| REVIEW
Mitochondrial Replacement Therapy: Are Mito-nuclear
Interactions Likely To Be a Problem?
Adam Eyre-Walker1
School of Life Sciences, University of Sussex, Brighton BN1 9QG, United Kingdom ORCID ID: 0000-0001-5527-8729 (A.E.-W.)
ABSTRACTIt has been suggested that deleterious interactions between the mitochondrial and nuclear genomes could pose a problem for mitochondrial replacement therapy (MRT). This is because the mitochondrial genome is placed in a novel nuclear environment using this technique. In contrast, it is inherited with half the mother’s genome during normal reproduction, a genome that it is relatively compatible with, since the mother is alive. Here, I review the evidence of whether mito-nuclear interactions are likely to pose a problem for MRT. The majority of the available experimental evidence, both in humans and other species, suggests that MRT is not harmful. These results are consistent with population genetic theory, which predicts that deleterious mito-nuclear interactions are unlikely to be much more prevalent in individuals born to MRT than normal reproduction, particularly in a species such as humans with low population differentiation. This is because selection is unlikely to be strong enough to establish significant linkage disequilibrium between the mitochondrial and nuclear genomes. These results are supported by a meta-analysis of 231 cases, from a variety of animals, in which the mitochondrial DNA (mtDNA) from one strain has been introgressed into the nuclear background of another strain of the same species. Overall, there is little tendency for introgression of mtDNA to be harmful.
KEYWORDScoadaptation; mito-nuclear incompatibility; mito-nuclear interaction; mitochondrial replacement therapy
A
number of diseases, such as Leber’s Hereditary Neurop-athy, Leigh’s syndrome, and Myoclonic Epilepsy with Ragged Red Fibers, are known to be caused by mutations in mtDNA (Tuppenet al.2010). These diseases are debilitating and have no cure. Furthermore, due to the maternal inheri-tance of mitochondrial DNA (mtDNA), a woman with one of these diseases has a high probability of transmitting the dis-ease to her offspring; the probability is not 100% because she may be heteroplasmic for the disease-causing mutation; therefore, by chance, she may have offspring with a low pro-portion of her mitochondria with the pathogenic mutation (Tuppenet al.2010). However, it is now possible to transfer her nucleus to the enucleated egg of a donor, effectively re-placing her mitochondria with those from a donor who does not carry the pathogenic mitochondrial mutation, and hence eliminate the chance that her offspring will be affected by the disease (Wolf et al.2015). This technique is known asmitochondrial replacement (MR) or mitochondrial replace-ment therapy (MRT) (Wolfet al.2015). The procedure was approved in 2015 by the British government for use in the clinic, and thefirst individual conceived by this technique has been reported to have been born in Mexico (Zhang et al.
2016). However, concerns have been raised about the poten-tial deleterious genetic effects of mixing the nuclear genome of the recipient with mitochondrial genome of the donor due to potential incompatibilities between the mitochondrial and nuclear genomes (Reinhardtet al.2013; Gemmell and Wolff 2015; Hamilton 2015; Morrowet al.2015). Others have ar-gued that MRT is not very dissimilar to normal sex and that such incompatibilities are unlikely to be an issue (Chinnery
et al. 2014) (http://www.hfea.gov.uk/8178.html). Here, I discuss whether such incompatibilities are likely to be a prob-lem in humans from a population and evolutionary genetic viewpoint.
