ.
c.
ALONSOJ. C. ALONso
J.
c.
h o ~ s o J.c.
ALONSOJ. C. ALONso
J.
c.
ALONSOF. M. COW, from soil F. M. COW, from soil F. M. COW, from soil F. M. COW, from soil F. M. COHAN, from soil F. M. COW, from soil F. M. COW, from soil F. M. COW, from soil
F. M. COW, from soil
F. M. COW, from soil
BGSC
F. M. COW, from soil
F. M. COW, from soil
F. M. COW, from soil F. M. COW, from soil F. M. COW, from soil F. M. COW, from soil F. M. COW, from soil
F. M. COW, from soil F. M. COHAN, from soil
F. M. COW, from soil F. M. COW, from soil
F. M. COW, from soil
F. M. COW, from soil F. M. COW, from soil
F. M. COW, from soil F. M. COW, from soil F. M. COW, from soil
A.R.s.c.c.~
ATCC'
ATCC
W.LC
ret+, trpC2, metB5, amyE, sigB,
recPl49 ( y ) I recH342 ( 7 )
recG40 ( E )
recB3 ( E )
addB72
(0)
recRl (a) recL16 (a) W.t.
W.t. W.t. W.t. W.t. w. t. w. t. w. t. w. t. w. t.
xin-1, attSPP
add45
(0)
W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. W.t. No ND* ND ND ND ND ND ND ND ND No No No Yes No No No No No No No No No No Yes No Yes No No Yes No No No No No No Yes No ND ND ND
a Strains isolated by F. M. COW were classified into species and subspecific groups by ROBERTS and COW
(1993, 1995).
'Wild type.
*
Not determined."Strains are isogenic with YB886 with the exception of the mutation indicated. 'Greek letter indicates recombination pathway affected by mutation.
g Strains with "RO" prefix were collected from soil near Rosamond, CA, in the Mojave Desert.
Strains with "RS" prefix were collected from soil near Rosamond, CA, in the Mojave Desert. ' Strains with "DV" prefix were collected from soil in Death Valley, C A .
I Strains with "IM" prefix were collected from soil near Hohhat, Inner Mongolia, China, in the Gobi Desert.
Agricultural Research Service Culture Collection at the National Center for Agricultural Utilization Service. 'American Type Culture Collection.
including the recipient (following COHAN et al. 1991). Each recipient strain was transformed with DNA from several do- nors representing different levels of sequence divergence from the recipient.
Frequencies of transformation are reported throughout as
the fraction of colony-forming units that were resistant to rifampicin, after accounting for spontaneous mutation toward rifampicin resistance (occurring in the absence of DNA at a frequency of 2.1 X
To test the effect of the concentration of donor DNA on the relationship between sexual isolation and sequence diver- gence, B. subtilis strain 1A2 was transformed with four concen- trations of genomic DNA (10, 1, 0.1, and 0.01 pg/ml). To test the effect of donor segment size, strain 1A2 was trans formed with PCR-amplified ?OB segments of two sizes (2351
and 3366 bp).
To test the effect of intracellular competition between exoge-
nous recipient- and donorderived DNA, the recipient strain
1A2 was presented with a mixture of rifampicin-resistant DNA
from a test donor and rifampicin-sensitive (rif’) DNA from the recipient, at a saturating concentration (10 pg/ml). To the extent that the mixed DNA transformed the recipient toward rifR at a lower frequency than did pure DNA from the test donor (presented at the same total concentration), we con- cluded that competition by the recipientderived rifS DNA had reduced heterogamic transformation. The DNA presented was either the 3.4kb PCR-amplified segment of ?OB or genomic
DNA. DNA from each strain was presented at 10 pg/ml (well
above saturating), so that DNA from both sources could enter many of the competent cells. We found that transformation with PCR-amplified segments of recipientderived
rif’
?OB caused an increase in mutation toward rifampicin resistance: the elevated mutation rate was 1.2 X lo-’ in the presence ofrecipientderived rifS DNA, compared to a normal mutation rate of 2.1 X
lo-*
in the absence of DNA. The transformation rates were corrected for this mutagenic effect of DNA.RESULTS
Sequence divergence between recipients and do-
nors: Sequence divergence between recipients and do- nors at the gene ?OB was estimated from restriction digest analyses of the 3367-bp PCR-amplified segment of $OB (restriction data for most strains were taken from Table 1 of ROBERTS and COHAN 1993). The 3367- bp segments of some recipient strains, including the laboratory strain YB886 and six wild isolates (RO-J-2, RSG4, RSE-2 of B. subtilis, and IM-A-312, IM-G45, and IM-F-1 of B. mojavensis), were not included in ROBERTS and COHAN’S (1993) analysis. We found that restriction of the 3367 bp segment amplified from strain YB886 yielded the same pattern of restriction fragments as did 1A2, so sequence divergence between YB886 and donors is the same as that for strain 1A2. To estimate the divergence between the donors and the remaining six recipients, we used restriction digest data for an 808- bp segment located within the 3367-bp segment (808- bp data reported by ROBERTS and COHAN 1995). We noted that for this 808bp segment, each of the above six recipient strains was identical in restriction digest pattern with at least one strain in ROBERTS and C o w ’ s
(1993) Table 1: B. subtilis strains RO-J-2, RS-(X, and RSE-2 were identical in restriction pattern to strain RO- A-4; B. mojavensis strain IM-A-312 was identical in pat-
tern to strain RO-G2, and B. mojavensis strains IM-(2-45 and IM-F-1 were identical in pattern to strain RO-H- 1 (ROBERTS and COHAN 1995). We assumed that the divergence at the 3367-bp segment between each of these six strains and a given donor was the same as that for strains showing the same digest pattern for the 808- bp fragment. This assumption is justified because diver- gence levels at the 8 0 8 and 3367-bp segments are very highly correlated: for each of the five B. subtilis strains
(1A2, RO-A-4, RO-NN-1, 2A2, and RO-E-2) and the three B. mojavensis strains (RO-G2, RO-H-1, and RO-
Q Q 2 )
for which we have both 808-bp (ROBERTS and COHAN 1995) and 3367-bp (ROBERTS and COHAN 1993) data, the divergence from the donors of this study at the 808bp segment is highly correlated with divergence at the 3367-bp segment ( r = 0.99, 0.99, 0.99, 0.97, 0.97, 0.98, 0.98, and 0.99 for the eight strains listed above, respectively; N = 6 sequence clusters of donors used in this study). The high degree of correlation is consistent with the very low rate of recombination found within B. subtilis and B. mojavensis (ROBERTS and COHAN 1995).Estimates of sequence divergence between recipients and donors are listed in Table
2.
