EXPERIMENTAL EVOLUTION
O F ANEW ENZYMATIC FUNCTION.
11.
EVOLUTION
OFMULTIPLE FUNCTIONS
FOR
EBGENZYME IN
E. COLZBARRY G. HALL
Microbiology Section, Biologicel Sciences Group, University of Connecticut, Storrs, Connecticut 06268
Manuscript received November 13, 1977 Revised copy received February 10, 1978
ABSTRACT
The evolution of ebgo enzyme of Escherichia coli, an enzyme which is unable to hydrolyze lactose, lactulose, lactobionate, or galactose-arabinoside effectively, has been directed in successive steps so that the evolved enzyme is able to hydrolyze these galactosides effectively. I show that in order for a strain
of E. coli with a lac2 deletion to evolve the ability to use lactobionate as a carbon source, a series of mutations must occur in the ebg genes, and that these mutations must be selected in a particular order. The ordered series of muta- tions constitutes a n obligatory evolutionary pathway for the acquisition of a new function for ebgo enzyme. A comparison of newly evolved strains with parental strains shows that when ebg enzyme acquires a new function, its old functions often suffer; but that in several cases old functions are either unaf- fected or are improved. I conclude that divergence of functions catalyzed by a n enzyme need not require gene duplication.
ACOB
(1977) discussed the importance of “historical accident” in evolutionJ
and made the point that “the appearance of new molecular structures during. . .
evolution must. .
.
have rested on alteration of pre-existing ones.” He further pointed out that “selective pressure resulting from changes in eco- logical niches led to biochemical changes in molecular types.”The ebg (evolved &galactosidase) system of E. coli has been employed as a model system for studying evolution from exactly the point of view put forth by JACOB. In this study the role of historical accident is examined by asking how an experimentally evolved function depends upon the “accident” of choice of the ancestral gene.
A common assumption is that evolution acts to optimize all the function of an organism for its particular ecological niche. This leads to a further assumption that any mutations that alter a pre-existing enzyme so profoundly that it can catalyze a new reaction will simultaneously reduce the effectiveness with which the enzyme carries out its “old” function. According to this hypothesis, evolu- tionary gain of a new function would always be accompanied by loss (or reduced effectiveness) of an old function. This hypothesis is experimentally examined
454 B. G. H A L L
by evolving new functions for ebg enzyme, then determining if old functions of that enzyme have been affected.
ebgA is a structural gene whose wild-type allele, ebgAo, specifies an enzyme with activity toward the synthetic substrate QNPG*, but with very little activity toward the natural sugar lactose
(HARTL
andHALL
1974;HALL
andHARTL
1975; HALL 1976b). Mutations in the ebgA gene can lead to the synthesis of
ebg enzyme with greatly enhanced activity toward lactose (CAMPBELL, LENGYEL
and
LANGRIDGE
1973;HALL
andHARTL
1974), and a number of different alleles specifying ebg enzymes with varying degrees of activity on lactose have been described(HALL
1976;HALL
and CLARKE 1977). Additionally, it has been pos- sible to evolve the ebgA gene toward an alternative destination so that it specifies a n enzyme with methyl-galactosidase activity, but with no enhanced lactase activity(HALL
1976).MATERIALS A N D M E T H O D S
Genetic nomenclature: ebgA is the structural gene specifying ebg enzyme. ebgR is the regula- tory gene specifying the ebg repressor (HALL and HARTL 1975). The wild-type allele, ebgAo,
is present i n wild-type E . coli K12, but its product, ebgo enzyme, can be detected only i n strains completely lacking the classical lac2 p-galactosidase activity.
Strains and growih conditions: E . coli K12 strains are described in Table 1. Minimal medium was as described by HALL and HARTL (1974), supplemented with 10 pg/ml of vitamin B1, and 2
x
10” M IPTG to induce synthesis of the lac permease. Macconkey-lactose medium was prepared according to the instructions supplied by Difco. Solid medium contains 1.5% Noble Special Agar (Difco). Cultures were maintained at 370. Liquid cultures were vigorously aerated by shaking.Measurements of growth rates: Cultures were grown overnight in minimal medium con- taining 2 x 1WM IPTG and sufficient sodium succinate (as a carbon source) to limit cell density to 1 O*/ml. Cultures were washed twice, resuspended i n minimal medium containing IPTG, and carbon sources added to the following concenirations: lactose, 0.1%; lactulose, 0.1%; galactose-arabinoside, 0.1 % and sodium lactobionate, 0.5%. Cultures were shaken a t 370, and aliquots periodically removed and the Asoo n m was determined. Growth rates are reported as a, the first order growth rate constant, in units of hours-1. a is thus the slope of the least-squares fit of ln(A& us. time (hours). Values are reported +. the 95% confidence interval, based upon a minimum of four independent growth curves.