In thinking about MRT, it is important to differentiate between two possible sources of deleterious genetic effects associated with the process that have been conflated, to some degree, by previous commentators (Reinhardtet al.2013). First, there are effects that are independent of nuclear back-ground,e.g., mtDNA X is always morefit than mtDNA Y in all Copyright © 2017 by the Genetics Society of America
doi:https://doi.org/10.1534/genetics.116.196436
Manuscript received November 10, 2016; accepted for publication February 8, 2017 Supplemental material is available online atwww.genetics.org/lookup/suppl/doi:10. 1534/genetics.116.196436/-/DC1.
nuclear backgrounds, and to the same degree. It seems unlikely that these effects will be a problem for MR since it is simple to avoid them; choose a donor who is healthy, and if you are concerned that the mtDNA might perform differently in a male, check that she has had a healthy male child. Second, there is potential for the mtDNA to interact with the nuclear genome such that mtDNA X is
fitter than Y in nuclear background A but (relatively) worse in nuclear background B, i.e., there is epistasis. These interactions are more problematic because they are essentially unpredictable, and it is these latter effects that chiefly concern us here.
Interactions between the nuclear and mitochondrial ge-nomes are expected because the genes involved in mitochon-drial function are found in both the nucleus and the mitochondria. In humans, mtDNA contains 36 genes whereas the nuclear DNA (nuDNA) encodes.1158 genes that have been shown to localize to the mitochondrion (Calvo et al.
2016), and among those genes that encode for the electron transport system, 13 are found in mtDNA and70 in nuDNA (Shoubridge 2001). Indeed, there are a number of cases in which it has been demonstrated that the nuclear background affects the phenotype of a mitochondrial mutation, or vice versa, [for example Bykhovskayaet al.(2004); Ballanaet al.
(2007); Bonaitiet al.(2010)].
Deleterious interactions between the nuclear and mi-tochondrial genomes are expected to be, on average, more prevalent during MR because the mitochondria will expe-rience a completely novel nuclear background in MR, whereas in normal sex the mitochondria will be inherited with half the mother’s nuclear genome, a genome with which the mitochondria have shown themselves to be rel-atively compatible, because the mother is alive. Neverthe-less, during normal sex, the mitochondria will experience a seminovel nuclear environment because they will not have previously experienced the half of the nuclear ge-nome from the father (unless there is a high degree of inbreeding). So, the critical question is not whether dele-terious mito-nuclear interactions (DMNI) are more likely under MR, but how much more likely they are. If DMNIs are common, strong, and MR substantially increases their frequency, then we should be concerned about them; but if they are rare, weak, or MR has little effect on their fre-quency, then they are not something that should concern us.
Within Populations
Both sexes
The association between alleles that are relatively compat-ible within the mother can be established by two processes. First, within-a-population selection against DMNIs estab-lishes linkage disequilibrium (LD) between the interacting mutations; for example, if there is a dominant embryoni-cally lethal interaction between a nuclear and
mitochon-drial mutation, then the mother will never carry the two variants that interact. However, this LD is broken down each generation by segregation. The rate of recombination is 0.5 between a nuclear and mitochondrial locus and only selec-tion of a similar magnitude (0:1) is likely to maintain substantial LD between the nuclear and mitochondrial ge-nomes. Analysis of simple models suggests that DMNIs will be, at most, only twice as common in individuals born through MR compared to individuals born through normal sex (Appendix); this occurs when the nuclear variant is completely dominant and the interaction is lethal. If the interaction is not strongly selected or the nuclear mutation is only partially dominant or recessive then there will not be strong LD between the nuclear and mitochondrial ge-nomes, and even if DMNIs are common, they will only be a slightly more frequent in MR relative to normal sex.
Such strong selection is thought to be rare in humans and most multi-cellular eukaryotes (Eyre-Walker and Keightley 2007). This is evident in humans from the fact that only 2– 3% of human babies are born with visible birth defects (http://www.cdc.gov/ncbddd/birthdefects/data.html and http://www.eurocat-network.eu/accessprevalencedata/ prevalencetables), and only 2% of couples are primary in-fertile (i.e., cannot have at least one child within 5 years of trying) (Mascarenhaset al.2012). So, unless most of these problems are due to genetics and in particular DMNIs, which seems unlikely and for which there is no evidence, strongly selected DMNIs must be rare. The limited available evidence fromDrosophila melanogastersuggests that inter-actions between the mitochondrial and nuclear genomes involve many mutations of small effect (Camus et al.
