O F
Rst
R. B. ASHMAN2
Purdue University Agricultural Experiment Stalion, Lafayette, Indiana 47906
Received July 22, 1969
LLELES of the
R locus in maize condition the formation of anthocyanin pig-
A
ment in various plant parts and in the aleurone layer of the seed. Alleles pro- moting anthocyanin production in both the plant and seed are designated R', and there is evidence indicating that some of these alleles are structurally tandem duplications with the plant ( P ) and seed ( S ) color components located in separate duplicated segments (STADLER and EMMERLING 1954,1956).Stippled aleurone (
RSt
) conditions colored spots in an otherwise colorless aleurone and green (no anthocyanin) plant color and is of special interest because of its paramutagenic activity. Paramutable R alleles are invariably reduced in aleurone pigmentation level when extracted from heterozygotes withRst
(BRINK et al. 1968).The stippled gene frequently mutates to self-colored aleurone
( R s c ) ,
and MCWHIRTER and BRINK ( 1 962) postulated that such mutations result from the transposition of a n inhibitor ( I R ) from a n aleurone-pigmenting gene, thus per- mitting its full expression. KERMICLE (1970) has shown that there is a class ofRsc
mutants associated with recombination between outside markers, and these were interpreted as resulting from the separation ofZR
from the aleurone-pig- menting gene (Sc) by crossing over.Mutations to near-colorless aleurone, r ( n c ) , in
RrRSt
plants are of several classes, and their isolation and paramutagenic action have been described (ASH-MAN 1965a, 1965b). Many of these mutants were associated with recombination between outside markers, and some also combined characteristics of the two parental alleles, i.e., paramutagenic activity and red plant color. The origin and characteristics of these mutants also suggest that the stippled phenotype is depen- dent on two or more components separable by crossing over. The investigations reported here describe the crossover-associated resynthesis of the
RSt
gene in heterozygous combinations of nonstippled alleles, and thereby provide experi- mental evidence for the compound structure of theRSt
gene.MATERIALS A N D METHODS
R s t resynthesis was tested in heterozygous combinations of r ( n c ) mutants with self-colored aleurone alleles, and in heterozygous combinations and homozygotes of three rg(nc) mutants from three different rg(nc) mutant classes. The origins and distinguishing characteristics of the self-colored and near-colorless aleurone alleles used i n the tests are described helow.
Journal Paper No. 3755 Purdue University Agricultural Experiment Station. 2 Department of Botany and Plant Pathology.
240 R. B. ASHMAN
Self-colored aleurone alleles: Rr has red plant color, gives mottled aleurone when present in one dose ( R r r ) , and is paramutable. It was obtained from Wisconsin and has been used ex- tensively in studies of paramutation.
RsC106, R"113, Rsc132, and R s c (unnumbered) are representatives of a large number of such mutants from Rat isolated in this and other laboratories. As a group they have green plant color, give self-colored aleurone in one dose, and are not paramutable. They vary in paramutagenic activity, and of the above mutants RSC132 is paramutagenic and the others are nonparamuta- genic. The R S C mutants used in these tests were isolated from X ~ t R s t plants, but it is not known if any were associated with crossing over.
Near-colorless aleurone alleLes: rg(nc)l-3 has stable seed color, unstable green plant color, and is paramutagenic. It is one of eleven mutants with thcse characteristics isolated from RrRst
plants marked distally and all carried the marker from the R't chromosome; seven were isolated from plants also marked proximally and all carried the marker from the R T chromosome. Mutants
of this class are clearly of crossover origin.
rg(nc)l-2 has unstable seed color, stab!e green plant color, and is paramutagenic. I t was one of five mutants with these characteristics isolated from RrRSt plants marked distally and four carried the marker from the Rlt chromosome; three were isolated from plants also marked proximally and all carried the marker from the Rst chromosome. Three mutants of similar phenotype with unstable seed color have been isolated from R"RSt plants. These data indicate a noncrossover origin for mutants of this class.
rs(nc)2-7 has stable seed color, stable green plant color, and is paramutagenic. It is one of three mutants with these characteristics isolated from RrRSt plants marked b3th proximally and distally, and two carried the proximal marker from the Rr and all the distal marker from the Rst
chromosome. Mutants of this class are probably of crossover origin. but the limited data are not conclusive.
r r ( n c ) mutants have stable seed color, red plant color. and are paramutagenic. Nineteen mutants with these characteristics isolated from R'RSt plants marked distally all carried the marker from the RSt chromosome; 1% isolated from plants also marked proximally all carried the marker from the Rr chromosome. Mutants of this class are clearly associated with crossing over.
