ABSTRACT
ANGSTADT, ANDREA Y. Evaluation of the Genomic Aberrations in Canine Osteosarcoma and Their Resemblance to the Human Counterpart. (Under the direction of Dr. Matthew Breen).
In the last decade the domestic dog has emerged as an ideal biomedical model of complex genetic diseases such as cancers. Cancer in the dog occurs spontaneously and several studies have concluded that human and canine cancers have similar characteristics such as
presentation of disease, rate of metastases, genetic dysregulation, and survival rates. Furthermore, in the genomic era the dog genome was found more homologous in sequence conservation to humans than mice, making it a valuable model organism for genetic study in addition to pathophysiological analysis. Osteosarcoma (OS), the most commonly diagnosed malignant bone tumor in humans and dogs, is one such cancer that would benefit from
comparative genomic analysis. In humans, OS is a rare cancer diagnosed in fewer than 1,000 people per year in the USA, while in the domestic dog population the annual number of new cases is estimated to far exceed 10,000. This high rate of disease occurrence in dogs
of dog OS cases. Characterization of the affect that CNA has on the expression of select cancer associated genes revealed that imbalance and transcriptional dysregulation in canine OS also paralleled human OS. Specifically, changes in RUNX2, TUSC3, and PTEN
expression levels correlated with genomic copy number status in dog OS. This analysis showcased RUNX2 as an ‘OS associated gene’ and TUSC3 as a tumor suppressor gene involved in canine OS. In addition, direct comparison of genomic imbalance in human and dog OS using high resolution oligonucleotide aCGH indicated that the ‘OS associated genes’ RUNX2, CDKN2A/CDKN2B, MYC, RB1, and PTEN resided in orthologous microaberration regions (<500kb) with similar CNA patterns supporting that these genes are key genetic players driving OS progression. Similarities in genome-wide CNA patterns in OS between orthologous regions of the human and dog genome were also found suggesting that
Evaluation of the Genomic Aberrations in Canine Osteosarcoma and Their Resemblance to the Human Counterpart
by
Andrea Y. Angstadt
A dissertation submitted to the Graduate Faculty of North Carolina State University
in partial fulfillment of the requirements for the degree of
Doctor of Philosophy
Functional Genomics
Raleigh, North Carolina 2010
APPROVED BY:
_______________________________ ______________________________
Dr. Matthew Breen Dr. Dahlia M. Nielsen
Committee Chair
DEDICATION
BIOGRAPHY
Andrea developed her love for biology at an early age through hiking trips, raising farm animals, and grade school science fair projects. After graduating high school she went to Penn State University and completed a degree in animal bioscience with a minor in
ACKNOWLEDGMENTS
TABLE OF CONTENTS
LIST OF TABLES... viii
LIST OF FIGURES ...x
LIST OF ABBREVIATIONS ... xiii
Chapter I: Literature Review...1
Overview...2
Dog as a suitable model of human disease ...3
The pathology of osteosarcoma ...9
Molecular and genetic dysregulation of osteosarcoma...13
Genomic Studies ...17
Thesis Outline ...31
References...32
Chapter II: Characterization of canine osteosarcoma by array comparative genomic hybridization and qRT-PCR: Signatures of genomic imbalance in canine osteosarcoma parallels the human counterpart...49
Abstract...50
Introduction...51
Materials and Methods...53
Tissue Specimens...53
Array Comparative Genomic Hybridization (aCGH)...55
Fluorescence in-situ Hybridization (FISH)...56
Quantitative RT-PCR...57
Statistical Analysis...58
Results...59
Clinical Assessment ...59
Abundant Genomic Instability in Canine OS ...60
Breed and Morphological Subtype Specific Associated DNA Copy Number Aberrations
...68
qRT-PCR...70
Discussion...71
Acknowledgements...78
References...90
Chapter III: A genome-wide approach to comparative oncology: High-resolution oligonucleotide aCGH of canine and human OS pinpoints communal microaberrations ...101
Abstract...102
Introduction...103
Materials and Methods...107
Tissue Specimens...107
Array Comparative Genomic Hybridization (aCGH)...109
Array Comparative Genomic Hybridization Analysis...110
Identification of Orthologous Canine and Human Regions...111
Results...112
Refining Regions of Genomic Aberration in Canine OS...112
Extensive Genomic Imbalance in Human OS ...117
Comparative Regions of Genomic Imbalance in Canine and Human OS ...121
Discussion...125
References...131
Chapter IV: Conclusion...141
Concluding remarks ...142
Future evaluation of the complexity in canine OS...147
Appendix I: Tackling the characterization of canine chromosomal breakpoints with an
integrated in-situ/in-silco approach: The canine PAR and PAB...159
Abstract...160
Introduction...160
Materials and Methods...162
Clone Selection ...162
Cytogenetic Evaluation...162
Computational Analysis of Overlapping BAC Clones ...164
Results...164
Cytogenetic interpretation of the canine PAR ...164
Computational Analysis of the PAB...164
Comparison of Canine and Human PAR-Specific Genes...165
Discussion...167
References...168
LIST OF TABLES
Chapter I:
Table 1: Tumor incidence rate (all sites) estimated in pet dogs by country and tumor
characteristics...7 Table 2: Previously defined molecular factors dysregulated in canine OS ...14 Table 3: Summary of the cytogenetic abnormalities associated with human OS...23 Table 4: Summary of highly recurrent genomic aberrations (≥30%) in 38 patients of canine OS indentified by low-resolution (10-20Mb) aCGH ...29 Table 5: Summary of copy number imbalances for known cancer-associated genes in 38 patients of canine OS identified by low-resolution (10-20Mb) aCGH...29
Chapter II:
Chapter III:
Table 1: Clinical summary of 23 canine OS tumor samples analyzed for DNA copy number aberrations (CNAs) on Agilent’s canine180,000 feature G3 SurePrint oligonucleotide aCGH platform...108 Table 2: Clinical summary of 15 human OS tumor samples analyzed for DNA copy number aberrations on Agilent’s 60,000 feature G3 SurePrint oligonucleotide aCGH platform...109 Table 3: Frequency of autosomal CNAs (gain and loss) identified by the ADM-2 algorithm with a threshold of 6 and log2 ratio cutoff of 0.2 in 23 cases of canine OS...116 Table 4: Frequency of CNAs (gain and loss) identified by the ADM-2 algorithm with a
LIST OF FIGURES
Chapter I:
Figure 1: Dog cancer incidence rates for females and males estimated from tumor specimens received by the ATR from local veterinarians in Genoa, Italy across the study period (1985-2002) for all cancers and selected cancer sites by calendar period...9 Figure 2: SKY analysis of human OS...20 Figure 3: FISH analysis of dog OS as directed by aCGH...30
Chapter II:
Figure 1: Frequency of copy number aberration in 123 canine OS samples ...62 Figure 2: Bivariate fit of CNA percent loss/gain for 123 canine OS cases by the physical size of the aberrations in Mb...63 Figure 3: Whole genome 1Mb aCGH profile of a 12 year old female Golden Retriever
diagnosed with osteoblastic OS located in the left proximal humerus, co-hybridized with DNA derived from a blood sample from the same patient ...67 Figure 4: Further characterization of copy number aberration occurrences across 1,066
regions of aberration commonality in the 38 dog autosomes ...69 Figure 5: Quantitative RT-PCR on 26 canine OS patients for six genes located in regions that had a high occurrence of CNAs ...71 Supplementary Figure 1: Kaplan-Meier survival curve comparing different treatment
Supplementary Figure 4: Graphical representation of the PCA performed to evaluate global differences in genome-wide DNA copy number aberrations evident in our five breed groups ...82 Supplementary Figure 5: Graphical representation of the PCA performed to evaluate global differences in genome-wide DNA copy number aberrations evident in our three
morphological subtypes ...83
Chapter III:
Figure 1: Array CGH profiles of OS12, an 8 year-old male Rottweiler with osteoblastic OS ...113 Figure 2: Frequency of copy number aberrations in 23 canine OS samples ...115 Figure 3: Box plots of frequency of autosomal CNAs (gain and loss) in 23 canine OS cases (array call resolution ~27kb) identified by the ADM-2 algorithm with a threshold of 6 and log2 ratio cutoff of 0.2...117 Figure 4: Array CGH profile (100kb resolution view) of HOS5, a 12-year old female with OS in the left distal femur ...118 Figure 5: Frequency of copy number aberrations in 15 human OS samples ...119 Figure 6: Box plots of frequencies of CNAs (gain and loss) in 15 human OS cases (array call resolution ~100kb) identified by the ADM-2 algorithm with a threshold of 6 and log2 ratio cutoff of 0.5...121 Figure 7: Canine orthologous regions of the human genome that demonstrate similar
aberration frequencies in 15 cases of human OS arrayed at ~100kb resolution and 23 canine OS cases arrayed at ~27kb resolution...122 Figure 8: Chromosome profiles of CNA frequencies of 23 canine OS cases (~27kb
resolution) and 15 human OS cases (~100kb resolution) ...124
Appendix I:
Figure 2: The sequence within BAC clones 172L08 and 397M10, canFam(chrX:6580407-6630000), searched against 6.2 million unassembled shotgun sequences read from a male poodle genome using BLASTn...165
Figure 3: Position of PAR genes in 1Mb from the p-arm telomere and their comparative status in human (NCBI Build 36.1), horse (equCab1), dog (canFam2), and mouse (NCBI Build 37) according to the UCSC genome browser (http://www.genome.ucsc.edu/) ...166
Appendix II:
Figure 1: Whole genome aCGH profile of an 8 year old male Great Pyrenees diagnosed with fibroblastic osteosarcoma located in the right proximal tibia co-hybridized with DNA derived from a blood sample from the same patient...171 Figure 2: Whole genome 1Mb aCGH profile of a 12 year old female Golden Retriever
LIST OF ABBREVIATIONS
OS-osteosarcoma
BAC-bacterial artificial chromosome CGH- comparative genome hybridization aCGH-array comparative genome hybridization FISH-fluorescence in-situ hybridization
SKY- spectral karyotyping mBAND-multicolor banding CNA-copy number aberration CFA-Canis Familiaris
HSA-Homo sapiens
BLAST-basic local alignment and search tool PAR- pseudoautosomal region
PAB- pseudoautosomal breakpoint
RB1-retinoblastoma 1
PTEN -phosphatase and tensin homolog
MYC-v-myc myelocytomatosis viral oncogene homolog (avian) RUNX2- runt-related transcription factor 2
TUSC3- tumor suppressor candidate 3
Chapter I
Overview
Cancer is a complex genetic disease, divided into over 100 subtypes. It presents with a wide range of symptoms that often result in extensive pain and suffering for an individual. Even in today’s society with the vast advances in health care it is still the second leading cause of death in humans, only topped by heart disease (Jemal et al. 2009). As cancer studies in humans are usually difficult because of ethical boundaries, selection of an appropriate model organism is extremely important. In the last decade the domestic dog (Canis familiaris, CFA) has emerged as an ideal model organism for the study of several human diseases including cancer because of strong anatomical, pathological, physiological, and genetic similarities (Paoloni and Khanna 2008).