2012). Furthermore, most mutations of large effect tend to be recessive (Simmons and Crow 1977), which will greatly reduce the level of LD between the mitochondrial and nuclear genomes (Appendix).
There is one form of selection that can increase the likelihood of DMNIs substantially in MR relative to normal individuals. As Morrowet al.(2015) point out, if there is selection on the oocyte before fertilization, then some DMNIs might be removed during normal sex that would not be removed by MRT. A variant of this model could allow the frequency of DMNIs to be zero under normal sex but nonzero under MRT; if a mutation is expressed in both the haploid and diploid stages, is lethal at both stages but recessive in the diploid stage, then a mother will never pass the interacting alleles on to her offspring and her offspring will never suffer from the DMNI, be-cause the nuclear allele is recessive. However, under MR it is possible to introduce the mitochondrial allele into an individual who is homozygous for the interacting nuclear mutation.
Males and postreproductive characters
to have more deleterious effects in males, because mtDNA is not (usually) inherited from males and hence selection in males can have no effect on the frequency of mitochon-drial mutations. Therefore, there is an expectation that mutations that are advantageous or neutral in females, but deleterious in males, can accumulate; this has been termed the“Mother’s Curse.”Indeed, it has been demon-strated that genetic variation in mtDNA affects several traits more in males than in females, such as gene expres-sion (Innocenti et al. 2011) and ageing (Camus et al.
2012), although a recent meta-analysis found no evidence for this (Dobleret al.2014). However, MRT will not exac-erbate these effects if the mitochondrial and nuclear ge-nomes are taken from the same population, for the simple reason that selection in males cannot generate the LD be-tween the mitochondrial and nuclear genomes that make DMNIs less likely in normal sex. For similar reasons, DMNIs might affect postreproductive characters, but these will not be more common among MR individuals because selection cannot generate LD in individuals who havefi n-ished reproducing.
Between Populations
The second process that can lead to an association between mitochondrial and nuclear variants in the mother is popu-lation substructure. This can cause there to be incompatibil-ities between individuals that are sampled from different populations if there is limited migration. For example, let us imagine that we have a nuclear locus with two allelesNand
n, and a mitochondrial locus with allelesMand m; let us assume that the ancestral population has the genotype of
NN M, that thenallele is either advantageous or neutral if the mitochondrial locus isM, and that themallele is advan-tageous or neutral if the nuclear locus is NN, but that the genotypes Nn mand nn mare deleterious (i.e., there is a negative epistatic interaction between thenandmalleles). Under these conditions, a single panmictic population is very unlikely to evolve fromNN Mtonn m, unless the pop-ulation size is small and genetic drift is effective. However, if the population splits into two or more subpopulations then thefirst could evolve to benn Mand the second toNN m(or vice versa). If we cross individuals from these populations, we will generate individuals who areNn mwho suffer from the negative epistatic interaction between allelesnandm. With limited migration, and as time progresses, these in-compatibilities can accumulate to an extent that hybrids between populations are extremely unfit; the accumulation of these incompatibilities can eventually lead to speciation, at which point they are referred to as Dobzhansky–Muller incompatabilities.
Human populations are well-known to show little evidence of population differentiation (Rosenberg et al.
2002), although there are clearly genetic differences, since there is regional variation in morphology and phys-iology. However, it seems unlikely that many strongly
DMNIs have accumulated between human populations be-cause there is no evidence that interbreeding between populations leads to a decrease in fitness. There is also no evidence of significantly more LD between human pop-ulations between mitochondrial and nuclear genes than one would expect given the limited levels of population substructure that exist in humans (Sloanet al.2015), al-though it should be appreciated that LD would have had to be very strong between substantial numbers of mitochon-drial and nuclear polymorphisms for this test to detect an effect.