Effect of the genotype of the recipient on the relationship between sexual isolation
and
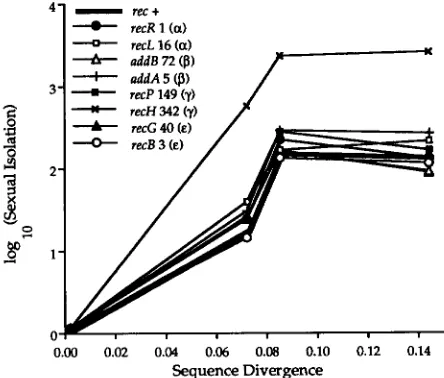
sequence divergenceEffect of recombination mutants on the sensitivity of sexual isolation to sequence divergence: The B. subtilis laboratory strain YB886 and eight isogenic mutants for recombination enzymes were transformed by a homo- gamic donor (a rifR mutant of strain 1A2) and by rifR mutants of the type strains of B. atrophaeus, B. amylolique- faciens, and B. lichenifomis. Whereas sexual isolation and sequence divergence were earlier shown to be log-lin- early related for recipient strain 1A2 (ROBERTS and Co-
HAN 1993), the relationship was decidedly not log linear for “wild-type” laboratory strain YB886 and for each of its mutant substrains (Figure 1; Table 3). Instead, these strains reached a plateau of sexual isolation at 8% se- quence divergence (with B. amyloliquefaciens as donor), with no further increase at 14% divergence (with B. lichenifwmis as donor) (paired t-test of the average dif- ference between the number of transformants with B. amyloliquefaciens and B. lichenifomis as donors: t =
1.77,
ns., N = 9 substrains of YB886).TABLE 2
Sequence divergence between recipient strains of B. subtilis and B. rnojavemis and donors
of different sequence similarity groups
B. subtilis
B. atrophaeus B. amyloliquefaciens B. lichenijbmis Recipient 168 group” W23 groupb B. mojavensis‘ NRS213 14580 ATCC 23350 ATCC
B. subtilis 0.003 t 0.002 0.031 t 0.000 0.046 2 0.000 0.070 0.083 0.142 B. mqavensis 0.049 2 0.002 0.044 2 0.000 0.005 2 0.002 0.107 0.112 0.167
All recipient strains from the same species showed the same sequence divergence from each group of donors. In those cases where we used more than one strain as donor from a sequence-similarity group (B. mojavensis and the 168 and W23 groups of
B. subtilis), we have listed the average sequence divergence between the group and the recipient, followed by the standard deviation among the members of the donor group.
a Donor strains include 1A2, RO-A-4, and RO-NN-1.
‘Donor strains include RO-G2, RO-H-1, and RO-QQ-2. Donor strains include 2A2 and RO-E-2.
pared with the wild type (average difference in loglo- transformed sexual isolation between recHand wild type was 1.31, yielding
=
21).Two of the eight mutations affected the level of com- petence for transformation (as measured by the fre- quency of homogamic transformation). These muta- tions included recPand recB. Note that the strong effect of recH on sexual isolation was not accompanied by a change in competence.
Assays of restriction endonuclease activity: We tested 10 B. subtilis strains for the ability to restrict lambda DNA, and three of these strains tested positive for re- striction activity. Two of these strains (DVGA-2-3 and DV4D-4) yielded the same pattern of restriction as
*
recE3W0.00 0.02 0.04 0.06 0.08 0.10 0.12 0.14 Sequence Divergence
FIGURE 1.-The relationship between sexual isolation and sequence divergence, for wild-type 1aboratoIy strain YB886 (a derivative of strain 168) and various isogenic substrains mu-
tant for recombination enzymes. The recombination pathway affected by each mutation is indicated in parentheses. Strain YB886 is indicated in bold line. The sequence divergence data are from Table 2, and the sexual isolation data are from Table 3. The figure includes a point for homogamic transformation (i.e., transformation by a rifR mutant of 1A2, another deriva- tive of strain 168), which by definition is at 0 sequence diver- gence and zero sexual isolation (log transformed).
strain RO-QQ2, suggesting that the endonuclease was BsuRI, an isoschizomer of Hue I11 (COW et al. 1991; WILSON 1991). The third strain, DV2-D-1 yielded seven restriction fragments ranging in size from 4 to 20 kb, consistent with restriction by a six-cutter endonuclease. COHAN and associates (1991) had earlier found restric- tion activity in two of 22 B. subtilis strains, so our best estimate of the frequency of restricting strains within B. subtilis is 16% (5/32); they also found that one in five B. mojavensis strains had restriction activity, yielding a frequency of 20% in this species. Because restricting strains constitute a sizeable fraction of strains in these species, it is important to investigate the effect of restric- tion on the sensitivity of sexual isolation to sequence divergence, as we discuss below.