Enzyme assays: ebg enzymes were purified as previously described (HALL 1976), and were assayed for activity toward lactose and lactulose as described by HALL 1976), and for activity toward ONPG and ONPF as described by HALL and HARTL 1975). The unit of activity for all assays is one nanomole of substrate hydrolyzed per minute. Protein concentration were deter- mined according to HALL and HARTL (1975).
Maiings and transductions: Matings were carried out according to MILLER (1972). S t r e p tomycin (100 micrograms/ml) was used as a counterselective agent. Transductions, mediated by phage P1 vir, were carried out according to MILLER (1972). The recipient strain in all matings and transductions was SJ-7. The donor allele tolC+ was selected by plating (or repli- cate plating) to Macconkey lactose medium which prevents the growth of tolC- cells. When employed, minimal medium contained 100 micrograms/ml of arginine and of methionine.
ENZYME EVOLUTION IN
E. coli
455TABLE 1
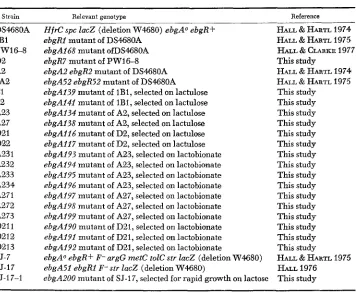
Bacterial strains, E. coli K12
Strain Relevant genotype
DS4680A 1B1 PW16-8 D2 A2 5A2
c1
c2 A23 A27 D21 D22 A23 1 A232 A233 A234 A271 A272 A273 D211 D212 D213 SJ-7 SJ-17 SJ-17-1HfrC spc lac2 (deletion W4680) ebgAO ebgRf ebgRl mutant o f DS468OA
ebgAl68 mutant ofDS468OA ebgR7 mutant o f PW16-8 ebgA2 ebgR2 mutant of DS4680A ebgA52 ebgR52 mutant o f DS4680A ebgAl39 mutant of IBI, selected on lactulose
ebgA141 mutant 08 IBI, selected on lactulose
ebgA134 mutant of A2, selected on lactulose
ebgAl38 mutant of A2, selected on lactulose
ebgAi16 mutant of D2, selected on lactulose
ebgA117 mutant of D2, selected on lactulose
ebgA193 mutant of A23, selected on lactobionate
ebgA194 mutant of A23, selected m lactobionate
ebgA195 mutant of A23, selected on lactobionate
ebgA196 mutant o f A23, selected on lactobionate
ebgA197 mutant of A27, selected on lactobionate
ebgA198 mutant of A27, selected on lactobionate
ebgA199 mutant of A27, selected on lactobionate
ebgA190 mutant of D21, selected on lactobionate
ebgA191 mutant of D21, selected on lactobionate
ebgA192 mutant of D21, selected on lactobionate
ebgAo ebgR+ P- argG metC tolC str lacZ (deletion W46SO)
ebgA51 ebgRl F- str lacZ (deletion W4680)
ebgA200 mutant o f SJ-17, selected f o r rapid growth on lactose
Reference
HALL & HARTL 1974 HALL & HARTL 1975 HALL & CLARKE 1977
HALL & HARTL 1974 HALL & HARTL 1975 This study
This study This study This study This study This study This study This study This study This s b d y This study This study This study This study This study This study
HALL & HARTL 1975 HALL 1976
This study This
RESULTS A N D DISCUSSION
The overall objective of this study was to follow the evolution of a series of new functions for ebg enzyme in order to determine: (1) the effect on an old function when a new function is evolved, and (2) the extent to which the start- ing allele limits the functions that can be evolved for a particular enzyme. I t was necessary to begin by determining the spectrum of functions that could be carried out by the unevolved enzyme ( e b g )
,
and by various forms of the evolved enzyme( e b g + ) , all of which had been evolved to hydrolyze lactose.