Experimental Evidence
MR
MR has been performed in three species: humans, macaques, and mice. In humans, the experiments have generally only been allowed to proceed to the blastocyst stage of develop-ment, but in one case the pregnancy was allowed to go to term (Zhang et al. 2016). In several of these experiments, MR zygotes were significantly less likely to develop to the blas-tocyst stage than unmanipulated controls (Tachibanaet al.
2009; Cravenet al.2010; Hyslopet al.2016). This could be due to the manipulation of the oocyte or the presence of DMNIs. The evidence is inconclusive. In one study, there was evidence that the problems were at least in part due to the experimental manipulation of the oocytes (Tachibana
et al. 2009). However, in the most informative experiment to date, Hyslop et al.(2016) performed two forms of MR; they reinjected the nucleus back into the oocyte it had been removed from [autologous pronuclear transfer (PNT)] and they performed reciprocal swaps between eggs (heterolo-gous PNT). They found that zygotes derived by autolo(heterolo-gous PNT were as likely to develop to the blastocyst stage as un-manipulated controls. However, the products of heterologous PNT were almost 50% less likely to develop to the blastocyst stage than either the unmanipulated controls or the autolo-gous PNT-derived embryos (one-tailed Fisher’s exact test
P= 0.009 andP= 0.027 for unmanipulated and autologous controls, respectively). Hyslopet al.(2016) suggest that the difference might arise because one of the eggs involved in the heterologous transfer had been previously frozen, whereas all other eggs used in the experiment were fresh. Unfortu-nately, they did not track the survival of the two products of heterologous PNT separately (i.e., frozenvs.fresh eggs used as the donor). Subsequent analysis showed that both autol-ogous and heterolautol-ogous PNT-derived blastocysts were of sim-ilar quality to unmanipulated controls and that gene expression profiles did not differ (Hyslopet al.2016). Anal-ysis of mitochondrial function in other experiments of MR-derived stem cells andfibroblasts showed either no sig-nificant difference to non-MR controls (Paull et al. 2013; Yamada et al.2016), or to significant differences in which the MR cells could be significantly better or worse (Kanget al.
correlation between mitochondrial performance and the di-vergence between the donor and recipient mtDNAs (Kang
et al. 2016). Overall, there seems little evidence that MRT is harmful in humans, but the experiment performed by Hyslop et al. (2016) suggests that further experiments are warranted.
MR has also been performed in macaques, with four indi-viduals born to this technique (Tachibana et al. 2009). At 3 years of age, these individuals appear to have developed normally with normal mitochondrial function (Tachibana
et al.2013) despite the macaques apparently coming from different subspecies (http://www.hfea.gov.uk/8178.html). In mice, it has been shown that MRT can be used successfully to replace mitochondria with respiratory defects (Satoet al.
2005) and that MRT yields healthy offspring, with normal respiration, as frequently asin vitrofertilization (Wanget al.
2014).
Conplastic strains
Although, MR has only been performed in three species and very few mito-nuclear combinations have been studied, mtDNA has been introduced into various nuclear back-grounds, by repeated backcrossing, in a number of other animal species; these strains are known as conplastic. This effectively performs MR, although it should be appreciated that very strong DMNIs are likely to be removed by selection during backcrossing.
The effects of introgressing mtDNA from one strain in another are mixed. In some cases, the introgression of mtDNA into a strain leads to a loss offitness; for example, in D. melanogaster, the introgression of a mtDNA from Brownsville, Texas, onto a standard lab background led to male sterility (Clancy 2008). These deleterious effects have led to the suggestion that there is coadaptation
be-tween the mitochondrial and nuclear genomes, and that MR could be harmful (Reinhardtet al.2013; Gemmell and Wolff 2015; Hamilton 2015; Morrow et al. 2015). How-ever, there are also cases in which fitness and health are enhanced in conplastic strains; for example, the introgres-sion of the mtDNA from the mouse strain NZB/OlaHsd into C57BL/6 led to an increase in longevity relative to the C57BL/6 strain with its own mtDNA (Latorre-Pellicer
et al.2016).