The general procedure in all tests was to pollinate the comhinations of alleles with rr (color- less aleurone, red plant) or ro (colorless aleurone, green plant) and score the ears for stippled kernels. The pollen carried a marker, waxy endosperm (wz), in many of the tests to aid in identifying stippled kernels resulting from pollen contamination. Exclusion of stippled kernels resulting from contaminant pollen was positive even in the absence of a pollen marker. Bonafide stippled mutants were recovered heterozygous with the r allele from the pollen parent, while stippled alleles from contamination were recovered heterozygous with the r ( n c ) allele from the particular R / r ( n c ) or r ( n c ) / r ( n c ) combination being tested. The r ( n c ) and r alleles are dis- tinguishable on the basis of phenotype and paramutagenic action, and some crosses were designed to take advantage of differences in plant color. The kernels from each progeny were shelled, bulked, and weighed, and the weight of 1000 kernels was used to estimate the total kernel number in each. Stippled kernels were grown out for verification of phenotme and pollen marker, if present.
Colden plant ( g ) , 14 units proximal to R, and a modifier of the stippled phenotype ( M " ) , six units distal, served as outside genetic markers in tests where outside markers are indicated.
RESULTS
R 24 1
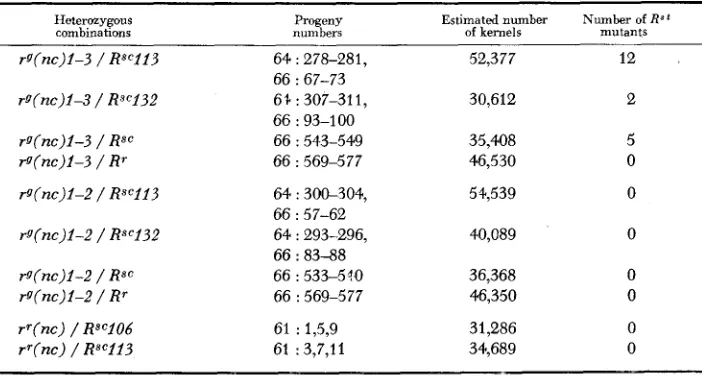
TABLE 1
Frequency of RSt resynthesis in heterozygous combinations of self-colored aleurone alleles with the fallowing near-colorless aleurone mutants: rg (nc) 1-3, stable seed color, unstable
green plant color; rg(nc) 1-2, unstable seed color, stable green plant color; rr(nc), stable seed color, red plant color
Heterozygous
combinations Progeny Estimated
number Number of R s t numbers of kernels mutants
rg(nc)l-3 / RsC113 64 : 278-281, 52,377 12
rg(nc)i-3 / Rscl32 6 % : 307-311, 30,612 2
rg(nc)l-3 / Rsc 66 : 543-549 35,408 5
rg(nc)l-3 / Rr 66 : 569-577 46,530 0
rg(nc)l-Z / RsC113 64 : 306304, 51;539 0
rg(nc)l-2 / Rsc132 64 : 293-296, 40,089 0
rg(nc)l-2 / Rsc 66 : 533-5?0 36,368 0
rg(nc)l-Z / Rr 66 : 569-577 46,350 0
66 : 67-73
66 : 93-1 00
66 : 57-62
66 : 83-88
rr(nc) / RSC106 rr(nc) / RSC113
61 : 1,5,9 61 : 3,7,11
31,286 0
34,689 0
were resynthesized in the combinations of rg(nc)1-3 with each of the three Rsc alleles. Combinations of the same three Rsc alleles with rg(nc)1-2 failed to yield any resynthesized RSt alleles, and none were recovered from combinations of r g ( n c ) l - 3 or rg(nc)1-2 with R'.