Characteristics of lac2 deletion strains selected for growth on lactose
Previous communications have described the properties of ebgo enzyme (wild type) and the enzymes e b g f a , ebg+b. ebgcc, and ebg+d, which were evolved for lactose utilization (HALL 1976; HALL and CLARK 1977).
Table 2 shows the growth rates of lac2 deletion strains of E. coli, which synthesize these enzymes constitutively. Strain 1 B1, which synthesizes the unevolved enzyme, fails to grow on any of the carbon sources listed. Strains A2
45 6 B. G . HALL
TABLE 2
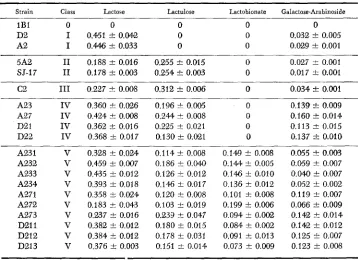
Patterns of growth of individual EBG strains on four galactosides
Strain Class Lactose Lactulose Lactobionate Galactose-Arabinoside
1B1 0 0 0 0 0
D2 I 0.451 f 0.042 0 0 0.032 t 0.005
A2 I 0.446 f 0.033 0 0 0.029 t 0.001
5A2 I1 0.188 f 0.016 0.255 f 0.015 0 0.027 & 0.001 SJ-17 I1 0.178 f 0.003 0.254 t 0.003 0 0.017 t 0.001
c 2 I11 0.227 i 0.008 0.312 t 0.006 0 0.034 zk 0.001
A23 A27 D21 D22 A231 A232 A233 A234 A271 A272 A273 D211 D212 D213 IV IV IV IV V V V V V V V V V V
0.360 i 0.026 0.424 f 0.008
0.362 k 0.016 0.368 t 0.017
0.196 t 0.005
0.244 t 0.088 0.225 t 0.021 0.130 f 0.021
0.328 f 0.0% 0.459 k 0.W7 0.435 t 0.012 0.393 t 0.0'18 0.358 t 0.024 0.183 f 0.043 0.237 f 0.016 0.382 t 0.012 0.384 f 0.012 0.376 k 0.003
0.114 t 0.008 0.186 t 0.040
0.126 t 0.012 0.146 t 0.017 0.120 i- 0.008 0.103 t 0.019 0.233 i- 0.047 0.180 t 0.015 0.178 f 0.031 0.151 t 0.014
0
a
0
0
0.149 t 0.0aS
0.144 f 0.005 0.146 t 0.010 0.136 f 0.012 0.101 f 0.008 0.199 t 0.006 0.094 t 0.002 0.084 t 0.002 0.W1 f 0.013 0.073 f 0.009
0.139 zk 0.009 0.160 t 0.014 0.113 f 0.015 0.137 I O . 0 1 0
0.055 2 0.003 0.059 f 0.007 0.040 t 0.007 0.052 f 0.002 0.119 t 0.007 0.066 t 0.009 0.142 k 0.014 0.142 t 0.012 0.125 f 0.007 0.123 t 0.008
Growth rate (#a +- 95% confidence interval).
(a
>
0.4 h r ' ) on lactose (4-O-/3-D-galactopyranosyl-D-glucose), but are unableto utilize the related disaccharide lactulose (4-O-p-D-galactopyranosyl-D- fructose) as a carbon source. In contrast, strains 5A2 and SJ-17, which synthe- size enzymes ebg+b and ebg+', respectively, grow slowly ((Y
<
0.2 h r ' ) on lac-tose, and grow at a moderate rate (a
>
0.2 hr-') on lactulose.In uitro assays of the purified enzymes from these strains confirm that the growth properties are the result of different ebg enzyme activities (Table 3 ) .
TABLE 3
Activities of EBG enzymes on various substrates
Activities of purified enzyme, units/mg Growth rates 01 hr-' 20mM 20mM 5mM 5 m ~ Strain Enzyme' Lactose Lactuluse Lactose Lactolose ONPG ONPF
A2 e b g f a 0.446 0 I010 0 5490 0
D2 ebg+d 0.451 0 1530 0 7150 0
5A2 ebg+b 0.188 0.255 270 460 9330 404
SJ-17 ebg+c 0.178 0.254 390 620 11,400 620
~
ENZYME EVOLUTION IN
E. co2i
457
Enzymes ebg+b and ebg+c are more active on lactulose than on lactose, while enzyme ebg+a and e b g f d are very active on lactose, but inactive on lactulose. Measurements of the activity of these enzymes on two synthetic substrates, ONPG and ONPF, reveal the same relationships: enzymes and ebg+c are active on both substrates, while e&+“ and ebg+d are restricted to ONPG.