In this thesis I took a genome-wide approach to identify, characterize, and directly compare genomic instability in canine and human OS. I sought to demonstrate the value of the dog as a model organism through the identification of parallels in cytogenetic
abnormalities in osteosarcoma (OS) while further evaluating and interpreting the disease aberrations in canines. I hypothesized that an array comparative genome hybridization (aCGH) study of the genomic imbalance at a higher resolution would reveal that the disease in the dog is as chaotic in nature as it is in the human counterpart. Evidence supporting this hypothesis is found in the previous low resolution aCGH (10-20Mb) study of canine OS, which concluded that many genomic regions were subject to copy number changes (Thomas et al. 2009). Thomas et al. (2009) also found that the pattern of copy number changes differed between dog breeds for specific cancer related genes within these regions. I
believed this pattern would continue in the high-resolution analysis, suggesting that genomic imbalances may be result of an individual’s genetic framework. In order to properly interpret OS abnormalities I originally had planned to combine 1Mb-resolution bacterial artificial chromosome (BAC) aCGH, fluorescence in-situ hybridization (FISH), and computational analysis to determine precise locations of chromosome breakpoint region within aberrant OS karyotypes. I demonstrated the success of this approach in Appendix I by characterizing the naturally occurring breakpoint present on all canine X chromosomes - the pseudoautosomal breakpoint (PAB) (Young et al. 2008). Due to technological advancements I was
subsequently able to enhance sections of this methodology by using oligonucleotide aCGH evaluation of both dog and human OS. Overall, this thesis demonstrates the translational potential of the dog as a model organism for human OS while further characterizing the chaotic genomic imbalance present in canine OS, enhancing our understanding of OS genetic abnormalities in man and his ‘best friend.’
Dog as a suitable biomedical model of human disease
human-based studies the use of an appropriate model organism for the study of human diseases is important. In the last decade the domestic dog (Canis familiaris) has emerged as an ideal model organism for the study of several human diseases. According to the
2009/2010 National Pet Owners Survey conducted by the American Pet Products
Manufacturers’ Association (APPMA) nearly half of US households (45.6%) are dog owners with the total number of pet dogs in the US estimated at approximately 77.5 million
(http://www.americanpetproducts.org/press_industrytrends.asp). These dogs are susceptible to over 450 spontaneously arising diseases and approximately 360 of these are analogous to human diseases (Parker and Ostrander 2005; Wayne and Ostrander 2007; Shearin and Ostrander 2010). In addition, a subset of these 77.5 million dogs is divided into over 350 distinct breeds worldwide (Shearin and Ostrander 2010) with 167 breeds recognized in the USA by the American Kennel Club (AKC, http://www.akc.org/).
Distinct dog breeds provide the most compelling reason to use the dog as a model for spontaneously occurring human diseases because of the unique population structure formed during the domestication and development of the modern day purebred dogs. Each breed is defined by specific behavioral and physical characteristics that were driven to exceptionally high frequency by population bottlenecks and strong artificial selection (Karlsson and Lindblad-Toh 2008). The process of each breed arising from a limited number of founders and frequent use of popular sires has caused genetic diversity in dogs to be largely between breeds with limited locus and disease heterogeneity within breeds (Ostrander and Kruglyak 2000; Lindblad-Toh et al. 2005; Shearin and Ostrander 2010). In addition, since most breeds are fewer than 200 years old they have long stretches of linkage disequilibrium (LD) and long haplotype blocks, reducing the overall number of markers and individuals compared with humans needed to investigate genetic variants associated with morphological and disease variation (Sutter and Ostrander 2004; Lindblad-Toh et al. 2005). In turn, this makes genetic studies in dogs theoretically simpler and more straightforward (Shearin and
geographically isolated human populations, such as those carried out in Finland or Iceland (Ostrander and Kruglyak 2000; Shearin and Ostrander 2010), as both environmental and genetic factors are somewhat controlled. Previous genome-wide analysis strategies have exploited the breed structure of dogs to investigate the genetic basis of specific traits in dogs. One study found an IGF-1 single nucleotide polymorphisms (SNP) haplotype suggestive of causing differences in body size in dogs (Sutter et al. 2007; Jones et al. 2008) and more recently 915 dogs from 80 domestic dog breeds were genotyped to discover 51 regions of the dog genome associated with phenotypic variation including average breed body size and external body dimensions (Boyko et al. 2010).
Domestic dog breeds have fascinated geneticists since the early 1900s when the first inherited characteristics and disorders such as coat color, hairlessness, barking, and
hemophilia were published in the Journal of Heredity (Little 1914; Wright 1917; Whitney 1929; Hutt et al. 1948). As technology advanced, incorporation of cytogenetic techniques were introduced to the understanding of the genetics behind dog phenotypes (Karlsson and Lindblad-Toh 2008). In 1997 the first genetic linkage map of the dog genome (Mellersh et al. 1997), containing 150 microsatellite markers was published. In 1999 the gene associated with canine narcolepsy was identified and intensified interest in the dog as a model for human disease (Lin et al. 1999). The next few years saw the development of chromosome-specific single locus FISH probes for dogs (Breen et al. 2001; Thomas et al. 2001a; Thomas et al. 2001b; Breen et al. 2004), a 1Mb-resolution radiation hybrid map of the canine genome (Guyon et al. 2003), and survey sequencing and comparative analysis of the dog genome (Kirkness et al. 2003). In 2005, the Canine Genome Sequencing Project (Lindblad-Toh et al. 2005) further enhanced canine research by initiating the development of state-of-the-art tools from which to map traits in the dog. The availability of an assortment of genetic tools are useful in assisting researchers in defining phenotypes, selecting sample sets, identifying trait loci through linkage, association or selection mapping, identifying gene function and
These tools also allow for the characterization of the genomic dysregulation that leads to disease phenotype in dogs (Lindblad-Toh et al. 2005; Karlsson and Lindblad-Toh 2008).
Although the dog had long been used as a model system for drug discovery and development research because of its similarities to human anatomy and physiology (Khanna et al. 2006), until the genomic age few had compared genetic disease similarities between dogs and humans. Analysis of the dog genome demonstrated that it was more homologous in sequence conservation to humans than mice, the foremost model for genetic studies in
mammals since the beginning of the twentieth century (Kirkness et al. 2003; Lindblad-Toh et al. 2005). The mouse has been one of the most widely used model organisms and does boast an impressive availability of experimental resources that have been useful when testing the effect of genetic manipulation on single genes or interactions of a subset of genes (Paigen 1995; Bucan and Abel 2002). Yet in contrast to other vertebrates such as dogs, the mouse is not very useful when attempting to identify putative mutations in human diseases that are polygenic in nature. Also diseases in mouse are often induced while diseases in dogs occur spontaneously over the course of their lifetime and include several diseases also seen in humans such as cancers, diabetes, heart disease, eye diseases, epilepsy, deafness and even psychiatric disease such as obsessive compulsive disorder (Overall 2000; Khanna et al. 2006; Gershwin 2007; Withrow and Vail 2007).