The relationship between sexual isolation and se- quence divergence for restricting and nonrestricting strains: We investigated the effect of restriction activity on sexual isolation by comparing the relationship be- tween sexual isolation and sequence divergence for re- stricting and nonrestricting wild-type strains.
We measured the competence of the six wild isolates of B. subtilis and B. mojavensis with restriction activity by assaying the frequency of homogamic transformation toward rifampicin resistance for each strain. Strain RO- A-4 of B. subtilis and strain RO-QQ2 of B. mojauensis were transformed at frequencies that were orders of magnitude greater than mutation toward rifampicin re- sistance (frequency of mutation was 2.1 x lop8). The other four strains were transformed toward rifampicin resistance at frequencies very close to that of mutation; so these strains were excluded from further study.
TABLE 3
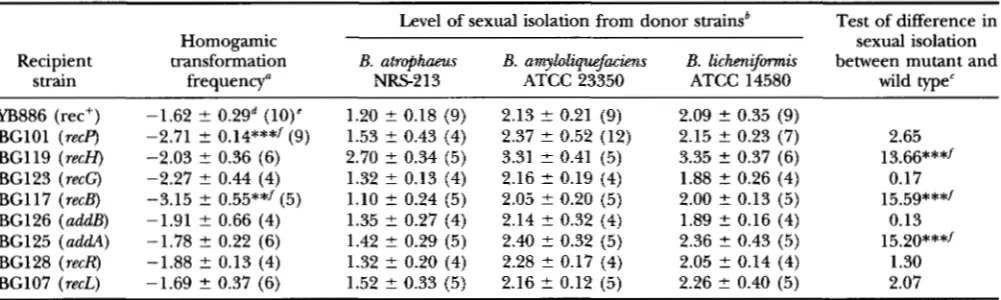
Transformation frequencies and levels of sexual isolation from other species, for recombination mutants of B. subtilis
Level of sexual isolation from donor strainsb Test of difference in
Recipient transformation B. atrophaeus B. amyloliquefacims B. lichaijimis between mutant and
Homogamic sexual isolation
strain frequency" NRS2 13 ATCC 23350 ATCC 14580 type" wild
YB886 (rec+) -1.62 2 0.2gd (10)' 1.20 2 0.18 (9) 2.13 ? 0.21 (9) 2.09 f. 0.35 (9)
BGlOl (recp) -2.71 2 0.14***/(9) 1.53 t 0.43 (4) 2.37 2 0.52 (12) 2.15 2 0.23 (7) 2.65 BG119 (recH) -2.03 ? 0.36 (6) 2.70 2 0.34 (5) 3.31 2 0.41 (5) 3.35 2 0.37 (6) 13.66***/ BG123 (recG) -2.27 2 0.44 (4) 1.32 2 0.13 (4) 2.16 2 0.19 (4) 1.88 f. 0.26 (4) 0.17 BG117 (recB) -3.15 2 0.55**/(5) 1.10 2 0.24 (5) 2.05 2 0.20 (5) 2.00 +. 0.13 (5) 15.59***/ BG126 (addB) -1.91 2 0.66 (4) 1.35 2 0.27 (4) 2.14 2 0.32 (4) 1.89 2 0.16 (4) 0.13 BG125 ( a d d ) -1.78 5 0.22 (6) 1.42 5 0.29 (5) 2.40 5 0.32 (5) 2.36 5 0.43 (5) 15.20***/ BG128 (re&) -1.88 2 0.13 (4) 1.32 5 0.20 (4) 2.28 2 0.17 (4) 2.05 2 0.14 (4) 1.30 BG107 (recL) -1.69 5 0.37 (6) 1.52 2 0.33 (5) 2.16 L 0.12 (5) 2.26 +. 0.40 (5) 2.07
a Frequency at which a strain was transformed by DNA from its own mutant substrain. Frequencies were loglo-transformed.
'Factor by which a recipient's transformation frequency was reduced, compared with homogamic transformation. Sexual
t value from a paired t-test of the difference between the mutant and wild type (YB886), for the average difference in loglo-
Standard deviation among experimental trials.
The mean of each mutant was compared to that of the isogenic wild type (YB886) in a t-test.
isolation values were loglo-transformed.
transformed sexual isolation values, over the three donors ( N = 3 donors).
"The numbers of experimental trials.
/ t test significant at P < 0.05, after adjustment for multiple comparisons with a sequential Bonferroni test.
*
P < 0.05,**
P< 0.01,
***
P < 0.001.an average datum for sequence divergence). We ex- cluded from this regression analysis sexual isolation be- tween the homogamic donor (i.e., the rifR mutant de- rived from the same strain as the recipient). This point would by definition be 7r = 0 and loglop = 0. Excluding the homogamic donor allowed us to analyze the possi- bility of sexual isolation among strains with near-zero sequence divergence (in this case, the possibility of sex- ual isolation caused by restriction activity).