The total kernel number from the three Rsc"/g(nc) 1-3 heterozygous combina- tions was 118,397, and 19 resynthesized RSt alleles were recovered-a frequency of 1.6 x No
RSt
mutants were recovered from 130,996 kernels from the three RSc/rg(nc)l-2 heterozygous combinations; based on a frequency of 1.6 Xabout 21 would have been expected, and their absence is clearly significant. The heterozygous combinations involving Rsc113 and Rsc132 carried a distal outside marker, and 11 of the 14 resynthesized
R S t
alleles carried the marker from the rg(nc) 1-3 chromosome. The heterozygous combination involving the un- numbered Rsc allele carried a proximal outside marker, and the five resynthesized RSt alleles all carried the marker from the Rsc chromosome. These data show Rst resynthesis in these heterozygous combinations to be associated with crossing over.242 R. B. ASHMAN
TABLE 2
Frequency of R*t resynthesis in homozygotes and in heterozygous combinations of three rg(nc) alleles representing three different classes of I-p(nc) mutation based on seed and plant
color stability: rg(nc) 1-3, stable seed color, unstable green plant color; rg(nc) 1-2, unstable seed color, stable green plant color;
rg(nc)2-7, stable seed and stable green plant color
Heterozygous
combinations Progeny
Estimated number Number of R a t
numbers of kernels mutants
rg(nc)l-Z / rg(nc)l-3 67 : 315-323 72,207 8 rg(nc)l-Z / rg(nc)Z-7 67 : 285-292 86,506 0 rg(nc)l-3 / rg(nc)2-7 67 : 300-307 80,916 0
rg(nc)l-Z / rg(nc)l-Z rg(nc)l-3 / rg(nc)l-3 rg(nc)2-7 / rg(nc)2-7
62 : 1036 62 : 1037 62 : 1006
32,155 0
22,260 0
29,062 0
ing directly to RSt and were not yielding Rst mutants merely by loss of a com- ponent by crossing over.
Resynthesis of R S t was not related to the paramutagenic activity of the Rsc alleles. The frequency was lower in heterozygous combinations with paramuta- genic R"132 than with nonparamutagenic R"113, but not enough mutants were recovered to establish a significant frequency difference.
The results of tests for RSt resynthesis in heterozygous combinations of r'(nc) mutants with self-colored aleurone alleles are shown in the bottom section of Table 1, and no resynthesized RSt alleles were recovered. Three r'(nc) mutants were tested with each of the two Rsc alleles, and the pooled data are reported in Table 1 .
The results of tests for R S t resynthesis in heterozygous combinations of three classes of rg(nc) mutants are shown ir? the top section of Table 2, and the results from tests of homozygotes are shown in the bottom section. Resynthesized
RSt alleles were recovered from only one of the heterozygous combinations,
rg(nc) I-3/rg(nc) 1-2, and none were recovered from the homozygotes. No outside markers were present in these tests.
The phenotypes of many of the resynthesized R S t alleles appear to be about the same as that of ou r standard R S t allele, but quantitative data for comparisons were not obtained. Two of 1 7 resynthesized RSt alleles have a somewhat lighter phenotype and three a darker, but they do not form clearly discontinuous groups. Four of the resynthesized R S t mutants were tested and found to be paramuta- genic but detailed scorings were not made, and it is not known whether the para- mutation level was equivalent to that of r g ( n c ) l - 3 or our standard R S t allele.
The plant color instability of r g ( 4 1 - 3 is frequently somatic, as evidenced by
R 243
DISCUSSION
The successful resynthesis of Rst alleles by crossing over is evidence that the stippled phenotype is dependent on at least two R locus components. In these tests one component was provided by any of three Rsc mutants and the other by rg(nc)1-3, a near-colorless aleurone stable seed and unstable green plant color mutant isolated from an RTRst plant and associated with crossing over; it carried the distal outside marker from the Rst chromosome and the red plant color com- ponent from RT, although in a suppressed state. Other rg(nc) mutants of the same seed and plant color stability class were isolated from RTRSt plants carrying a n outside proximal marker and all carried the marker from the R' chromosome.