In particular, the unique genetic architecture and the availability of genomic tools make dogs an ideal genetic model for the study of human cancers. As an artifact of the strong selection that breeders have imposed over the years to produce populations of dogs with common morphological and behavioral traits (Shearin and Ostrander 2010) an
incidence rate of canine cancer, studies have shown that tumors occur at a frequency paralleling human neoplasms (Cadieu and Ostrander 2007; Breen and Modiano 2008). Studies conducted by European and North American veterinary cancer registries have sought to estimate the occurrence rate of spontaneous tumors in pet dogs (Table 1), but these studies should not be considered a complete representation of the occurrence of canine cancer as they have variable reference population. Although the Animal Tumor Registry (ATR) of Genoa, Italy did recently provide an accurate assessment of the occurrence rate of select canine cancers (Figure 1). The incidence rates for all canine cancers separated out for gender and for selected site-specific cancers by calendar period are shown in Figure 1 (A and B). The ATR found that all cancer incidences were three times higher in female dogs than in male dogs, but this difference can be attributed to the high rate of mammary cancer in females. Although there is variation in the incidence of canine cancer we can infer from these studies that in terms of epidemiology cancer is a common disease in dogs much like that seen in human populations.
Table 1: Tumor incidence rates (all sites) estimated in pet dogs by country and tumor characteristics (Merlo et al. 2008). aSize of the estimated canine population at risk bCrude rate defined as the number of cases per 100,000 dogs/year cAll dogs insured with a single UK pet insurance company dClaims for veterinary treatment identified as being related to neoplasia
Country Years
Population location
Number of dogs estimated to be
at riska Tumors Incidence rateb
UK 1997-1998 UK insured dogsc 130,684 Cancersd 747.9
All neoplasia-related claimsd 1948
California, USA 1963-1966 Alameda County 1031 Cancers 381
Nonmalignant tumors 1130
Ontario, Canada 1999 Veterinary clinics 63,500 Cancers 850
All tumors 3970
Italy 1985-1994 Genoa County 127,600 Cancers 310
Nonmalignant tumors 760
Along with the similar cancer morbidities, the ‘essence’ of the problem of human cancers is also captured by canine cancer by the characterization of tumor growth over long periods of time in the setting of an intact immune system and inter-individual and intra-tumoral heterogeneity. Similarly, the development of recurrent or resistant disease and metastasis to relevant distant sites such as the lungs and/or liver is also seen in dogs (Khanna et al. 2006). In addition, the manner in which dogs respond to tumor treatment is comparable to humans (Cadieu and Ostrander 2007) thus creating an opportunity to use dogs for timely assessment of new cancer therapies as the course of cancer progression is compressed in dogs (Khanna et al. 2006). Characterization of canine cancers at the genomic level and
Figure 1: Dog cancer incidence rates for (A) females and (B) males estimated from tumor specimens received by the ATR from local veterinarians in Genoa, Italy across the study period (1985-2002) for all cancers and selected cancer sites by calendar period. Vertical bands represent the 95% confidence intervals of the estimated incidence rates. (Adapted from Merlo et al. 2008)
The pathology of osteosarcoma
review of 1462 cases, which concluded that dogs weighing > 40 kg accounted for 29% of all cases whereas only 5% of the reported cases weighed < 15 kg (Withrow and Vail 2007). Other studies provide further support for the claim that large dog breeds such as Scottish Deerhounds, Saint Bernards, Great Danes, Irish Setters, Doberman pinschers, Rottweilers, German Shepherds, and Golden Retrievers, are more predisposed to develop OS (Brodey and Riser 1969; Brodey and Abt 1976; Straw et al. 1991; Ru et al. 1998; McNeill et al. 2007). Canine OS also tends to affect middle-aged to older dogs, with a median age of seven years, though studies have reported a bimodal distribution with a second small peak in young dogs aged between 18 and 24 months (Spodnick et al. 1992; Boston et al. 2006; Mueller et al. 2007; Withrow and Vail 2007). The majority of human OS cases occur in adolescence but there is a well described second peak of incidence in the elderly (seventh and eighth decades) (Mirabello et al. 2009a; Mirabello et al. 2009b). The occurrence of OS in large breeds as well as within pedigrees suggest an hereditary basis for the formation of canine OS (Withrow and Vail 2007). Previous publications reported male dogs to be more affected by the disease than females (up to 1.5:1) (Brodey and Riser 1969; Brodey and Abt 1976; Misdorp and Hart 1979; Spodnick et al. 1992) yet evaluation of 1775 cases of canine OS treated at Colorado State University between 1978 and 2005 found the male to female ratio to be equal (Withrow and Vail 2007).
Most cases of canine OS (75%) originate in the appendicular skeleton (humerus, femur, radius, tibia, and ulna) in the metaphysis of long bones (Liptak et al. 2004; Selvarajah and Kirpensteijn 2010) with the remainder occurring in the axial skeleton (flat bones of the skull, ribs, vertebrae, sternum, and pelvis) (Hammer et al. 1995; Dickerson et al. 2001). The front limbs are affected twice as often as rear limbs, with the distal radius (35%) and
In human patients the disease most often arises in the metaphysis of long bones such as the distal femur, proximal tibia, and proximal humerus (Mirabello et al. 2009a; Mirabello et al. 2009b), which is similar to the physiological locations of canine OS.
In humans as well as dogs OS has an aggressive and invasive nature leading to local skeletal destruction as seen in radiographic evidence of both osteoproductive and osteolytic lesions. It is also highly metastatic, predominantly spreading to the lungs with a lower spread frequency to distant bones, regional lymph nodes (Hillers et al. 2005), and other soft tissues (Peremans et al. 2003; Gorman et al. 2006). Several histological subclassifications exist for OS that are based on the type and amount of matrix and characteristics of cancer cells (i.e. osteoblastic, fibroblastic, telangiectatic, chondroblastic and mixed forms)
(Kirpensteijn et al. 2002a; Tang et al. 2008). Examination of the tumor’s histopathological features is the current diagnostic standard and there can be considerable variation in the histological appearance both between and within individual neoplasms making it important to obtain a histological analysis of the tumor following definitive excision. Several factors, such as alkaline phosphatase (ALP) levels, histological grade, and microvascular density, have also been identified as prognostic indicators in dogs, with the most important factor being the detection of metastases already present at the time of diagnosis (Mueller et al. 2007). Histologically, metastatic lesions usually appear identical to the primary tumor but they frequently exhibit a greater degree of necrosis (Selvarajah and Kirpensteijn 2010). A histopathological grading system has been applied to canine OS and it was concluded that tumors classified as grade III had a significantly poorer prognosis than grade I and II neoplasms (Kirpensteijn et al. 2002a). However, this grading system was not significantly prognostic for tumors from non-appendicular sites.
Although advancements in disease management for dogs have occurred over the years, treatment of OS has seen little advancement. Dogs that receive palliative care alone have a median survival time of 1-3 months, while those receiving surgery (usually
months (Brodey and Abt 1976; Spodnick et al. 1992; Kirpensteijn et al. 2002a; Kirpensteijn et al. 2002b; Withrow and Vail 2007) and patients receiving a combination of amputation and chemotherapy treatment have the best prognosis; 50% 1-year and 20% 2-year survival rates (Mueller et al. 2007; Withrow and Vail 2007). ‘Limb-sparing’ is used only to
selectively remove tumors located in the distal radius, ulna, and tibia (Straw and Withrow 1996; Boston et al. 2007) and even with removal of the primary tumor and chemotherapy treatment 90% still succumb to metastatic disease (Selvarajah and Kirpensteijn 2010). In human OS the introduction of new chemotherapy regimens in the 1980’s, which included treatment both before and after definitive surgical resection, helped improve the five-year survival rate to approximately 70%, but little improvement to this rate has been made in the last decade (Mirabello et al. 2009b)
chemotherapy treatment (Selvarajah et al. 2009), suggesting genetic composition of both patient and tumor play a role in prognostic outcome.
Molecular and genetic dysregulation of OS
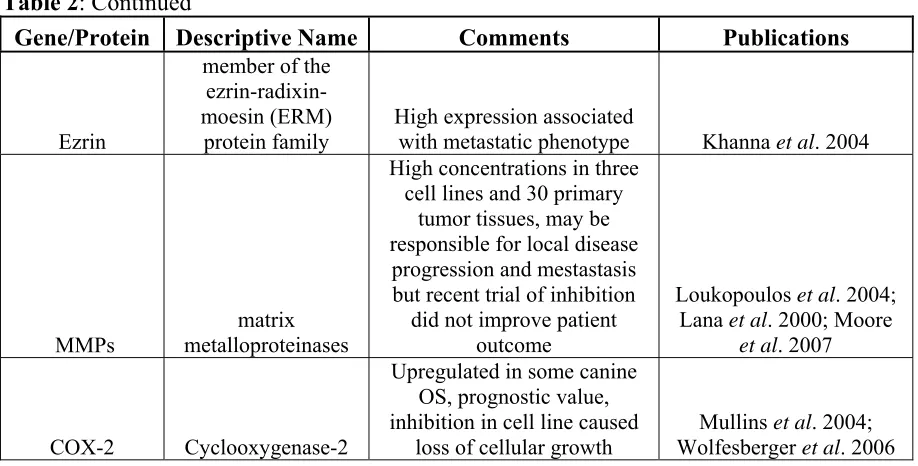
Table 2: Previously defined molecular factors dysregulated in canine OS.