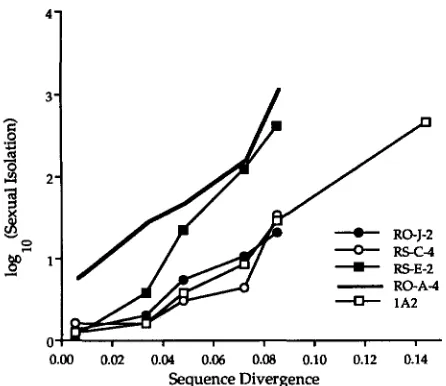
Strain RO-A4 of B. subtilis shows sexual isolation as a recipient even from its closest relatives within the species: the frequency of transformation by donor strains with very little sequence divergence from RO-A4 (i.e., other members of the 168 group of B. subtilis), but that lack the strain's modification activity (COHAN et al. 1991), is 5.2 times lower than when RO-A4 acts as its own donor (Table 4). This is in contrast to the nonrestricting wild- type strains of B. subtilis; these strains were transformed at the same rate with the DNA of close relatives as with their own DNA. These results corroborate earlier work of COHAN and associates (1991).
We next investigated whether the effect of sequence divergence on sexual isolation might be the same for restricting and nonrestricting strains. To this end, re- stricting strain RO-A4 and four nonrestricting strains of B. subtilis were transformed with rifR donors from the 168 and W23 groups of B. subtilis and from B. moja- vensis, B. a t r o p h m , and B. amyloliquefaciens (Table 4; Figure 2). Included among the four nonrestricting
strains was the laboratory strain 1A2, the wild-type deriv- ative of strain 168 (data for 1A2 from ROBERTS and COHAN 1993). The sensitivity of sexual isolation to se- quence divergence was quantified by the slope
4
of theregression of loglo(p) on 7r. The sensitivity parameter
4
was very similar for restricting and nonrestricting strains of B. subtilis, with the4
estimate of RO-A4(4
= 26.65) falling well within the range of the nonrestricting strains(4
ranging from 14.44 to 32.76; for strain 1A2,4
= 20.12 ? 1.74, when homogamic transformation is omitted from the calculation) (data from ROBERTS and COHAN 1993). Thus, while restriction activity causes sex- ual isolation, incremental increases in sequence diver- gence cause the same multiplicative increases in sexual isolation for the restricting strain tested and the nonre- stricting strains of B. subtilis.In contrast to the case of B. subtilis strain RO-A4, the restricting strain R w Q 2 of B. mqiavensis did not show sexual isolation from its close relatives (Table 4). COW
et al. (1991) also found lower sexual isolation caused by restriction activity of RO-QQ2 than by that of ROA4.
As was the case for the B. subtilis comparison, compar- ison of the restricting strain RO-QQ2 with the four nonrestricting strains of B. mojavensis showed similar
4
values for restricting and nonrestricting strains: the sensitivity parameter4
for RO-QQ-2(4
= 20.69) was within the range of those for nonrestricting strains(ranging from 16.79 to 23.53) (Figure 3).
Variation among wild-type strains for the relationship between sexual isolation and sequence divergence:
*
i
b-
a e ,
s o
o w
2 ;
iu
'
-gn s ;
2 9
y
.
:
'38
.E
'3 a
ST
.e cd
a, .9
VI
'I
.N$
s
*I
D
ad
3
g
3
0i!
2
e. Wi
s
Iri
x f,ad .e
w a
' I ) l d
$
g
Iri.3
O k
i i
.9 05 r t :
!i
z
g
.ea"
.s
4
-a &
3
.e
2
2
0
:
a 1ru 0 ;
9
6 c '
mt
F
#
M
.CI
t
8
rrj1 a
&
9
8
m*s
2
g
z
6. E 4 0 31 2
4 W2
s
._ 0w 1
8
H c
.$ .:
QF
g
p
??E
y
Ea
0-c)
.E
.5
p
.%
e
.e m Y V O C 3
y
y1 Z + c o-
:'s
$E
(a r d . 9 * a . c Y jw q w q q c q c " ? c " ? o
-
E b s g
+j
g
tl t l t l t I tI +I $ 1 tl t l
E
i' E 34
j
2
q y q c q '?qC?qcq .$
z c
m o m 0 d m 3 m m
< ' 3
3
E n 3- m m m * - ( ~ m m
.e
m * a m m m m m m
~ m m ~ m a o m o m - m m m m w
g
Q Z ? i
5 o h &$ 2
5
x
a, 0 M $
8
3
a
'2
22.3
e,2
a U . . c 12
1 O p $ 2m m m m m * * m
9
0 a.,s
z
&
9
c o d ,2
.em m m o w ( D m *
;
g m c cs u o
2
g
2
gg.2
+I t l tl tl $1 +I t I +I
g
Y Q1s
'S8
;
z - ? z z
x?!x
a
.y c E * 4 0 $E
g
,art:o m e 2
E a 2i ? g
s
m m m m m * * m *
$
g . s i u Z a
. _ ^"_^_" a, 1 . 5 3
8
;
z z z z
5 c . c c c
g
5 u g ;
g a ,
o m m m m * m ( D *
5
c s ; " ,+I +I +I t l +I t l t l tl tl g
" iu a
9
o e rt: gq 5 9 1 " 1 L n q 9 1
E
. g L ' c t d = o o m$
2o o m m m + + m m g 6 2 0 3 '3 0
' 5
E
E 5 . 9 a c" h m ? - . n n G Z
E
g z g g
3
4
& & & &
& & s & g
g
c a , * e , 3:
w v v v w v v v % g g 5 c 1 % o o m r o m + m o c o Frr u , y . z e 8 0 0 0 0 00 0 0 0 0 . 5 s E $ . q $
g
+I +I +I +I t l t i +I t I tl
e
c"
&
$5
1b H m * a m ( D + o o
$ 5 ; 5 a g
E
a0 0 " ~~~~~ 2 e o s - E ;
$
2
hh hh h c a , + a c ; ;
g
.g
m G-q
2 C , . S . d ?
;
.4
eseg
s
c . C E . 5 a h o - 2 gp z z
p g k z
m mg z 2 g s 5
m b J e - 3S
e.s
F
tl tl tl +I tl t l tl- 3
3-
8
I c a E .El 'Ca,..e 2 $
2 2 2 2
2 2 2
g E . a l C _ o &
L , 3 ub
grt:,:
Y
e , " o a,5
g td<
5 5 b , 3 02
a,E
h- -
Z a gZ&-G;;M.