Based upon these observations, two classes of ebg+ strains selected for lactose utilization can be defined: Class I strains are characterized by rapid growth on lactose and failure to grow on lactulose. The ebg enzymes of Class I strains tend to be fairly specific for ONPG and lactose and are more active toward lactose than ebg enzymes from Class I1 strains. Class I1 strains are characterized by slow growth on lactose and somewhat faster growth on lactulose. The ebg enzymes from Class I1 strains have a broader range of substrate specificities than enzymes from Class
I
strains, being active toward lactose, lactulose, ONPG and ONPF.The described strains are the most thoroughly characterized, but they are not the sole representatives of their classes. Thirty-six strains that have evolved from the wild-type organism with the ebgAo allele by selection for lactose utilization, and that are also ebgR- (constitutive), have been classified. Of these 31, or 86% were Class
I
strains, based upon growth rates at>
0.4 hr-l on lactose and failure to grow on lactulose. The remainder were ClassI1
strains, based upon growth rates a: between 0.17 h r ’ and 0.20 hr-l on lactose, and between 0.22 and 0.26 hr-l on lactulose. Both classes arise as the result of single mutations within theebgAo gene
(HALL
1977).W e may also consider the ability of these evolved strains to utilize another related disaccharide, galactose-arabinoside. The wild-type (unevolved) enzyme does not permit galactose-arabinoside to be utilized as a sole carbon source. The evolved strains, selected for lactose utilization, have all acquired a minimal ability to use galactose-arabinoside. There is no difference between Class
I
and Class I1 strains with respect to galactose-arabinoside utilization. The growth rates range from a = 0.017 h r l to cr = 0.032 hr-l, or doubling times of 21 to 40 hours. The observation that Class I and Class I1 strains exhibit very similar growth rates on galactose-arabinoside argues against the hypothesis that ebgenzymes from Class
I1
strains are simply very “sloppy” and will accept virtually any galactose-p-disaccharide. In these experiments lactose utilization was the primary, or selected, function. Class I1 strains also acquired a secondary (non- selected) function, lactulose utilization.Direct selection for lactulose utilization
What happens when the primary and secondary functions are reversed, i.e.,
when lactulose utilization is the primary (selected) function?
W e selected 16 independent lactulose utilizing derivatives of strain 1B1
458 B. G . HALL
0 .
0 .
4 4
4
4
4
-
-
0.1 0.2 0.3 0.4 0.5
d hr-'
Lactose
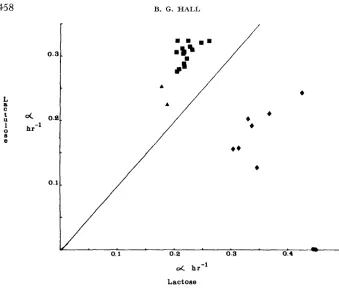
FIGURE 1.-Growth rate on lactulose us. growth rate on lactose. Circles: Class I strains; triangles: Class I1 strains; squares: Class I11 strains; diamonds: Class IV strains. Strains which grow faster on lactulose than on lactose fall above the diagonal line; strains which grow faster on lactose than on lactulose fall below the diagonal line.
together, the mean growth rate on lactose is a = 0.223 f 0.008 hr-l, and a = 0.305 f 0.009 h r ' on lactulose. Like Class
I1
strains, these grow faster on lactulose than on lactose. Based upon significantly faster growth rates on each carbon source, these strains were designated Class 111. All 33 such strains (some obtained in other experiments) isolated by direct selection for lactulose utiliza- tion were able to utilize lactose as a carbon source as well. This observation is in sharp contrast to the observation that direct selection for lactose utilization results in only 14% of the isolates being able to utilize lactulose. One of these strains, for which growth on lactulose was the primary function selected, C2, was chosen for further study. Table 2 shows the growth rates of C2.ENZYME EVOLUTION IN
E. coli
459In what way does the efficiency of a function depend upon the preexisting function, i.e., upon the starting allele? Class I strains grow well upon lactose, but they cannot utilize lactulose.