To investigate whether this form of MR tends to be dele-terious on average, estimates of the effect of replacing the “native”with“foreign”mitochondria were compiled from the literature, where native mtDNA is isolated with the nuclear genome, and foreign mtDNA is sampled from the same spe-cies, but alongside a different nuclear genome. Note that, in this case, we are not differentiating between additive and epistatic effects; hence, low fitness in the conplastic strain might be because the mtDNA reduces fitness in all strains that it is in, or it might be because it has an interaction with a specific nuclear background. The data set comprised exper-iments in animals in which an organismal-level trait had been measured; I ignored studies in which only mitochondrial function had been studied, since it is organismal-level traits that we are ultimately interested in and there is a poor cor-relation between mitochondrial function and organismalfi t-ness [e.g., see Latorre-Pelliceret al.(2016)]. The magnitude of the effect was quantified as the proportional difference between the lines with the foreign and native mtDNAs (i.e., the difference divided by the mean, calculated such that pos-itive values represent the case when the line with the foreign mtDNA was better than the line with the native mtDNA); as such, only studies using an untransformed scale were in-cluded (e.g., residual sperm length was not included). Many of the traits that have been studied are probably under
stabilizing selection in natural populations and hence it can be difficult to judge whether an increase or decrease in the trait is beneficial; therefore, we assigned the beneficial direc-tion based on what we might expect if mitochondrial funcdirec-tion was seriously compromised;i.e., individuals with very poorly functioning mitochondria might be expected to have a lower chance of survival, develop more slowly, be smaller, and have lower fertility. For some traits, we took the direction as that which would be desirable to humans—e.g., increased life span and ability to learn—but for some traits, such as explor-atory behavior in mice, it was difficult to assign a beneficial direction; these studies were ignored. Where the same trait had been assayed multiple times or at different temperatures, we took the middle assay time and temperature to reduce nonindependence.
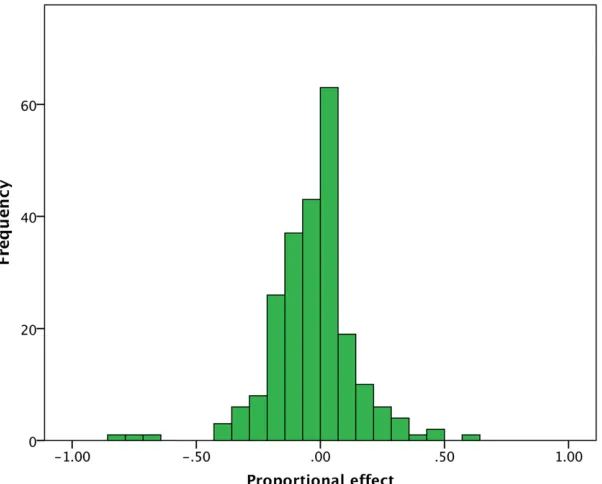
The data set comprised 231 estimates, the vast majority of which came from either Drosophila species or the beetle
Callosobruchus maculatus (Supplemental Material, Table S1). There were also a moderate number of estimates from the house mouseMus mus domesticus. It should be appreci-ated that, while there are a substantial number of estimates, they come from a limited set of lines; for example, the 92 es-timates from C. maculatus come from 25 lines that have been repeatedly used. Therefore, there is substantial non-independence in the data and, as a consequence, no formal statistical analysis was performed. It should also be noted that the vast majority of estimates come from inbred lines; this may exacerbate any interactions between the nuclear and mitochondrial genomes, since mutations with large ef-fects tend to be recessive (Simmons and Crow 1977). The extent of inbreeding depression in some of these data are evident in the work of Clancy on female longevity in
D. melanogaster (Clancy 2008). He backcrossed mtDNAs from different localities on to a standard homozygous nu-clear background; the mean longevity for mtDNAs from Alstonville, Dahomey, and Japan was 27.4 days. However, when those lines were made heterozygous by crossing to other strains offlies, but not the line the mtDNA was sam-pled from, the mean longevity increased to 38.9 days, a 42% increase.