Gene/Protein Descriptive Name Comments Publications
RB1 retinoblastoma 1
Not mutated, but cell lines contained mutations that indirectly inactivated genes
Mendoza et al. 1998; Levine and Fleischli 2000
TP53 tumor protein p53
Mutated and/or overexpressed, protein perform similar biological
activites to those seen in human OS
van Leeuwen et al. 1997; Johnson et al. 1998; Levine and Fleischli 2000; Kirpensteijn et al. 2008; Zhang et al. 2009
PTEN
phosphatase and tensin homolog
Mutated or downregulated in cell lines and tumors, no
prognostic significance
reported to date Levine et al. 2002
erbb-2/HER-2 v-erb-b2 erythroblastic leukemia viral oncogene homolog 2/epidermal growth factor receptor 2
Overexpressed in several canine OS lines and primary
tumors Flint et al. 2004
IGF-1/IGF-1R
insulin-like growth factor-1/insulin-like
growth factor-1 receptor
role in tumor cell invasion and growth, previous clinical
trial supression did not improve outcome
MacEwen et al. 2004; Khanna et al. 2002
MET/HGFR Mesenchymal-epithelial transition/hepatocyte growth factor receptor
Expressed in lung metastases and expression in primary
tumor thought to lead to metastases
Ferracini et al. 2000; Fieten et al. 2009; De
Maria et al. 2009
sis/PDGF
platelet-derived growth factor beta polypeptide (simian sarcoma viral (v-sis) oncogene homolog)
Low level overexpression in canine OS cell lines and
primary tumors
Kochevar et al. 1990; Levine et al. 2002
MYC
v-myc myelocytomatosis
viral oncogene homolog (avian)
Low level overexpression in canine OS cell lines and
primary tumors, recent research saw no significant
changes in expression relative to osteoblasts
Kochevar et al. 1990; Levine et al. 2002
TERT
Telomerase reverse transcriptase gene
Upregulated in canine OS but undergos alternative splicing, need more study
into therapuetic value
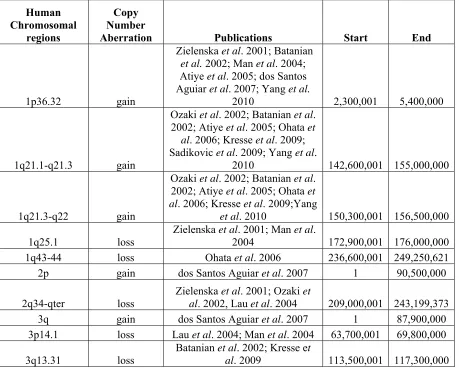
Table 2: Continued
Gene/Protein Descriptive Name Comments Publications
Ezrin
member of the ezrin-radixin-moesin (ERM)
protein family
High expression associated
with metastatic phenotype Khanna et al. 2004
MMPs
matrix metalloproteinases
High concentrations in three cell lines and 30 primary
tumor tissues, may be responsible for local disease progression and mestastasis but recent trial of inhibition
did not improve patient outcome
Loukopoulos et al. 2004; Lana et al. 2000; Moore
et al. 2007
COX-2 Cyclooxygenase-2
Upregulated in some canine OS, prognostic value, inhibition in cell line caused
loss of cellular growth
Mullins et al. 2004; Wolfesberger et al. 2006
oncogenesis and chemotherapy response showed no significant changes in MYC expression in tumors relative to normal osteoblasts suggesting that MYC is not dysregulated at a
transcriptional level in human OS (Sadikovic et al. 2010). Mesenchymal-epithelial transition (MET) is another proto-oncogene and encodes a protein known as c-Met or hepatocyte growth factor receptor (HGFR). MET can activate oncogenic pathways and participate in angiogenesis and metastases in neoplasias. Expression of MET has been found in canine OS lung metastases (Ferracini et al. 2000) and expression of MET in primary tumors was found to predict metatasis via lymphatics (Fieten et al. 2009). MET mRNA is expressed in both human and canine OS cell lines and small molecular inhibitors of MET have been shown in vitro to impair the invasive properties of canine OS cells suggesting the gene as an avenue of study for future treatment options (De Maria et al. 2009).
There are a few genes linked to OS that are not specifically classified as tumor suppressor genes or oncogenes. One such gene, the telomerase reverse transcriptase gene (TERT), is responsible for catalyzing the addition of telomeric sequences onto the 3’-ends of chromosomes de novo and is involved in cell proliferation and tumorigenesis in canine OS. The majority of canine OS patients are telomerase positive suggesting that it may be a
valuable target for canine OS therapy (Kow et al. 2008). A recent study, however, concluded that dog TERT undergoes alternative splicing and the different isoforms may have
implications in gene regulation. Thus, more studies would need to be conducted on the regulation of telomerase activity in canine normal and OS cells in order to properly understand the role of this gene in the disease (Angelopoulou et al. 2008).
In addition to gene based studies of canine OS, several groups have analyzed protein dysregulation in the disease. The matrix metalloproteinases (MMPs) are zinc-dependent enzymes that may be partially responsible for local tumor progression and metastasis. The MMPs have been implicated as biomarkers of shorter disease free interval (DFI) in human OS (Uchibori et al. 2006) as well as a predictor of survival following neo-adjuvant
OS cases had greater MMP expression in tumor than stromal cells, suggesting the proteins may be partially responsible for local disease progression and metastasis (Lana et al. 2000). Yet, a recent study that inhibited MMP-2 expression using BAY 12-9566, a non-peptide biphenyl MMP inhibitor, in combination with doxorubicin chemotherapy in 303 dogs with OS concluded that the inhibition of MMP-2 did not improve survival rates (Moore et al. 2007). The insulin-like growth factor-1 (IGF-1) and its receptor IGF-1R have been shown to play a significant role in OS cell growth and invasion of canine and human OS cell lines (MacEwen et al. 2004). Yet, a previous randomized blind, placebo-controlled preclinical study in dogs with OS concluded that suppression of IGF levels did not sufficiently improve patient outcome (Khanna et al. 2002). Cyclooxygenase (COX-2), an enzyme involved in apoptosis, is upregulated in canine appendicular OS and increased expression was correlated with a significant decrease in survival time for patients relative to tumors that had minimal expression of the enzyme (Mullins et al. 2004). Analysis of the affect of COX-2 inhibition by meloxicam, a non-steroidal anti-inflammatory drug, on the canine OS cell line D-17 found that meloxicam induced a time- and dose-dependent inhibition of cellular growth
(Wolfesberger et al. 2006), yet more studies are needed to conclude its potential for treatment of osteosarcoma in dogs. High expression of another protein, ezrin, a member of the ezrin-radixin-moesin (ERM) family, is associated with early development of metastases in canine OS patients. In pediatric OS, patterns of high expression are linked to poor clinical outcomes (Khanna et al. 2004).
Genomic Studies
made it easier to assess changes in the dog and make direct genomic comparisons between the two.
In human OS, extensive research has identified the cytogenetic and molecular abnormalities, presenting a thorough knowledge basis from which to compare aberrations to canine OS. In contrast to other human sarcomas conventional OS is not associated with a specific recurrent translocation or any other chromosomal rearrangement. In fact, it is characterized by an array of sequential and well-orchestrated genetic changes that involve numerous tumor-suppressor genes and oncogenes (Sandberg and Bridge 2003).
Cytogenetically, human OS shows markedly abnormal karyotypes that contain complex structural changes (translocations and/or rearrangements) and DNA copy number changes (Bayani et al. 2007). Evaluation of human OS karyotypes using a combination of
comparative genomic hybridization (CGH), spectral karyotyping (SKY), and multicolor banding (mBAND) has identified extensive and frequent genome reorganization, including a high degree of aneuploidy, gene amplification, and multiple unbalanced chromosomal rearrangements (Bridge et al. 1997; Zielenska et al. 2001; Ozaki et al. 2002; Bayani et al. 2003; Ozaki et al. 2003; Lim et al. 2004; Man et al. 2004; Lim et al. 2005; dos Santos Aguiar et al. 2007; Selvarajah et al. 2008; Sadikovic et al. 2009). The ploidy number in human OS ranges from haploidy to near-hexaploid and fewer than 5% of the human OS tumors that contain copy number aberrations (CNAs) had only single karyotype changes (Sandberg and Bridge 2003). An example of SKY characterization of the chaotic karyotypes in human OS is shown in figure 2 (Bayani et al. 2003). This study showed that most
karotypes were pseudotriploid for all chromosomes, or had loss or gain of several chromosomes. The overall findings revealed a high frequency of centrometric
rearrangements and recurrent aberrations of human 8q23-q24, 17p11-p13, and 20q (Bayani et al. 2003) (Figure 2A,C).