Employing the same selection method, I obtained six lactulose-utilizing deriva- tives of strain A2 and two lactulose utilizing derivatives of strain D2 (Class
1
strains). The growth rates of these strains on lactose and on lactulose are shown in Figure
1
(diamonds). It is immediately apparent that all of these isolates fall into a new class characterized by growth on each carbon source, but with the rate on lactose exceeding the rate on lactulose. Five derivatives of strainA2
had similar growth rates, a = 0.328 +. 0.025 h r l on lactose, and a = 0.177*
0.026 h r l on lactulose. Strain A23 (Table 2) is representative of this group. The sixth isolate, A27, grew significantly faster on both carbon sources, 01 = 0.424 hr-l onlactose and 01 = 0.244 h r l on lactulose. D21 and D22 were derived from strain
D2; their properties are given in Table 2.
These strains make up Class IV. They are doubly mutant in the ebgA wild- type gene. They are characterized by (1) faster growth on lactose than on lactulose, and (2) three- to four- fold faster growth on galactose-arabinoside than the parental Class I strains. Note that with the exception of strain A27, acquisition of the ability to utilize lactulose has resulted in a decrease in the growth rate on lactose as compared to the parental strains. By contrast, the sec- ondary function, galactose-arabinoside utilization, was vastly improved in these double mutants. The Class
IV
phenotype has never been obtained as a one step mutation of ebgA".Selection for the utilization of sodium lactobionate as a carbon source
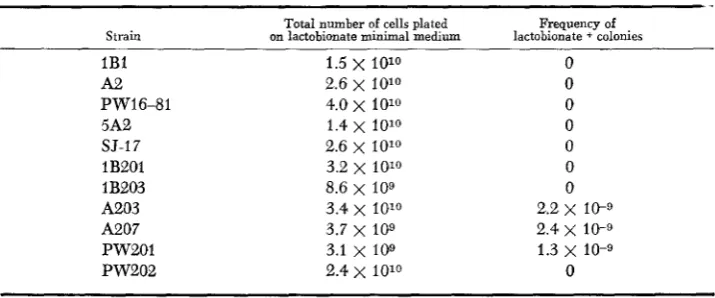
The strains listed in Table 2, Classes 0 through IV, were tested for the ability to utilize sodium lactobionate as a carbon source. As indicated, none were able to use lactobionate. Attempts to isolate lactobionate-utilizing derivatives of those strains by the method used to select lactose and lactulose utilization were unsuc- cessful. Direct selection by plating cells on minimal medium containing lac- tobionate as the sole carbon source succeeded in producing lactobionate-ultilizing derivatives of three Class IV strains at the frequencies indicated in Table 4. The failure to obtain colonies by plating the Class 0 through
I11
strains led to another attempt in which more cells were plated. Again no lactobionate-utilizing deriva- tives were obtained from Classes 0 through 111. If the mutation frequency for lactobionate utilization were 1 O-g in these strains, the probability of failure to detect any lactobionate-utilizing clones is less than 0.001 for each strain. This leads to the conclusion that these lactobionate-utilizing strains, Class V, are triply mutant in the ebgAo wild-type gene.Mapping
460 B. G . HALL
TABLE 4
Frequency of mutation to lactobionate utilization
Total number of cells plated Frequency of Strain on lactobionate minimal medium lactobionate + colonies
1B1 1.5
x
1010 0A2 2.6 x 1010 0
PW16-81 4.0
x 1010
05A2 1.4 x 1010 0
SJ-17 2.6 x 1010 0
1B2Ol 3.2 x 1010 0
1 B203 8.6 x IO9 0
A203 3.4 x 1010 2.2 x 1c-9
A207 3.7 x 109 2.4 x 10-9
PW201 3.1 x 109 1.3 x 10-9
PW202 2.4
x
1010 0phage P1, but are HfrC. These were mated with strain SJ-7 (F- toLC- ebgA’),
and tolC+ recombinants were selected. In each case, the gene for lactose fermen- tation was linked to tolC, with the frequency of recombination between the two ranging from 0.16 to 0.33. The published value for recombination between tolC
and ebgA is 0.29 (HALL and HARTL 1975). From each cross a lactose fermenting
tolC+ recombinant was purified, and it was verified that the recombinant had the
Hfr
parental phenotype with respect to the carbon sources that it could utilize. Phage P I vir was grown on each of the chosen recombinants and used to transduce strain SJ-7. Lactose fermenting transductants were selected on lactose minimal medium and replicated to Macconkey-lactose medium to score for co-transduction of the toZC+ allele. The co-transduction frequencies ranged from 9 % to 31%, in good agreement with previously published values of 11% to 43% co-transduction between ebgA and tolC. Again, a lactose fermentingtolC+ transductant was purified from each transduction experiment, and it was verified that the transductant could exploit the same carbon sources as the donor strain. From these experiments, I concluded that the phenotypes of all the evolved strains result from mutations in the ebgA gene.