Taking all the data together, there is a slight tendency for conplastic strains to be less fit than native strains (Figure 1); 54% (125) of the comparisons show a nega-tive effect of introgression, 44% (102) show a posinega-tive effect, and 2% (4) show no discernible effect (Figure 1).
However, the average effect is very close to zero at23.4% and the majority of effects are small; 56% of the estimates are,10%.
If we consider each species individually, for which we have.10 estimates, we observe qualitatively similar patterns (Table 1). There is one exception, the copepodTigriopus cal-ifornicus. In this species, the strain with the native mtDNA is alwaysfitter than the strain with the foreign mtDNA, and the average effect size is .10%. It is notable that this species shows extremely high levels of population differentiation (Edmands and Harrison 2003); FST between the Southern populations of this species, from where the lines were sam-pled, is 0.81 (Edmands and Harrison 2003). However, high levels of population differentiation do not always result in consistently negative consequences of introgressing mtDNA from one population into another, as illustrated byC. macu-latus, which shows high levels of population differentiation (Tudaet al.2014) but no overall trend toward negative ef-fects (Table 1). It is worth noting thatFSTin humans is very low at 0.05 (Rosenberget al.2002), similar to the levels observed in D. melanogaster(Verspoor and Haddrill 2011), which also show no consistent pattern in the effects of introgressing mtDNA into different nuclear backgrounds (Table 1).
It has been hypothesized that the effects of MR might be more apparent in males (Reinhardtet al.2013; Morrowet al.
2015) because mtDNA is not inherited from this sex, and hence mutations that are deleterious in males, but neutral or beneficial in females, can accumulate. There is little evi-dence that effects of introgression are worse in males; the mean effect in males is 23.3% (86 estimates) and in fe-males it is 22.6% (93 estimates) (Figure 2). There is also little evidence that sampling the mtDNAs from different populations is more deleterious than taking them from the same population; the mean effect for between popu-lations is23.7% (181 estimates) and within it is20.0% (34 estimates).
The presence of a deleterious interaction between the nuclear and mitochondrial genomes is often interpreted as evidence of coadaptation, i.e., that the two genomes have coevolved together. However, it is possible that such cases simply represent segregating mutations that have a deleteri-ous interaction. For example, the mtDNA inD. melanogaster
from Brownsville, Texas, might cause male sterility on the w1118 nuclear background, because the nuclear and
Table 1 The proportion of cases in which the strain with the native mtDNA is better than the strain with the foreign mtDNA, and the average proportional effect (negative values indicate the foreign strain has lowerfitness)
Species
Number of Comparisons
Proportion of Cases in which Native Is Better
Average Proportional Effect (%)
Callosobruchus maculatus 92 0.41 21.5
Drosophila melanogaster 66 0.59 23.4
Drosophila subobscura 32 0.44 20.1
Mus musculus domesticus 14 0.64 25.0
mitochondrial genomes have coevolved and diverged from the population that w1118 was sampled from; or it might simply be that there is a mitochondrial mutation segregat-ing in Texas that is incompatible with the ssegregat-ingle w1118 genome. To establish coevolution and coadaptation it is necessary, but not sufficient, to show that all or most mtDNAs taken from Texas are compatible with all or most nuclear genomes taken from Texas, and incompatible with all or most genomes sampled from the w1118 population. This type of experiment is rarely done. The above meta-analysis suggests that there is little evidence for coadap-tation between the mitochondrial and nuclear genomes because there is little tendency for the introgression of a foreign mtDNA onto a nuclear background to be deleterious.