KERMICLE ( 1970), expanding on a model proposed by MCWHIRTER and
BRINK
( 1962), used ScZR to denote the R5 complex with (.) signifying the lack of func- tional independence of Sc, the aleurone-pigmenting component, fromZR,
a n inhibitor. Outside markers showedZR
to be distal to Sc. If the resynthesis of Rst is viewed in terms of this model, Sc was provided by the R8" alleles andZR
byr g ( n c) 1-3. Resynthesized RSt alleles recovered from heterozygous combinations
with outside markers carried the proximal marker from the R"" chromosome and the distal marker from the rg(nc)1-3 chromosome, facts consistent with the order in the above model. The unstable plant color characteristic of rg(nc)1-3 could be based in the association of
ZR
with a colored plant component,P.
A second class of near-colorless aleurone mutant, represented in these tests by
I"( nc) 1-2, did not yield resynthesized Rst alleles in heterozygotes with R"". rg(nc)1-2 has unstable seed and stable green plant color and carried the distal outside marker from the RSt chromosome when isolated from a n R7RSt plant. No
proximal outside marker was present in the plant from which r g ( n c ) l - 2 was isolated, but three other rg(nc) mutants of the same seed and plant color stability class were isolated from R R S t plants marked proximally and all carried the marker from the RSt chromosome. It is likely, therefore, that mutant rg(nc)1-2 was not associated with crossing over, but the evidence is circumstantial.
rg ( n c ) 1-2 mutates to self-colored aleurone and must carry a suppressed SC, but
the absence of resynthesized RSt alleles in Rsc//r"( nc) 1-2 heterozygotes indicates that it does not carry ZR. These facts, together with the probable noncrossover origin of rg(nc)1-2, suggest that
ZR
mutates to a form that inhibits all Sc expres- sion in endosperm tissue but does not inhibit germinal changes to full Sc expression. The apparent alteration of I n could be from mutation or from a change in its positional relationship with Sc; the data do not distinguish between these two possibilities.The nonstippled (self-colored) phenotype of RSc/rg( n c ) 1-3 endosperms is noteworthy since it implies a location specificity for the interaction of Sc and
ZR
to give a stippled phenotype. Both components are present in such a heterozygote but the stippled phenotype is not produced until they are brought together in thecis configuration by a crossover. r g ( nc) 1 - 2 / ~ 9 ( n c ) 1-3 endosperms also possess
244 R. B. ASHMAN
instance is likely due to the suppression of Sc in the endosperm by an altered form of
ZR.
RSt resynthesis in this heterozygote results from the replacement of an altered with an unalteredZR.
Other indications of possible position effects are suggested by the phenotypes of some resynthesized RSt alleles. Although quantitative measurements were not made, the phenotypic deviations of five resynthesized RSt alleles from our stan- dard RSt allele are sufficient to suggest that slight changes in the positional rela- tionships of RSt components may alter the stippled phenotype, or that the com- por?ents may undergo functional alterations in the process of separation and resynthesis.
Mutant rg(nc)2-7, stable seed and green plant color, was not tested for Rst resynthesis in heterozygotes with R". It did not yield resynthesized RSt alleles when tested in heterozygotes with either rg ( nc) 1-2 or r g ( nc) 1-3, and may carry
neither Sc nor
ZR.
Only a few mutants of this class have been isolated, and while the limited data indicate an association with crossing over, they are not conclu- sive. rg(nc)2-7 and two other mutants of this class (numbered 2-1 and 2-11) are known to be as fully paramutagenic as RSt (ASHMAN 1965b), and if they lack both Sc andZR
these Rst components are not paramutagenically active. VAN DERWALT and BRINK (1969) reported an RSt allele from Bolivia that was not para- mutagenic and concluded that the basis of paramutagenicity is distinct from the components associated with the stippled phecotype.