An obligatory pathway for directed evolutioii
The observation that lactobionate-utilizing strains (Class
V)
could not be obtained from the Class 0 strain 1B1 suggests that a lactobionate-hydrolyzingebg enzyme cannot arise via a single point mutation. Likewise, the failure to obtain lactobionate-utilizing derivatives of Classes I, 11, or I11 suggests that two mutations are not sufficient. Thus, I concluded that three point mutations in the
ebgA gene are required to produce an enzyme capable of hydrolyzing lactobio- nate. Additionally, since lactobionate is not an inducer of ebg enzyme synthesis (HALL and CLARKE 1977), a mutation in the regulatory gene ebgR is required
ENZYME EVOLUTION IN
E. coli
46
1is
5
x(HALL,
unpublished data), then the frequency with which lac- tobionate utilizers arise would be 1.3x
[that is (3 X 10-9)3 X( 5
Xlo-*)].
This is the equivalent of expecting one lactobionate utilizer in about 10” kilo- grams of E. coli.Evolution does not, of course, proceed by the simultaneous occurrence of sev- eral random mutations in a given individual in a population. Instead evolution occurs via a series of mutations, each of which is fixed in the population either because it confers a selective advantage or by random “drift.” In the laboratory, directed evolution involves powerful selection for each mutation. Thus, although the three mutations that permit lactobionate utilization could occur in any order; it is necessary to select the mutations in a particular order. I n this case the order of selection is:
lactose
Class
I
ebgR-, ebgAo (Class 0) selection +phenylgalactoside selection
ebgR+
lactobionate
Class
V.
selection
’
ClassIV
lactulose selection
’
The first two steps of this pathway are interchangeable, but the remaining order is apparently obligatory. That the order is important is illustrated by the follow- ing example: note that if the first €unction selected (from an ebgR- strain) is lactulose utilization, Class I11 strains are obtained. These strains, like Class
Iv
strains, can grow on both lactose and lactulose. Yet Class I11 strains do not yield lactobionate utilizing derivatives. Even if lactose utilization is the first function selected, the lactose utilizing mutants must be screened to eliminate lactulose utilizers (Class II), if further selection for lactobionate utilization is to be SUC- cessful. Class I1 and Class I11 strains can be considered “dead ends” with respect to evolving lactobionate utilization, simply because there is no known way to select the double mutant allele that can give rise to the triplemutant,lactobionate- using allele. Thus the pathway presented is an obligatory pathway for the evolu- tion of lactobionate utilization via the ebg enzyme.The effects of “new” functions on “old‘? functions
I n examining the effect of newly evolved functions on the old function carried out by a n enzyme, it would be ideal to ask how well evolved ebg enzymes carry out the function determined by the ancestral, or unevolved, enzyme ebg”.
Unfortunately, the natural function of ebg enzyme is unknown, so that this is not yet possible. Instead, any allele that specifies an enzyme with a metabolically significant function and can give rise to another allele with a n additional met- abolically significant function can be considered as specifying a n old function, and the derivative allele a new function. Thus, we can compare Class
I
strains with their Class IV derivatives and Class IV strains with their ClassV
derivatives to ask how new functions affect old functions. Figure 2 shows the “pedigrees”462 B. G . HALL
181
( w i l d type ebgA allele 1 I5 A 2 SJ-17
FIGURE 2.-Pedigree of ebgA alleles in evolved strains. Strain 1B1 carries the wild type allele, ebgAO. Strains on line one have a single mutation in the ebgA gene; those i n line two have two mutations in ebgA; those in line three have three mutations in ebgA. All strains are
ebgR-. Strains enclosed in rectangles were selected for growth on lactose; those enclosed in diamonds were selected for growth on lactulose; those in circles were selected for growth on lactobionate. This pedigree shows only the descent of the ebgA gene; i.e., strains SJ-17, A2, 5A2 and D2 were not derived directly from IBI, but their ebgA alleles were derived directly from the ebgA allele carried in I B I .