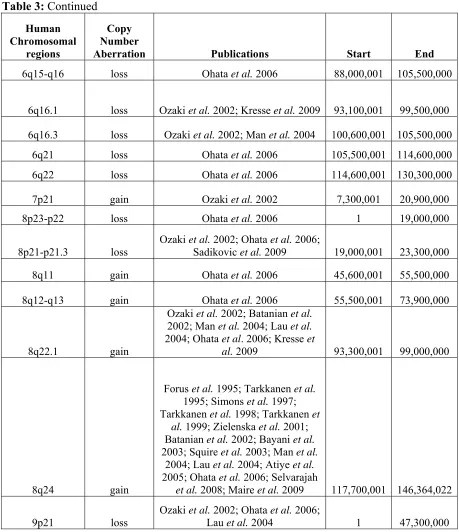
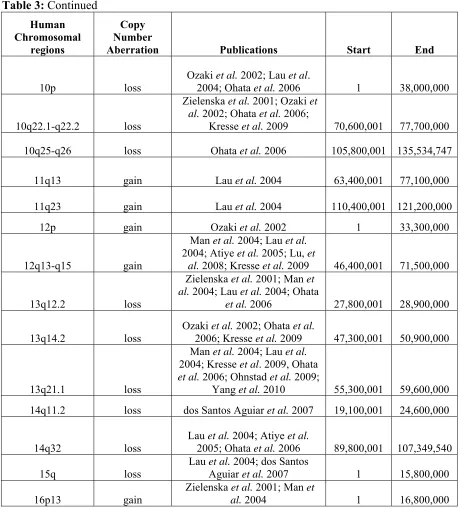
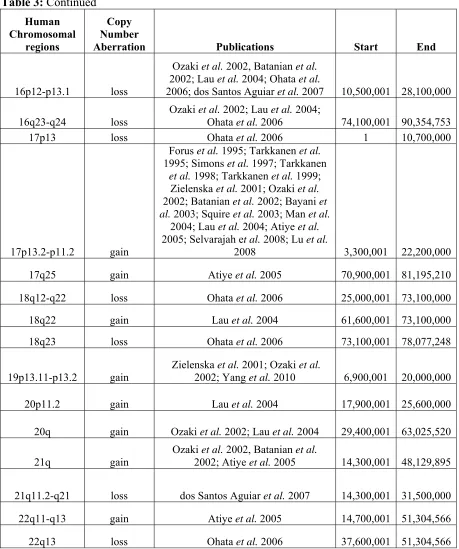
CGH identified numerous regions of copy number changes (Lau et al. 2004; Atiye et al. 2005; Ohata et al. 2006; dos Santos Aguiar et al. 2007), but development of array CGH technology over the last decade has allowed researchers to ‘zoom in’ on CNAs by enhancing the genomic resolution and ability to identify subtle imbalances such as microdeletions (Squire et al. 2003; Selvarajah et al. 2007). Looking across publications seeking to characterize the genomic instability in human OS, overlapping regions of copy number changes could be identified (Table 3). The regions with large overlap are located within human chromosome bands 6p22-p21, 8q24 and 17p13.2-p11.2 and are consistently involved in high copy number gain (log2 ratio of tumor:reference ≥ 1.0) or amplification (log2 ratio of tumor:reference ≥ 2.0) (Forus et al. 1995; Tarkkanen et al. 1995; Simons et al. 1997;
Tarkkanen et al. 1998; Tarkkanen et al. 1999; Zielenska et al. 2001; Batanian et al. 2002; Bayani et al. 2003; Man et al. 2004; dos Santos Aguiar et al. 2007; Selvarajah et al. 2008; Sadikovic et al. 2009; Yang et al. 2010). These large regions of recurrent genomic
Figure 2: SKY analysis of human OS. (A-C) All aberrations, the inverted DAPI and RGB (red-green-blue) images of the fluorescently labeled chromosomes are illustrated. (D) Only SKY-classified colors are illustrated. (A) Aberrations involving chromosome 8 in four primary OS tumors and three cell lines. (B) Evolution of new rearrangements of
der(4)t(4;7)(p16;?p15) in primary tumor OS10. (C) Aberrations involving chromosome 20 in eight primary OS tumors and two cell lines. (D) Complete karyotypes from SKY analysis of primary tumor 0S9 (Bayani et al. 2003).
In the last few years, some investigators have sought to identify the effect of cytogenetic abnormalities in human OS on tumor progression through evaluation of both copy number and expression changes. In turn they have showcased specific genes that they believe to be important in OS because of dysregulation at both the numerical and
transcriptional level. One study used a region specific array for human chromosome 6p12-21 with 108 overlapping bacterial artificial chromosomes (BACs) and P1 artificial clones
blotting and immunohistochemistry indicated that CDC5L, a cell cycle regulator important for G2-M transition, was a likely candidate oncogene for the 6p12-21 amplicon in human OS (Lu et al. 2008). A similar study profiled 36 primary tumors and 20 cell lines with a 1Mb resolution genomic microarray that consisted of 4,549 BAC and PAC clones as well as the minimal tiling-path between 1q12 and the beginning of 1q25 (Kresse et al. 2009). This study revealed that a small region in 3q13.31 (2.1Mb) was frequently deleted (56%). This region was validated by FISH and contained the LSAMP (limbic system-associated membrane protein) gene which has been reported as a candidate tumor suppressor gene in other human cancer types (Kresse et al. 2009). In addition qRT-PCR analysis found that expression of LSAMP was downregulated and that lower expression was associated with poor survival (Kresse et al. 2009).
Over the last three years researchers have begun to use oligonucleotide based aCGH technology to identify and characterize genetic abnormalities in human OS. Using the Human Genome CGH 44K microarray (Agilent Technologies) to analyze ten tumor specimens Maire et al. were able to identify CNAs spanning the entire human genome at a median resolution of 75kb (Maire et al. 2009). They investigated the expression level of RECQL4, a gene that is mutated in patients with Rothmund-Thomson syndrome that develop OS and also maps to the cytoband 8q24 (often rearranged or gained in copy number) (Maire et al. 2009). There was no evidence to indicate that disruption of 8q24 in OS led to the elevated expression of RECQL4, but FISH and aCGH analysis of the tumor DNA showed that expression levels were strongly copy number-dependent (Maire et al. 2009). The same Human Genome CGH 44K array (Agilent Technologies) was used to evaluate the role of the WWOX gene in 10 human OS cases in combination with immunohistochemical staining of 55 formalin-fixed and paraffin embedded tissues (Yang et al. 2010). Loss of WWOX is
Sadikovic et al. were the first to demonstrate combined analysis of multiple levels of dysregulation in human OS by using novel cytogenetic and gene regulation molecular tools (Sadikovic et al. 2009). They used an integrative approach for genome-wide high resolution profiling of genetic, epigenetic, and gene expression to study the combined-effects of genetic and epigenetic mechanisms on cancer-related gene networks (Sadikovic et al. 2009). This approach was then applied to a panel of primary human OS tumors in order to characterize the correlation between global changes in copy number, DNA methylation, and gene expression. They reported strong associations between copy number aberration, over
expression, and hypomethylation leading to disease progression (Sadikovic et al. 2009). To date this integrative approach has not been completed on canine OS, but some studies have looked at specific aspects of whole genome genetic and epigenetic dysregulation. One study using gene expression profiling on a canine specific cDNA microarray identified 51
transcripts to be differently expressed in canine OS with common upregulation of these genes in patients with a short (<6 months) survival time. These over-expressed genes were
Table 3: Summary of the cytogenetic abnormalities associated with human OS. Human Chromosomal regions Copy Number
Aberration Publications Start End
1p36.32 gain
Zielenska et al. 2001; Batanian et al. 2002; Man et al. 2004; Atiye et al. 2005; dos Santos Aguiar et al. 2007; Yang et al.
2010 2,300,001 5,400,000
1q21.1-q21.3 gain
Ozaki et al. 2002; Batanian et al. 2002; Atiye et al. 2005; Ohata et
al. 2006; Kresse et al. 2009; Sadikovic et al. 2009; Yang et al.
2010 142,600,001 155,000,000
1q21.3-q22 gain
Ozaki et al. 2002; Batanian et al. 2002; Atiye et al. 2005; Ohata et al. 2006; Kresse et al. 2009;Yang
et al. 2010 150,300,001 156,500,000 1q25.1 loss
Zielenska et al. 2001; Man et al.
2004 172,900,001 176,000,000
1q43-44 loss Ohata et al. 2006 236,600,001 249,250,621
2p gain dos Santos Aguiar et al. 2007 1 90,500,000
2q34-qter loss
Zielenska et al. 2001; Ozaki et
al. 2002, Lau et al. 2004 209,000,001 243,199,373
3q gain dos Santos Aguiar et al. 2007 1 87,900,000
3p14.1 loss Lau et al. 2004; Man et al. 2004 63,700,001 69,800,000 3q13.31 loss
Batanian et al. 2002; Kresse et
Table 3: Continued Human Chromosomal regions Copy Number
Aberration Publications Start End
4p15.1 loss Man et al. 2004 27,700,001 35,800,000
4p14 gain Ohata et al. 2006 35,800,001 41,200,000
4p12-p13 gain Ohata et al. 2006 41,200,001 48,200,000
4q13-q21 gain
Ozaki et al. 2002; Batanian et al.