The r " ( n c ) mutants in heterozygotes with Rsc failed to give resynthesized RSt alleles, and therefore lack the RSt component carried by rg(nc)1-3,
ZR
in the above model. Mutations to r r ( n c ) and to rg ( n c ) 1-3 type mutants in RrRSt plants were associated with recombination between outside markers, and both carried the same combination of markers. Evidently, two crossover sites on the paired homologues are involved in the genesis of these two classes of mutants, or, ifduplicate segments make more than one pairing configuration possible, the two crossovers could involve just one site on one homologue but two on the other. The crossover sites involved have special significance relative to the chromosomal basis of paramutation since r g ( nc) 1-3 type mutants are fully paramutagenic
and rr ( n c ) mutants have reduced activity ( ASHMAN 1965b). These facts, together with others presented above, indicate that
ZR
is distinct from, but closely linked to, the chromosomal site of paramutagenic activity.The evidence for absence of Sc and
ZR
in rg(nc)2-7 type and r r ( n c ) mutants indicates that these R locus components are not the basis of the near-colorless aleurone phenotype. Another aleurone pigmenting component must be present in the R locus complex, and in no class of mutants isolated from RTRst plants was this component separated from paramutagenic activity ( ASHMAN 196513). The distribution of outside markers places the near-colorless aleurone component and the chromosomal site of paramutagenic activity distal toZR.
SUMMARY
R LOCUS C O M P O N E N T S
245
colorless, r ( n c )
,
aleurone. Both mutant groups are heterogeneous, and independ- ent inceptions may vary in association with recombination, level of paramuta- genic action, and seed and plant color stability. These facts suggest that the stippled phenotype is dependent on R locus components that may be separable by crossing over. Heterozygous combinations involving several r ( n c ) mutants and self-colored aleurone alleles were tested for resynthesis of the Rst allele. Resynthesis was successful in heterozygous combinations of Rsc alleles, either paramutagenic or nonparamutagenic, with a near-colorless aleurone mutant, rg(nc)l-3, isolated from a nRrRSt
plant and associated with recombination. Nine- teen Rst mutants were recovered from 118,397 kernels, and outside markers showed resynthesis to be associated with recombination. Heterozygous combina- tions ofRsc
with a noncrossover rg ( nc) mutant, and with rr ( n c ) mutants did not yield resynthesized RSt alleles. Resynthesis of Rst also was successful in one heterozygous combination of different r g ( n c ) mutants. The data on R S t resynthe- sis indicate that the stippled phenotype is dependent on at least two R locus components separable by crossing over, that one component is the structural gene for self-colored aleurone ( S c ) , and the second component is an inhibitor(IR)
ofSc. Neither component is the basis of the near-colorless aleurone phenotype nor paramutagenic activity. Interaction between the components is not interchromo- somal, and the stippled phenotype requires the cis configuration.
LITERATURE CITED
ASHMAN, R. B., 1965a Mutants from maize plants heterozygous RlHsl and their association with crossing over. Genetics 51: 305-312. -, 1965b Paramutagenic action of mutants from maize plants heterozygous RrRSt. Genetics 52: 835-841.
Paramutation: directed genetic change. Science 159: 161-170.
Somatic and meiotic instability of R-stippled, an aleurone spotting factor in maize. Genetics 6p: 247-258.
Mutation of a self-colored allele to a Navajo pattern. Maize Genet. Coop. News Letter 35: 142.
Continuous variation in the level of paramutation at the R locus in maize. Genetics 47: 1053-1074.
Problems of gene structure 111. Relationship of un- equal crossing over to the interdependence of Rr elements (S) and (P). Science 119: 585.
-
, Relation of unequal crossing over to the interdependence of R r elements(P) and (S). Genetics 41 : 124-137.
Geographic distribution of paramutable and paramutagenic R alleles in maize. Genetics 61 : 677-695.
BRINK, R. A., E. D. STYLES and J. D AXTPLL, 1968
KERMICLE, J. L., 1970
MCWHIRTER, K. S., 1961
MCWHIRTER, K. S. and R. A. BRINK, 1962
STADLER, L. J. and M. EMMERLKNG, 1954
1956