Haplotype Matching
It is potentially possible to completely eliminate the prospect of DMNIs by using donor mtDNA that is identical to the recipient’s in all but the pathogenic mutation, by sampling the donor from a close maternal relative. More generally, it has been suggested that sampling the donor mtDNA from the same haplogroup as the recipient might be a worthwhile strategy (Vogel 2014; Wolfet al.2015). The fact that there is little evidence that transferring a mtDNA from one nuclear background to another, either by MR or introgression, is del-eterious, suggests that this is not necessary in terms of DMNIs. However, there is evidence from human MR experi-ments that the maternal mtDNA can outcompete the donor mtDNA leading to loss of the donor mtDNA (Kang et al.
2016). In some cases, this reversion appears to be due to a
specific polymorphism in the control region of the mtDNA (Kanget al.2016).
Summary
It has been suggested that placing mtDNA into a completely novel genetic background, as will happen in MR, may lead to harmful effects because of deleterious interactions between the mitochon-drial and nuclear genomes. There is one experiment that suggests that this might be the case; Hyslop et al. (2016) found that oocytes that had received their nucleus from a donor were less likely to develop into a blastocyst than oocytes who had had their own nucleus injected back into them. This might be due to DMNIs, but it also could be due to the fact that the one of the oocytes had been frozen in the heterologous treatment. Other-wise, there is little evidence that MR is harmful because of DMNIs. MR in humans leads to blastocysts with normal gene expression (Hyslop et al.2016) and OXPHOS function (Paull
et al. 2013; Kang et al. 2016; Yamada et al. 2016). MRT also appears to have no deleterious effects in macaques (Tachibana et al.2009, 2013) and mice (Wang et al.2014). Furthermore, the introgression of mtDNA into a new “ for-eign”nuclear background in other species does not appear to be deleterious on average. These results are consistent with population genetic theory. Only if selection against DMNIs is very strong will they be more frequent in individu-als born using MR than normal reproduction if the species is relatively panmictic, and even then, the effects are at most twice as likely. Finally, it is worth noting that the risk of DMNIs from MRT must be much smaller than the risks of having an affected child through normal reproduction, if the mother has a pathogenic mitochondrial mutation.
Acknowledgments
I am grateful to Damian Dowling, Milhailo Jelic, and Zorana Novicic for sending me their results.
Literature Cited
Ballana, E., J. M. Mercader, N. Fischel-Ghodsian, and X. Estivill,
2007 MRPS18CP2 alleles and DEFA3 absence as putative
chromosome 8p23.1 modifiers of hearing loss due to mtDNA
mutation A1555G in the 12S rRNA gene. BMC Med. Genet. 8: 81.
Bonaiti, B., M. Olsson, U. Hellman, O. Suhr, C. Bonaiti-Pellie
et al., 2010 TTR familial amyloid polyneuropathy: does a mitochondrial polymorphism entirely explain the
parent-of-origin difference in penetrance? Eur. J. Hum. Genet. 18: 948–
952.
Bykhovskaya, Y., E. Mengesha, D. Wang, H. Yang, X. Estivill
et al., 2004 Human mitochondrial transcription factor B1
as a modifier gene for hearing loss associated with the
mitochondrial A1555G mutation. Mol. Genet. Metab. 82:
27–32.
Calvo, S. E., K. R. Clauser, and V. K. Mootha, 2016 MitoCarta2.0:
an updated inventory of mammalian mitochondrial proteins.
Nucleic Acids Res. 44: D1251–D1257.
Camus, M. F., D. J. Clancy, and D. K. Dowling, 2012 Mitochondria,
maternal inheritance, and male aging. Curr. Biol. 22: 1717–1721.
Chinnery, P. F., L. Craven, S. Mitalipov, J. B. Stewart, M. Herbert
et al., 2014 The challenges of mitochondrial replacement. PLoS Genet. 10: e1004315.
Clancy, D. J., 2008 Variation in mitochondrial genotype has
sub-stantial lifespan effects which may be modulated by nuclear
background. Aging Cell 7: 795–804.
Craven, L., H. A. Tuppen, G. D. Greggains, S. J. Harbottle, J. L.