2002; Ohata et al. 2006 59,500,001 88,000,000
4q22-q23 gain Ohata et al. 2006 88,000,001 101,100,000
4q24-q26 gain Ohata et al. 2006 101,100,001 120,800,000
5p14-p15.2 gain
Ozaki et al. 2002; Zielenska et al. 2001; Lau et al. 2004; dos Santos
Aguiar et al. 2007 15,000,000 28,900,000
5q11.2 gain
Ohata et al. 2006; dos Santos
Aguiar et al. 2007 50,700,001 58,900,000
5q12.3-q13.2 loss Yang et al. 2010 63,200,001 73,300,000
5q14.3-q22.2 loss Yang et al. 2010 82,800,001 113,100,000
5q32-q33.1 loss Ozaki et al. 2002 144,500,001 152,700,000
5q34 loss Ohata et al. 2006 159,900,001 168,500,000
6p22-p21 gain
Forus et al. 1995; Tarkkanen et al. 1995; Simons et al. 1997; Tarkkanen et al. 1998; Tarkkanen et
al. 1999; Zielenska et al. 2001; Batanian et al. 2002; Bayani et al. 2003; Squire et al. 2003; Man et al.
2004; dos Santos Aguiar et al. 2007; Selvarajah et al. 2008; Yang
et al. 2010 15,200,001 46,200,000
6p21-p12 gain
Ozaki et al. 2002; Man et al. 2004; Lau et al. 2004; dos Santos Aguiar
et al. 2007; Lu et al. 2008 30,400,001 57,000,000 6p12.3-p21.1 gain
Atiye et al. 2005; Sadikovic et al.
2009 40,500,001 51,800,000
Table 3: Continued Human Chromosomal regions Copy Number
Aberration Publications Start End
6q15-q16 loss Ohata et al. 2006 88,000,001 105,500,000
6q16.1 loss Ozaki et al. 2002; Kresse et al. 2009 93,100,001 99,500,000 6q16.3 loss Ozaki et al. 2002; Man et al. 2004 100,600,001 105,500,000
6q21 loss Ohata et al. 2006 105,500,001 114,600,000
6q22 loss Ohata et al. 2006 114,600,001 130,300,000
7p21 gain Ozaki et al. 2002 7,300,001 20,900,000
8p23-p22 loss Ohata et al. 2006 1 19,000,000
8p21-p21.3 loss
Ozaki et al. 2002; Ohata et al. 2006;
Sadikovic et al. 2009 19,000,001 23,300,000
8q11 gain Ohata et al. 2006 45,600,001 55,500,000
8q12-q13 gain Ohata et al. 2006 55,500,001 73,900,000
8q22.1 gain
Ozaki et al. 2002; Batanian et al. 2002; Man et al. 2004; Lau et al. 2004; Ohata et al. 2006; Kresse et
al. 2009 93,300,001 99,000,000
8q24 gain
Forus et al. 1995; Tarkkanen et al. 1995; Simons et al. 1997; Tarkkanen et al. 1998; Tarkkanen et
al. 1999; Zielenska et al. 2001; Batanian et al. 2002; Bayani et al. 2003; Squire et al. 2003; Man et al.
2004; Lau et al. 2004; Atiye et al. 2005; Ohata et al. 2006; Selvarajah
et al. 2008; Maire et al. 2009 117,700,001 146,364,022
9p21 loss
Ozaki et al. 2002; Ohata et al. 2006;
Table 3: Continued Human Chromosomal regions Copy Number
Aberration Publications Start End
10p loss
Ozaki et al. 2002; Lau et al.
2004; Ohata et al. 2006 1 38,000,000
10q22.1-q22.2 loss
Zielenska et al. 2001; Ozaki et al. 2002; Ohata et al. 2006;
Kresse et al. 2009 70,600,001 77,700,000
10q25-q26 loss Ohata et al. 2006 105,800,001 135,534,747
11q13 gain Lau et al. 2004 63,400,001 77,100,000
11q23 gain Lau et al. 2004 110,400,001 121,200,000
12p gain Ozaki et al. 2002 1 33,300,000
12q13-q15 gain
Man et al. 2004; Lau et al. 2004; Atiye et al. 2005; Lu, et
al. 2008; Kresse et al. 2009 46,400,001 71,500,000
13q12.2 loss
Zielenska et al. 2001; Man et al. 2004; Lau et al. 2004; Ohata
et al. 2006 27,800,001 28,900,000
13q14.2 loss
Ozaki et al. 2002; Ohata et al.
2006; Kresse et al. 2009 47,300,001 50,900,000
13q21.1 loss
Man et al. 2004; Lau et al. 2004; Kresse et al. 2009, Ohata et al. 2006; Ohnstad et al. 2009;
Yang et al. 2010 55,300,001 59,600,000
14q11.2 loss dos Santos Aguiar et al. 2007 19,100,001 24,600,000
14q32 loss
Lau et al. 2004; Atiye et al.
2005; Ohata et al. 2006 89,800,001 107,349,540
15q loss
Lau et al. 2004; dos Santos
Aguiar et al. 2007 1 15,800,000
16p13 gain
Zielenska et al. 2001; Man et
Table 3: Continued Human Chromosomal regions Copy Number
Aberration Publications Start End
16p12-p13.1 loss
Ozaki et al. 2002, Batanian et al. 2002; Lau et al. 2004; Ohata et al.
2006; dos Santos Aguiar et al. 2007 10,500,001 28,100,000
16q23-q24 loss
Ozaki et al. 2002; Lau et al. 2004;
Ohata et al. 2006 74,100,001 90,354,753
17p13 loss Ohata et al. 2006 1 10,700,000
17p13.2-p11.2 gain
Forus et al. 1995; Tarkkanen et al. 1995; Simons et al. 1997; Tarkkanen
et al. 1998; Tarkkanen et al. 1999; Zielenska et al. 2001; Ozaki et al. 2002; Batanian et al. 2002; Bayani et al. 2003; Squire et al. 2003; Man et al.
2004; Lau et al. 2004; Atiye et al. 2005; Selvarajah et al. 2008; Lu et al.
2008 3,300,001 22,200,000
17q25 gain Atiye et al. 2005 70,900,001 81,195,210
18q12-q22 loss Ohata et al. 2006 25,000,001 73,100,000
18q22 gain Lau et al. 2004 61,600,001 73,100,000
18q23 loss Ohata et al. 2006 73,100,001 78,077,248
19p13.11-p13.2 gain
Zielenska et al. 2001; Ozaki et al.
2002; Yang et al. 2010 6,900,001 20,000,000
20p11.2 gain Lau et al. 2004 17,900,001 25,600,000
20q gain Ozaki et al. 2002; Lau et al. 2004 29,400,001 63,025,520
21q gain
Ozaki et al. 2002, Batanian et al.
2002; Atiye et al. 2005 14,300,001 48,129,895
21q11.2-q21 loss dos Santos Aguiar et al. 2007 14,300,001 31,500,000
22q11-q13 gain Atiye et al. 2005 14,700,001 51,304,566
While many expression-based studies of canine OS have been completed, analysis of genomic CNAs in canine OS has only recently been done. Low-resolution (10-20Mb) aCGH analysis of 38 cases of canine OS consisting of two breeds (Rottweilers and Golden
Retrievers) identified a high degree of genomic instability that resulted in extensive
cytogenetic disorganization, as is the case in human OS (Thomas et al. 2009). An example of this genomic instability is shown in figure 3, FISH analysis of 10 BAC clones on
metaphase and interphase nuclei from a case of osteoblastic OS in a female Rottweiler (Thomas et al. 2009). The metaphase spreads contained numerous bi-armed metacentric chromosomes which are likely the result of fusion events between acrocentric chromosomes. This figure also shows the extent of the copy number aberrations concluded from the aCGH analysis as MYC has upwards of 11 copies and PTEN is a homozygous deletion.