G - z z z G -
Q E m $ 2 2 a f - g
e,h 0 5
E
" "e g s g s
s S g
E E 1 g a U E Z e , p c 9 9 9 o u V Y i c 9 9 - 1; $ . z
2 s
u & a '30 0 0 0 0 0 0 0 0 h E t d e b 5 s u
3
t l t l t I tl t l +I t l +I +I
g
2
g
5
a
c a, 5 .Yo y o q
c"?cqoew
sa,+c
0 0 0 00 0 0 0 0
3gC&;2?
x3 d
s
$ 5 2
;
2
e,s
h " h n o h hh
,.S-z
3 a $ d $.",
,2 2 2 %
z z z s z
m
*
m m (0 0 m m 00h $ 2 : . p a
S a
m . 2 " w " y q ( 4 q y ys 4 . 5 -
E c0 0 0 0 0 0 0 0 0 tda g
5 -
td 0 3 . 3E . %
" h h hh"
^ ^ "
" 3 4 "" ~~"~ % 1 m 3 2.
h U 3
8
v v v v ""
p 1 y ~ m t - m ( ~ n
0 0 0 0 0 0 0 0 2 e,
( D a m * bema U
h hh h " h h h
2
r
5
G ? G ? - ! ' ? ? " ? @ ! ' : M Q c l o
0 0 0 0 0 0 0 0 0 2
$ 4 2 2
s
w m m t - a m m m c D
( D a m - m a m i
h
d o x a
c o o y o O o y " 0 .&.I
'410"cq 9 9 1 9 9
-
g c
8 4 3
Yw w ( D o o 3
~~
3 o m m
v
Q1 0 s -
*LC)+* (D m m
1 c * i u
v w v
~ I "
m m m L b m * m o Q h c G a , c 0 0
a 2 0 5 3
-
2
b * O N m m m w m
h 3
';,td C m . Z ;;i
g
E5 - C
Ys!
t l +I +I +I +I +I $1 t l tI
%?Z!
$$?$X
c,
3
;
;,t
u .9k
5e
:
Z h m E o e $ q = + h h 1 E o a z +
I I I I I I I I I
E 2 2 . g ' s . g
Z
e
o nE , s
c4 :2
"E
;
g
.E
& O e, Ed m
T z
y e , e , oe,:34e;,
2
_ % b ' ? . [ ~ ~ ~ ~ $
e E s $ n $ ~ $ $ l M O
O - - o ~ ~ - E E m c E a , u
$ $ &
E Z Z Z & &
e g ? c E c " , V 1 ; , % , g Zm m * w m m m a *
h 5 Y a,
2
m c +.e x * O O $"-c RO-J-2 RSE-2
-
RO-A-40
0.00 0.06 0.02 0.04 0.08 0.10 0.12 0.14 Sequence Divergence
FIGURE 2.-The relationship between sexual isolation and sequence divergence, for wild-type strains of Bacillus subtilis.
The bold line represents a strain with restriction activity. The sequence data are from Table 2, and the sexual isolation data are from Table 4. Each point represents the average sequence divergence and sexual isolation between a recipient and all the donor strains tested from a sequence cluster. This figure does not include a point for homogamic transformation.
was too low to yield a reliable estimate of the transfor- mation rate with the more divergent B. lichenqormis as donor. However, we were able to determine that the rates of transformation with B. l i c h i j i m i s (14.2% di- vergent from B. subtilis recipients and 16.7% divergent from B. mojavensis recipients) as donor was significantly less than that with B. amyloliguefaciens (8.3% divergent from B. subtilis recipients and 11.2% divergent from B. mojavensis recipients; paired t-test of the average differ- ence between the numbers of transformants with B. arnyloliqwfaims and B. lichifornis as donors: t = 6.19, P
<
0.001, N = 9 strains). This is consistent with the log-linear relationship between sexual isolation and se- quence divergence extending to the level of 14% se- quence divergence, as was found with strain 1A2 asrecipient (ROBERTS and COHAN 1993) and is inconsis- tent with the pattern found for strain YB886.
As indicated earlier, the wild-type strains of B. subtilis showed over a twofold spread in the sensitivity parame- ter
4.
An analysis of covariance indicated that these strains were significantly heterogeneous for4
(F4,16 = 4.94, P<
0.01). This heterogeneity was not due to the restricting strain RO-A-4, as an analysis of covariance with this strain excluded also showed significant hetero- geneity (Fs,13 = 5.76, P<
0.01).The wild-type strains of B. mojavensis were much more homogeneous for the sensitivity parameter, with
4
rang- ing only from 16.8 to 23.5. An analysis of covariance showed no heterogeneity among strains (F4,12<
1).Effect of the conditions of transformation on the relationship between sexual isolation and sequence divergence
The following set of experiments tests the effects of the conditions of transformation on the relationship
IM-A312
"c I"c-45
*
IM-PI-
x) 0.02 0.04 0.06 0.08 0.10 0.12 Sequence Divergence
FIGURE 3.-The relationship between sexual isolation and sequence divergence, for wild-type strains of B. mojauensis. The bold line represents a strain with restriction activity. The se- quence data are from Table 2, and the sexual isolation data are from Table 4. This figure has the same format as Figure 3.
between sexual isolation and sequence divergence, us- ing B. subtilis laboratory strain 1A2 as the recipient.