Class I strains (A2 and D2) utilize lactose, but not lactulose. They also utilize galactose-arabinoside, but very inefficiently. When the new function, lactulose utilization, is selected, eight of the nine derivative strains showed about a 20% decreased growth rate on lactose (see text describing selection of Class IV strains). I n contrast, these same strains showed a 400% increase in their growth rate on galactose-arabinoside. I n this case, a new function has been added at the cost of a minor decrease in one old function (lactose use) and with the bonus of a substantial increase in another old function (galactose-arabinoside use). Overall, it would appear that Class
I
strains do not pay a high price in evolving the new function lactulose utilization. Although they grow at only 80% of the parental rate on lactose, Class IV strains also grow at about 80% of the rate that Class 11 and I11 strains do on lactulose. Note that Class I1 and I11 strains grow only 50%as fast on lactose as do Class IV strains.
We can also consider the effects of selecting for a n improvement i n a n old function, rather than selecting for a new function.
Strain SJ-17, a Class I1 strain, grows slowly on lactose ( a = 0.178 hr-l). A repetitive dilution method was employed to select a derivative of SJ-17 with a significantly improved growth rate on lactose.
M
ENZYME EVOLUTION IN
E .
coli 463IPTG
(to induce synthesis of the lac permease), diluted the next day 500-fold into lactose-minimal medium, and shaken at 37". Two days were required for the culture to become turbid. From the initial turbidity, final turbidity and the elapsed time, the estimated average growth rate during round one of selection was = 0.16 hr'. The round one culture was again diluted 500-fold into lactose minimal medium and shaken until turbid. At the end of each round of selection, the average growth rate during that round was estimated. The growth rate, a7in six rounds of selection progressively increased as follows: 0.16, 0.17, 0.21, 0.30,0.35, and 0.37.
At the end of six rounds, the culture was streaked onto lactose minimal medium and ten clones were purified. All clones grew in lactose minimal medium with rates between a = 0.3 and a! = 0.4 h r l . One of these, named SJ-17-1, was chosen f o r further study.
To be sure that the increased growth rate exhibited by SJ-17-1 was due to a mutation in the ebgA gene, phage P I was grown on SJ-17-1 and used to transduce strain SJ-7. Lactose-utilizing transductants were selected, and scored for co- transduction of the toZC+ allele, as before. The co-transduction frequency was 0.16. Three transductants were purified, and the growth rates on lactose were between a! = 0.37 and 01 = 0.40 h r l .
Strain SJ-17, a Class
I1
strain, grows slowly on lactose (a = 0.1 78 h r l ) . Atwice as fast as the parental strain, SJ-17. However that increased growth rate on lactose was accompanied by a five-fold decrease in the growth rate on lactulose to a 0.047
*
0.005. The growth rate on galactose-arabinoside was improved a little more than two-fold to a! = 0.043*
0.005. Gene duplication as a solutionto the problem of increasing the amount of a particular enzyme activity was observed in the selection of hyper-producers of @-galactosidase via growth of
E. coti in a lactose-limited chemostat (HORIUCHI, TOMIZAWA and
NOVICK
1962). The observation that the growth rate on lactose increased, but that the growth rate on lactulose dramatically decreased, argues strongly that strain SJ-1 7 did not improve its growth rate on lactose via gene duplication. Thus, it seems likely that strain SJ-17-1 has arisen via a second mutation in the ebgA gene. In this case, the parental strain seems to have paid a heavy,price for increased effective- ness of lactose utilization, since it has virtually lost lactulose utilization and gained little in terms of galactose-arabinoside utilization.The selection of lactobionate utilization from Class IV strains affords a wider base upon which to compare the parental with the derived strains, since Class
464 B. G . HALL
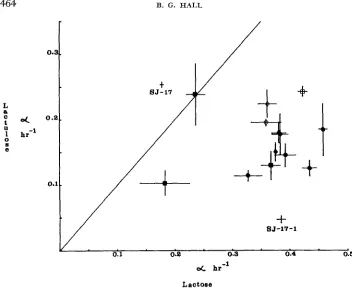
L ao t ose
FIGURE 3.-Growth rates of Class IV and derived Class V strains o n lactulose us. lactose. Growth rates are displayed as a cross, in which the intersection of the two lines is the mean growth rates on lactose and lactulose; and in which the lengths of the lines indicate the 95%
confidence interval about that mean. Symbols at the intersections of lines. Open circle: A203;
Closed circles: Class V strains derived from A203; Open square: A207; Closed squares: Class V strains derived from A207; Open diamond: PW201; Closed diamonds: Class V strains derived from PW201.