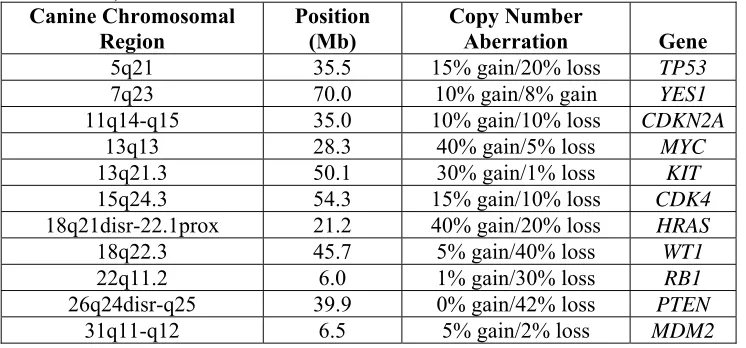
Table 4: Summary of highly recurrent genomic aberrations (≥30%) in 38 patients of canine OS indentified by low-resolution (10-20Mb) aCGH. (Adapted from Thomas et al. 2009)
Canine Chromosomal Region Position (Mb) Copy Number Aberration
1q36 116.7 gain
13q13 28.3 gain
13q14 29.4 gain
13q21.1 36.1 gain 13q21.3 50.1 gain 16q12prox 13.6 loss
16q21mid 30.6 loss 18q21-22.1 21.2 gain 18q22.3 45.7 loss 26q24-q25 28.1 loss
26q25 39.9 loss
29q12 13 loss
31q15.3 37.6 gain 35q14-q15 12.5 loss
Table 5: Summary of copy number imbalances for known cancer-associated genes in 38 patients of canine OS identified by low-resolution (10-20Mb) aCGH. (Adapted from Thomas et al. 2009)
Canine Chromosomal Region Position (Mb) Copy Number Aberration Gene
5q21 35.5 15% gain/20% loss TP53
7q23 70.0 10% gain/8% gain YES1
11q14-q15 35.0 10% gain/10% loss CDKN2A
13q13 28.3 40% gain/5% loss MYC
13q21.3 50.1 30% gain/1% loss KIT
15q24.3 54.3 15% gain/10% loss CDK4
18q21disr-22.1prox 21.2 40% gain/20% loss HRAS
18q22.3 45.7 5% gain/40% loss WT1
22q11.2 6.0 1% gain/30% loss RB1
26q24disr-q25 39.9 0% gain/42% loss PTEN
Figure 3: FISH analysis of dog OS as directed by aCGH. (a) & (b) Multicolor FISH analysis of 10 BAC clones on metaphase (left) and interphase nuclei (right) from a case of
Thesis Outline
In the following chapters of this thesis I enhanced the previous study of genomic imbalance in canine OS (Thomas et al. 2009) by conducting aCGH analysis; at 1Mb-resolution on 123 cases of canine OS and at ~27kb 1Mb-resolution for 23 of the 123 cases. Specific dog breeds, Greyhounds, Golden Retrievers, Rottweilers, and Great Pyrenees, were recruited to identify if any regions of genomic instability were associated with genetic background. Subsequent metaphase and interphase fluorescence in-situ hybridization (FISH) analysis were performed on cells from select canine OS patients to visualize and calculate the extent of copy number gain and loss. My aCGH analysis found that previously defined cancer related genes resided in genomic regions of high aberration frequency on CFA 6, CFA 9, CFA 12, CFA 13, CFA 16, and CFA 26. I then determined the effect of copy number aberration (CNA) on the transcriptional regulation of these genes. Lastly, 15 cases of human OS were profiled at ~100kb resolution to identify orthologous regions of CNAs between dogs and humans. In summary, this thesis (1) characterizes the genomic imbalance in canine OS, (2) concludes that ortholgous regions between humans and canines have similar patterns of CNAs, and (3) identifies cancer-associated genes in regions of high aberration frequency warranting future evaluation in canine and human OS. The parallels that I found in the following chapters between genetic aberrations in canine and human OS help to identify recurrent cross-species abnormalities that could be driving cancer progression and
References
Angelopoulou, K., M. Zavlaris, N. Papaioannou and I. Vlemmas (2008). "Canis familiaris telomerase reverse transcriptase undergoes alternative splicing." Mamm Genome
19(9): 647-53.
Anninga, J. K., M. J. van de Vijver, A. M. Cleton-Jansen, P. M. Kristel, A. H. Taminiau, M. Nooij, R. M. Egeler and P. C. Hogendoorn (2004). "Overexpression of the HER-2 oncogene does not play a role in high-grade osteosarcomas." Eur J Cancer 40(7): 963-70.
Araki, N., A. Uchida, T. Kimura, H. Yoshikawa, Y. Aoki, T. Ueda, S. Takai, T. Miki and K. Ono (1991). "Involvement of the retinoblastoma gene in primary osteosarcomas and other bone and soft-tissue tumors." Clin Orthop Relat Res(270): 271-7.
Atiye, J., M. Wolf, S. Kaur, O. Monni, T. Bohling, A. Kivioja, E. Tas, M. Serra, M. Tarkkanen and S. Knuutila (2005). "Gene amplifications in osteosarcoma-CGH microarray analysis." Genes Chromosomes Cancer 42(2): 158-63.
Bailey, D., H. Erb, L. Williams, D. Ruslander and M. Hauck (2003). "Carboplatin and doxorubicin combination chemotherapy for the treatment of appendicular osteosarcoma in the dog." J Vet Intern Med 17(2): 199-205.
Barabas, K., R. Milner, D. Lurie and C. Adin (2008). "Cisplatin: a review of toxicities and therapeutic applications." Vet Comp Oncol 6(1): 1-18.
Batanian, J. R., L. R. Cavalli, N. M. Aldosari, E. Ma, C. Sotelo-Avila, M. B. Ramos, J. D. Rone, C. M. Thorpe and B. R. Haddad (2002). "Evaluation of paediatric
osteosarcomas by classic cytogenetic and CGH analyses." Mol Pathol 55(6): 389-93.
Bayani, J., S. Selvarajah, G. Maire, B. Vukovic, K. Al-Romaih, M. Zielenska and J. A. Squire (2007). "Genomic mechanisms and measurement of structural and numerical instability in cancer cells." Semin Cancer Biol 17(1): 5-18.
Berg, J., M. C. Gebhardt and W. M. Rand (1997). "Effect of timing of postoperative chemotherapy on survival of dogs with osteosarcoma." Cancer 79(7): 1343-50.
Berg, J., M. J. Weinstein, D. S. Springfield and W. M. Rand (1995). "Results of surgery and doxorubicin chemotherapy in dogs with osteosarcoma." J Am Vet Med Assoc
206(10): 1555-60.
Bergman, P. J., E. G. MacEwen, I. D. Kurzman, C. J. Henry, A. S. Hammer, D. W. Knapp, A. Hale, S. A. Kruth, M. K. Klein, J. Klausner, A. M. Norris, D. McCaw, R. C. Straw and S. J. Withrow (1996). "Amputation and carboplatin for treatment of dogs with osteosarcoma: 48 cases (1991 to 1993)." J Vet Intern Med 10(2): 76-81.
Boston, S. E., F. Duerr, N. Bacon, S. Larue, E. J. Ehrhart and S. Withrow (2007).
"Intraoperative radiation for limb sparing of the distal aspect of the radius without transcarpal plating in five dogs." Vet Surg 36(4): 314-23.
Boston, S. E., N. P. Ehrhart, W. S. Dernell, M. Lafferty and S. J. Withrow (2006). "Evaluation of survival time in dogs with stage III osteosarcoma that undergo treatment: 90 cases (1985-2004)." J Am Vet Med Assoc 228(12): 1905-8.
Boyko, A. R., P. Quignon, L. Li, J. J. Schoenebeck, J. D. Degenhardt, K. E. Lohmueller, K. Zhao, A. Brisbin, H. G. Parker, B. M. Vonholdt, M. Cargill, A. Auton, A. Reynolds, A. G. Elkahloun, M. Castelhano, D. S. Mosher, N. B. Sutter, G. S. Johnson, J. Novembre, M. J. Hubisz, A. Siepel, R. K. Wayne, C. D. Bustamante and E. A.
Ostrander (2010). "A simple genetic architecture underlies morphological variation in dogs." PLoS Biol 8(8).
Breen, M., C. Hitte, T. D. Lorentzen, R. Thomas, E. Cadieu, L. Sabacan, A. Scott, G. Evanno, H. G. Parker, E. F. Kirkness, R. Hudson, R. Guyon, G. G. Mahairas, B. Gelfenbeyn, C. M. Fraser, C. Andre, F. Galibert and E. A. Ostrander (2004). "An integrated 4249 marker FISH/RH map of the canine genome." BMC Genomics 5(1): 65.
radiation-hybrid/linkage map of the domestic dog genome to all chromosomes." Genome Res 11(10): 1784-95.
Breen, M. and J. F. Modiano (2008). "Evolutionarily conserved cytogenetic changes in hematological malignancies of dogs and humans--man and his best friend share more than companionship." Chromosome Res 16(1): 145-54.
Bridge, J. A., M. Nelson, E. McComb, M. H. McGuire, H. Rosenthal, G. Vergara, G. E. Maale, S. Spanier and J. R. Neff (1997). "Cytogenetic findings in 73 osteosarcoma specimens and a review of the literature." Cancer Genet Cytogenet 95(1): 74-87.
Brodey, R. S. and D. A. Abt (1976). "Results of surgical treatment in 65 dogs with osteosarcoma." J Am Vet Med Assoc 168(11): 1032-5.
Brodey, R. S. and W. H. Riser (1969). "Canine osteosarcoma. A clinicopathologic study of 194 cases." Clin Orthop Relat Res 62: 54-64.
Bucan, M. and T. Abel (2002). "The mouse: genetics meets behaviour." Nat Rev Genet 3(2): 114-23.
Cadieu, E. and E. A. Ostrander (2007). "Canine genetics offers new mechanisms for the study of human cancer." Cancer Epidemiol Biomarkers Prev 16(11): 2181-3.