Effect of donor fragment length on the relationship between sexual isolation and sequence divergence:
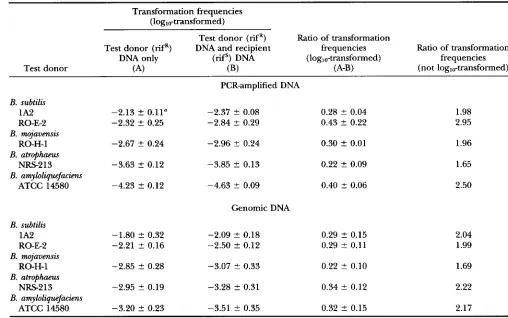
Strain 1A2 of B. subtilis was transformed with PCR-ampli- fied
?OB
segments, of size 3367 and 2351 bp, from a set of donors representing different levels of sequence divergence from the recipient (Table 5). Transforma- tion rates were, on average, 10 times lower in transfor- mation with 2351-bp segments than with 3367-bp seg- ments (mean difference between loglo-transformed frequencies of transformation with 2351- and 3367-bp fragments was 0.99 ? 0.23, P<
0.05, N =4
donors). The decreased transformation rate with smaller frag- ments of DNA is consistent with earlier work (MOR- RISON and GUILD 1971).The recipient strain was slightly more sensitive to se- quence divergence when presented the shorter 2351- bp segments than when transformed with 3367-bp seg- ments: the slopes of the relationship between sexual isolation and sequence divergence were 27.76 for the 2351-bp fragment and 20.78 for the 336Gbp fragment. However, an analysis of covariance showed no heteroge- neity of slopes between the two size treatments (F1,4
<
1; Figure 4 and Table 5).TABLE 5
Transformation frequencies and levels of sexual isolation in transformation
with the 2351- and the 3367-bp PCR-amplified segments
Sexual isolation from test donors'
Homogamic B. Sensitivity
( 9 )
of sexualFragment transformation B. mojavensis B. astrophaew amyloliguefaciens isolation to sequence
length (bp) frequency" RO-H-1 NRRL-NRS-213 ATCG23350 divergence'
3367 -2.26 +- 0.33d (18)" 0.27 ? 0.38 (11) 1.18 ? 0.55 (5) 1.82
+-
0.31 (5) 20.82 2 6.78 2351 -3.07 2 0.03d (4)' 0.30 ? 0.06 (4) 1.60 ? 0.02 (4) 2.41+-
0.07 (4) 27.82 ? 9.45 Segments are of @OB, with B. subtilis strain 1A2 as recipient. Concentration of DNA was 10 pg/ml."
standard error presented is the standard error of the slope.
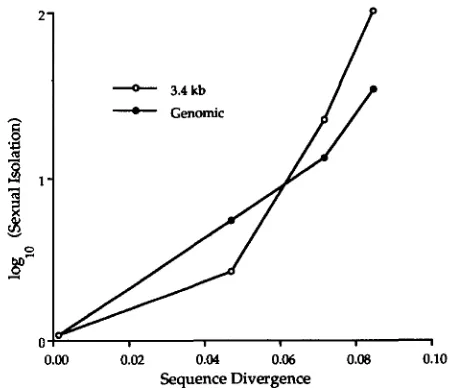
'
Standard deviation among experimental trials. Numbers of experimental trials performed.presented in this experiment at 0.1 pg/ml for both genomic and PCR-amplified donor DNA. In this com- parison, the longer donor molecules, as above, brought about a lower sensitivity to sequence divergence (with sensitivity slopes of 22.70 for the 3.4kb segment and 17.30 for genomic DNA), although the difference was not significant in an analysis of covariance (F1,4
<
1; Figure 5 and Table 6).Effect of donor DNA concentration on the relation-
ship between sequence divergence and sexual isola- tion: We hypothesized that high concentrations of d e nor DNA might saturate the mismatch repair system and perhaps alter the sensitivity of transformation to sequence divergence. We transformed B. subtilis strain lA2 with four concentrations of genomic DNA 10 pg/
a Frequency at which strain 1A2 was transformed by DNA from its own mutant substrain. Frequencies were loglo-transformed.
bFactor by which transformation frequency was reduced, compared with homogamic transformation, when strain 1A2 was
'Sensitivitv was measured as the sloDe of the repression of log-transformed sexual isolation on sequence divergence. The transformed with DNA from other strains. Sexual isolation values were loglo-transformed.
ml (above saturation, which is -3 pg/ml) (M. S. ROE
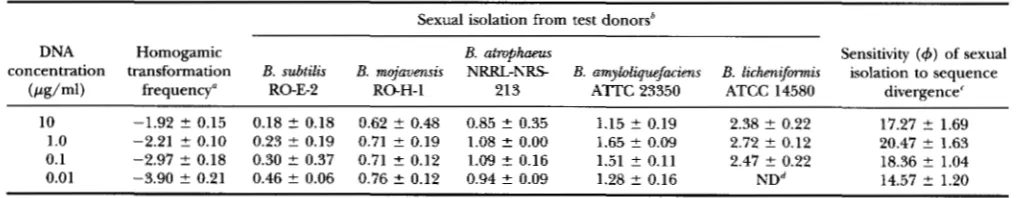
ERTS and F. M. C o w , unpublished results), 1 pg/ml (below saturation, but high enough to allow con- gression), 0.1 pg/ml and 0.01 pg/ml (too low to allow congression, so that only a single DNA molecule enters each cell) (CUTTING and VANDER HORN 1990). Trans- formation rates were on average 11
7
times higher with 10 pg/ml donor DNA than with 0.01 pg/ml (Table7).