derivatives all exhibit a reduced growth rate on lactose, two exhibit a reduced rate on lactose, two exhibit a reduced rate on lactulose, and two a reduced rate on galactose-arabinoside. D21 strains show no change in growth rate on lactose, a reduced rate on lactulose, and one strain shows an increased rate on galactose- arabinoside. Thus, among the lactobionate-utilizing derivatives of Class
IV
strains, virtually every possible effect is seen upon the functions (lactose, lactulose, and galactose-arabinoside utilization). Among these strains, there were 18 instances of reduction in the efficiency of functions, nine cases of no change, and three cases of increased efficiency of function when a new function (lac- tobionate utilization) evolved.
Conclusions
ENZYME EVOLUTION IN E. coli 465
ebgR. The pathway requires that the mutations are selected in a particular order. This is not because these mutations must occur in a particular order, but because selection must be applied to build up a sufficiently large population of a singly mutant strain to detect a double mutant, etc.
When various selective pressures were applied, changing the ecological niche by offering a new carbon source, E . coli responded by evolving ebg enzyme SO
that it could carry out the required new functions.
In
many cases this resulted in a reduced efficiency with which ebg enzyme carried out its old functions; how- ever, in a significant number of cases, either no change was observed or there was an increase in the efficiency with which an old function was carried out. These results make it clear that evolution of a new function does not necessarily affect the old function of an enzyme adversely. Thus “divergence,” in the sense of the initial appearance of two functions from a single function, need not always be preceded by duplication of genes specifying vital functions.On the other hand, it is not clear that evolution of two “optimally adapted” functions from one “optimally adapted” function can occur without gene dupli- cation. None of the five strains “bested adapted’’ to lactose utilization is also “best adapted” to either lactulose or lactobionate utilization. Neither is the best lactulose utilizing strain best adapted to lactose or lactobionate utilization; or the best lactobionate utilizer best adapted to either lactose or lactulose. The only exception is that the best galactose-arabinoside utilizing strain is also among the best lactose utilizers. However, since no selection for galactose-arabinoside uti- lization was employed, this is probably fortuitous. Experiments to determine whether two functions can be “optimized” simultaneously without gene duplica- tion are in progress.
LITERATURE CITED
CAMPBELL, J. H., J. A. LENGYEL and J. LANGRIDGE, 1973
HALL, B. G., 1976
Evolution of a second gene for P-galactosidase in Escherichia coli. Proc. Nat. Acad. Sci. U.S. 70: 1841-1845.
Experimental evolution of a new enzymatic function. Kinetic analysis of the ancestral (ebgo) and evolved ( e b g + ) enzymes. J. Mol. Biol. 107: 71-84. 1977 Number of mutations required to evolve a new lactase function in Escherichia coli. J. Bacteriol. 129: 540-543.
HALL, B. G. and N. D. CLARKE, 1977 Regulation of newly evolved enzymes. 111. Evolution of
the ebg repressor during selection for enhanced lactase activity. Genetics 85: 193-201.
HALL, B. G. and D. L. HARTL, 1974 Regulation of newly evolved enzymes. I. Selection of a novel lactase regulated by lactose in Escherichia coli. Genetics 76: 391-400.
-,
1975 Regulation of newly evolved enzymes. 11. The ebg repressor. Genetics 81 : 427-435.Second naturally occurring p-galactosidase in E. coli.
Nature 248: 152-153.
lsolation and properties of bacteria capable of high rate of P-galactosidase synthesis. Biochem. Biophys. Acta 55: 152-163.
-
,HARTL, D. L. and B. G. HALL, 1974
HORIUCHI, T., TOMIZAWA, J. and A. NOVICK, 1962
JACOB, F., 1977 MILLER, J. H., 1972
York.
Evolution and tinkering. Science 1%: 1161.