Chun, R., I. D. Kurzman, C. G. Couto, J. Klausner, C. Henry and E. G. MacEwen (2000). "Cisplatin and doxorubicin combination chemotherapy for the treatment of canine osteosarcoma: a pilot study." J Vet Intern Med 14(5): 495-8.
De Maria, R., S. Miretti, S. Iussich, M. Olivero, E. Morello, A. Bertotti, J. G. Christensen, B. Biolatti, R. A. Levine, P. Buracco and M. F. Di Renzo (2009). "met oncogene
activation qualifies spontaneous canine osteosarcoma as a suitable pre-clinical model of human osteosarcoma." J Pathol 218(3): 399-408.
de Nigris, F., C. Botti, R. Rossiello, E. Crimi, V. Sica and C. Napoli (2007). "Cooperation between Myc and YY1 provides novel silencing transcriptional targets of
Dickerson, M. E., R. L. Page, T. A. LaDue, M. L. Hauck, D. E. Thrall, M. E. Stebbins and G. S. Price (2001). "Retrospective analysis of axial skeleton osteosarcoma in 22 large-breed dogs." J Vet Intern Med 15(2): 120-4.
dos Santos Aguiar, S., L. de Jesus Girotto Zambaldi, A. M. dos Santos, W. Pinto, Jr. and S. R. Brandalise (2007). "Comparative genomic hybridization analysis of abnormalities in chromosome 21 in childhood osteosarcoma." Cancer Genet Cytogenet 175(1): 35-40.
Ferracini, R., P. Angelini, E. Cagliero, A. Linari, M. Martano, J. Wunder and P. Buracco (2000). "MET oncogene aberrant expression in canine osteosarcoma." J Orthop Res
18(2): 253-6.
Fieten, H., B. Spee, J. Ijzer, M. J. Kik, L. C. Penning and J. Kirpensteijn (2009). "Expression of hepatocyte growth factor and the proto-oncogenic receptor c-Met in canine
osteosarcoma." Vet Pathol 46(5): 869-77.
Flint, A. F., L. U'Ren, M. E. Legare, S. J. Withrow, W. Dernell and W. H. Hanneman (2004). "Overexpression of the erbB-2 proto-oncogene in canine osteosarcoma cell lines and tumors." Vet Pathol 41(3): 291-6.
Forus, A., D. O. Weghuis, D. Smeets, O. Fodstad, O. Myklebost and A. Geurts van Kessel (1995). "Comparative genomic hybridization analysis of human sarcomas: II. Identification of novel amplicons at 6p and 17p in osteosarcomas." Genes Chromosomes Cancer 14(1): 15-21.
Fossey, S. L., A. T. Liao, J. K. McCleese, M. D. Bear, J. Lin, P. K. Li, W. C. Kisseberth and C. A. London (2009). "Characterization of STAT3 activation and expression in canine and human osteosarcoma." BMC Cancer 9(1): 81.
Foukas, A. F., N. S. Deshmukh, R. J. Grimer, D. C. Mangham, E. G. Mangos and S. Taylor (2002). "Stage-IIB osteosarcomas around the knee. A study of MMP-9 in surviving tumour cells." J Bone Joint Surg Br 84(5): 706-11.
Gershwin, L. J. (2007). "Veterinary autoimmunity: autoimmune diseases in domestic animals." Ann N Y Acad Sci 1109: 109-16.
Gorlick, R., A. G. Huvos, G. Heller, A. Aledo, G. P. Beardsley, J. H. Healey and P. A. Meyers (1999). "Expression of HER2/erbB-2 correlates with survival in osteosarcoma." J Clin Oncol 17(9): 2781-8.
Gorman, E., A. M. Barger, J. M. Wypij and M. E. Pinkerton (2006). "Cutaneous metastasis of primary appendicular osteosarcoma in a dog." Vet Clin Pathol 35(3): 358-61.
Guyon, R., T. D. Lorentzen, C. Hitte, L. Kim, E. Cadieu, H. G. Parker, P. Quignon, J. K. Lowe, C. Renier, B. Gelfenbeyn, F. Vignaux, H. B. DeFrance, S. Gloux, G. G. Mahairas, C. Andre, F. Galibert and E. A. Ostrander (2003). "A 1-Mb resolution radiation hybrid map of the canine genome." Proc Natl Acad Sci U S A 100(9): 5296-301.
Hammer, A. S., F. R. Weeren, S. E. Weisbrode and S. L. Padgett (1995). "Prognostic factors in dogs with osteosarcomas of the flat or irregular bones." J Am Anim Hosp Assoc
31(4): 321-6.
Hillers, K. R., W. S. Dernell, M. H. Lafferty, S. J. Withrow and S. E. Lana (2005). "Incidence and prognostic importance of lymph node metastases in dogs with appendicular osteosarcoma: 228 cases (1986-2003)." J Am Vet Med Assoc 226(8): 1364-7.
Hutt, F. B., C. G. Rickard and R. A. Field (1948). "Sex-linked hemophilia in dogs." J. Heredity 39: 3-9.
Jemal, A., R. Siegel, E. Ward, Y. Hao, J. Xu and M. J. Thun (2009). "Cancer statistics, 2009." CA Cancer J Clin 59(4): 225-49.
Johnson, A. S., C. G. Couto and C. M. Weghorst (1998). "Mutation of the p53 tumor suppressor gene in spontaneously occurring osteosarcomas of the dog." Carcinogenesis 19(1): 213-7.
Jones, P., K. Chase, A. Martin, P. Davern, E. A. Ostrander and K. G. Lark (2008). "Single-nucleotide-polymorphism-based association mapping of dog stereotypes." Genetics
Karlsson, E. K. and K. Lindblad-Toh (2008). "Leader of the pack: gene mapping in dogs and other model organisms." Nat Rev Genet 9(9): 713-25.
Khanna, C., K. Lindblad-Toh, D. Vail, C. London, P. Bergman, L. Barber, M. Breen, B. Kitchell, E. McNeil, J. F. Modiano, S. Niemi, K. E. Comstock, E. Ostrander, S. Westmoreland and S. Withrow (2006). "The dog as a cancer model." Nat Biotechnol
24(9): 1065-6.
Khanna, C., J. Prehn, D. Hayden, R. D. Cassaday, J. Caylor, S. Jacob, S. M. Bose, S. H. Hong, S. M. Hewitt and L. J. Helman (2002). "A randomized controlled trial of octreotide pamoate long-acting release and carboplatin versus carboplatin alone in dogs with naturally occurring osteosarcoma: evaluation of insulin-like growth factor suppression and chemotherapy." Clin Cancer Res 8(7): 2406-12.
Khanna, C., X. Wan, S. Bose, R. Cassaday, O. Olomu, A. Mendoza, C. Yeung, R. Gorlick, S. M. Hewitt and L. J. Helman (2004). "The membrane-cytoskeleton linker ezrin is necessary for osteosarcoma metastasis." Nat Med 10(2): 182-6.
Kirkness, E. F., V. Bafna, A. L. Halpern, S. Levy, K. Remington, D. B. Rusch, A. L.
Delcher, M. Pop, W. Wang, C. M. Fraser and J. C. Venter (2003). "The dog genome: survey sequencing and comparative analysis." Science 301(5641): 1898-903.
Kirpensteijn, J., M. Kik, G. R. Rutteman and E. Teske (2002a). "Prognostic significance of a new histologic grading system for canine osteosarcoma." Vet Pathol 39(2): 240-6.
Kirpensteijn, J., M. Kik, E. Teske and G. R. Rutteman (2008). "TP53 gene mutations in canine osteosarcoma." Vet Surg 37(5): 454-60.
Kirpensteijn, J., E. Teske, M. Kik, T. Klenner and G. R. Rutteman (2002b). "Lobaplatin as an adjuvant chemotherapy to surgery in canine appendicular osteosarcoma: a phase II evaluation." Anticancer Res 22(5): 2765-70.
Knecht, C. D. and W. A. Priester (1978). "Musculoskeletal tumors in dogs." J Am Vet Med Assoc 172(1): 72-4.
Kow, K., D. H. Thamm, J. Terry, K. Grunerud, S. M. Bailey, S. J. Withrow and S. E. Lana (2008). "Impact of telomerase status on canine osteosarcoma patients." J Vet Intern Med 22(6): 1366-72.
Kresse, S. H., H. O. Ohnstad, E. B. Paulsen, B. Bjerkehagen, K. Szuhai, M. Serra, K. L. Schaefer, O. Myklebost and L. A. Meza-Zepeda (2009). "LSAMP, a novel candidate tumor suppressor gene in human osteosarcomas, identified by array comparative genomic hybridization." Genes Chromosomes Cancer 48(8): 679-93.
Kuntz, C. A., W. S. Dernell, B. E. Powers and S. Withrow (1998). "Extraskeletal osteosarcomas in dogs: 14 cases." J Am Anim Hosp Assoc 34(1): 26-30.
Lana, S. E., G. K. Ogilvie, R. A. Hansen, B. E. Powers, W. S. Dernell and S. J. Withrow (2000). "Identification of matrix metalloproteinases in canine neoplastic tissue." Am J Vet Res 61(2): 111-4.