Slopes of the log-linear relationship between sexual isolation and sequence divergence were very similar across DNA-concentration treatments, with values rang- ing from 20.47 for 1 pg/ml to 17.27 for 10 pg/ml (anal- ysis of covariance: F3,16
<
1; Table7
and Figure 6). Thus, donor DNA concentrations do not affect the relation- ship between sexual isolation and sequence divergence.-
2.4kbP
-
3.4kb0.02 0.04 0.06 0.0s 0.10
-
3.4kb//
0:02 0:06 0.0s 0:10
Sequence Divergence
FIGURE 4.-The relationship between sexual isolation and sequence divergence with transformation by the 2 . 4 and 3.4 kb PCR-amplified segments of the @OB gene. DNA was pre- sented at a concentration of 10 pg/ml. The sequence data are from Table 2, and the sexual isolation data are from Table 5. The figure includes a point for homogamic transformation, which is by definition at 0 sequence divergence and 0 sexual isolation (log transformed).
0.00 0.02 0.04 0.06 0.08 0.10
Sequence Divergence
TABLE 6
Transformation frequencies and levels of sexual isolation in transformation with the 3367-bp PCR-amplified segment and with genomic DNA
Sexual isolation from test donorsb
Homogamic B. Sensitivity
( 4 )
of sexualDonor transformation B. mojavensis B. astrophaeus amyloliquefaciens isolation to sequence
fragment frequency" RO-H-1 NRRL-NRS-2 1 3 ATCG23350 divergence"
3367 bp -3.56 -C 0.03d (4)" 0.39 ? 0.50 (4) 1.33 ? 0.50 (4) 1.98 2 0.28 (4) 22.74 ? 6.38 Genomic DNA -2.97 2 0.15d (3)" 0.71 2 0.12 (3) 1.09 ? 0.16 (3) 1.51 ? 0.11 (3) 17.30 2 1.65
Segment is of ?OB, with B. subtilis strain 1A2 as recipient. Concentration of DNA was 0.1 pg/ml.
a Frequency at which strain 1A2 was transformed by DNA from its own mutant substrain. Frequencies were loglo-transformed.
bFactor by which transformation frequency was reduced, compared with homogamic transformation, when strain 1A2 was
Sensitivity was measured as the slope of the regression of log-transformed sexual isolation on sequence divergence. The
Standard deviation among experimental trials.
transformed with DNA from other strains. Sexual isolation values were loglo-transformed.
standard error presented is the standard error of the slope.
'Numbers of experimental trials performed.
Intracellular competition between exogenous recipi- ent- and donor-derived DNA: We tested whether the simultaneous uptake of DNA from recipient and donor strains may exaggerate the sensitivity of sexual isolation to sequence divergence. When competent cells were presented 10 yg/ml of a 1:l mixture of recipient-de- rived rifS DNA and testdonor-derived rifR DNA, the frequency of transformation toward rifR was consis- tently about one-half the frequency when cells were presented only donor-derived rifR DNA at the same concentration (Table 8). The average ratio of the fre- quency of transformation with test donor DNA only to that with mixed DNA was 2.21 for PCR-amplified DNA and 2.02 for genomic DNA, in each case the ratio was not significantly different from 2, the number expected if the rifR and rifS donors competed equally well for the homologous site on the chromosome (paired t-test of loglo-transformed ratio, testing against a null expecta- tion of loglo[2]: t = 0.64, ns., for PCR-amplified DNA, and t = 0.44, ns., for genomic DNA, in each case, N =
5 donors). There was a similar slope relating sexual isolation to sequence divergence for the testdonor- DNA-only and the mixed-DNA treatments (analysis of covariance: F1,6
<
1). We conclude that the sensitivity of transformation to sequence divergence is not exag- gerated by intracellular competition between divergent and nondivergent donor molecules.DISCUSSION
Sexual isolation ( p ) in Bacillus transformation was previously shown to be closely predicted as a log-linear function of sequence divergence (n): loglo(p) = c#m
+
a, where the slope4
represents the sensitivity of sexual isolation to sequence divergence (ROBERTS and COHAN 1993). In this previous work, the sensitivity parameter was estimated at6
= 18.87. Because this earlier work was based on only one strain as recipient and on only a limited diversity of transformation conditions, we have in the present study attempted to test the generality ofTABLE 7
Transformation frequencies and levels of sexual isolation, for strain 1A2 as recipient when transformed with different concentrations of genomic DNA
Sexual isolation from test donorsb
DNA Homogamic B. atrophaezls Sensitivity ( 4 ) of sexual
concentration transformation B. subtilis E. mojavmsis NRRLNRS E. amyloliquefacims B. lichaiformis isolation to sequence (Pg/ml) frequency" RO-E-2 RO-H-1 213 ATTC 23350 ATCC 14580 divergence'
10 -1.92 2 0.15 0.18 2 0.18 0.62 t 0.48 0.85 t 0.35 1.15 2 0.19 2.38 t 0.22 17.27 2 1.69 1.0 -2.21 2 0.10 0.23 2 0.19 0.71 2 0.19 1.08 2 0.00
0.1
1.65 2 0.09 2.72 2 0.12 20.47 2 1.63 0.01
-2.97 2 0.18 0.30 2 0.37 0.71 2 0.12 1.09 2 0.16 1.51 t 0.11 2.47 2 0.22 18.36 t 1.04 -3.90 2 0.21 0.46 2 0.06 0.76 2 0.12 0.94 2 0.09 1.28 2 0.16 N D ~ 14.57 2 1.20
Means and SD are based on three experimental trials.
Frequency at which strain 1A2 was transformed by its own DNA. Frequencies were loglo-transformed.
Factor by which recipient's transformation frequency was reduced, compared with homogamic transformation, when recipient
Sensitivity was measured as the slope of the regression of log-transformed sexual isolation on sequence divergence. The
Not determined because the rate of transformation was too close to that of mutation.
strain 1A2 was transformed by DNA from other strains. Sexual isolation values were loglo-transformed.
3 1
I
P'
-
lO.O)rg/d
7
-0-l . O p g / d
-
0.1 pg/d-
0.01 w / d0.00 0.02 0.04 0.06 0.08 0.10 0.12 0.14 0.16 Sequence Divergence
FIGURE 6.-The relationship between sexual isolation and sequence divergence in transformation with different concen- trations of genomic DNA. The sequence data are from Table 2, and the sexual isolation data are from Table 7. The figure includes a point for homogamic transformation, which is by definition at 0 sequence divergence and 0 sexual isolation (log transformed).
this relationship across a diversity of recipient strains and transformation conditions. We aimed to test the generality of two aspects of this relationship: that the relationship between sexual isolation and sequence di- vergence follows a log-linear form and that the sensitiv- ity parameter
4
is -20.Effect of recipient strains on the relationship between sexual isolation
and
sequence divergenceThe log-hear nature of the relationship between sex- ual isolation and sequence divergence: All strains tested as recipients in this study showed a good log- linear relationship 4 . 3 % sequence divergence for B. subtilis and 11.2% for B. mojavensis (with B. amyloligwfa- ciens and more closely related strains as donor). In the case of the nine naturally occurring strains of B. subtilis and
B.
mojavensis, the level of competence was too low to estimate accurately the sexual isolation with the most divergent donor tested (B. lichenijionnis, with 14.2% di- vergence from B. subtilis and 16.7% from B. mojavensis).
Nevertheless, these strains were transformed at a much lower level with B. lichenijiiis than with B. amyloliquefa- ciens DNA this is at least consistent with the log-linear relationship extending to 16.7% divergence.The laboratory strain YB886 showed a different pat- tern of sensitivity to sequence divergence. This strain and all its mutant substrains showed a good fit to a log- linear model up to 8.3% divergence, but the log-linear relationship broke down at 14.2% divergence: the fre- quency of transformation with the more divergent B. lichenijiiis as donor was indistinguishable from that with the less divergent B. amyloliguefaciens as donor (Fig- ure 1). The known mutations in yB886 (trpc'L, metB5,
amyE, sigB, xinl, attSPp) do not suggest a cause for the breakdown of the log-linear relationship between sexual isolation and sequence divergence. It is therefore surprising that strain yB886 has such a different pattern of sexual isolation from that for strain 1A2 (ROBERTS and COHAN 1993), because both strains are derivatives of strain 168.
Evolution appears to have proceeded in the direction from the log-linear pattern of 1A2 to the nonlog-linear pattern of YB886, because all the naturally occurring strains tested share the 1A2 pattern. This evolution to- ward a nonlog-linear pattern of sexual isolation was spontaneous (i.e., it did not involve artificial selection), and it occurred in the laboratory within two decades. It thus appears that Bacillus strains have the potential to evolve easily from the log-linear to a nonlog-linear pattern of sexual isolation. However, our sample of nine wild strains from two species suggests that this evolu- tionary change has not occurred in nature.
The effect of recombination mutants on the sensitiv- ity of sexual isolation to sequence divergence: Of the eight recombination mutants tested in this study, only three were significantly different from the isogenic wild- type strain (YB886) in their sensitivity to sequence diver- gence. Two of these mutants, recB and addA, were actu- ally quite similar to wild type in their levels of sensitivity to sequence divergence (with a 19% reduction and a 79% increase in sexual isolation, respectively, compared with wild type). The mutation in recH, however, caused much greater sensitivity to sequence divergence than wild type (with a 21-fold increase in sexual isolation). The mutation in recH reduced the frequency of trans- formation by divergent donors without affecting the frequency of transformation by closely related donors. It is not clear how the mutation in recH would accom- plish this, although one possibility is that the recffprod- uct normally interacts with the mismatch repair system. In Salmonella, the mismatch repair enzymes coded by mutL, mutS, and mutH were shown to favor resolution of the heteroduplex (an intermediate step in recombi- nation, containing a recipient- and a donor-derived strand) toward the recipient-derived strand (RAYSSI- GUIER et al. 1989). Perhaps the recH product normally acts so as to enhance the activity of such mismatch repair enzymes.
The eight recombination mutants were chosen to represent the four pathways of recombination charac- terized by ALONSO et al. (1992), with two enzymes repre- sented per pathway. Mutations in two enzymes from the same pathway might have been expected to yield similar effects on sexual isolation. However, this was not the case for the two mutants of the y pathway. The muta- tion in recH increased the sensitivity of sexual isolation to sequence divergence, while the recP mutation had no effect. These results suggest that, at least with respect to the effects on sexual isolation, the recHand recPprod- ucts may belong to different recombination pathways.