Murphy et al., 2010 Pronuclear transfer in human embryos
to prevent transmission of mitochondrial DNA disease. Nature
465: 82–85.
Dobler, R., B. Rogell, F. Budar, and D. K. Dowling, 2014 A
meta-analysis of the strength and nature of cytoplasmic genetic
ef-fects. J. Evol. Biol. 27: 2021–2034.
Edmands, S., and J. S. Harrison, 2003 Molecular and
quanti-tative trait variation within and among populations of the
intertidal copepod Tigriopus californicus. Evolution 57:
2277–2285.
Eyre-Walker, A., and P. D. Keightley, 2007 The distribution of
fitness effects of new mutations. Nat. Rev. Genet. 8: 610–
618.
Gemmell, N., and J. N. Wolff, 2015 Mitochondrial replacement
therapy: cautiously replace the master manipulator. BioEssays
37: 584–585.
Hamilton, G., 2015 The mitochondria mystery. Nature 525: 444–
446.
Hyslop, L. A., P. Blakeley, L. Craven, J. Richardson, N. M. Fogarty
et al., 2016 Towards clinical application of pronuclear trans-fer to prevent mitochondrial DNA disease. Nature 534:
383–386.
Innocenti, P., E. H. Morrow, and D. K. Dowling, 2011 Experimental
evidence supports a sex-specific selective sieve in mitochondrial
genome evolution. Science 332: 845–848.
Kang, E., J. Wu, N. M. Gutierrez, A. Koski, R. Tippner-Hedgeset al.,
2016 Mitochondrial replacement in human oocytes
carry-ing pathogenic mitochondrial DNA mutations. Nature 540:
270–275.
Latorre-Pellicer, A., R. Moreno-Loshuertos, A. V. Lechuga-Vieco, F.
Sanchez-Cabo, C. Torrojaet al., 2016 Mitochondrial and
nu-clear DNA matching shapes metabolism and healthy ageing.
Nature 535: 561–565.
Mascarenhas, M. N., S. R. Flaxman, T. Boerma, S. Vanderpoel, and
G. A. Stevens, 2012 National, regional, and global trends in
infertility prevalence since 1990: a systematic analysis of 277 health surveys. PLoS Med. 9: e1001356.
Morrow, E. H., K. Reinhardt, J. N. Wolff, and D. K. Dowling,
2015 Risks inherent to mitochondrial replacement. EMBO
Rep. 16: 541–544.
Paull, D., V. Emmanuele, K. A. Weiss, N. Treff, L. Stewart et al.,
2013 Nuclear genome transfer in human oocytes eliminates
mitochondrial DNA variants. Nature 493: 632–637.
Reinhardt, K., D. K. Dowling, and E. H. Morrow, 2013 Mitochondrial
replacement, evolution, and the clinic. Science 341: 1345–1346.
Rosenberg, N. A., J. K. Pritchard, J. L. Weber, H. M. Cann, K. K. Kidd
et al., 2002 Genetic structure of human populations. Science
298: 2381–2385.
Sato, A., T. Kono, K. Nakada, K. Ishikawa, S. Inoue et al.,
2005 Gene therapy for progeny of mito-mice carrying
patho-genic mtDNA by nuclear transplantation. Proc. Natl. Acad. Sci.
USA 102: 16765–16770.
Shoubridge, E. A., 2001 Nuclear genetic defects of oxidative
phos-phorylation. Hum. Mol. Genet. 10: 2277–2284.
Simmons, M. J., and J. F. Crow, 1977 Mutations affectingfitness
in Drosophila populations. Annu. Rev. Genet. 11: 49–78.
Sloan, D. B., P. D. Fields, and J. C. Havird, 2015 Mitonuclear
linkage disequilibrium in human populations. Proc. Biol. Sci. 282: 20151704.
Tachibana, M., M. Sparman, H. Sritanaudomchai, H. Ma, L. Clepper
et al., 2009 Mitochondrial gene replacement in primate
off-spring and embryonic stem cells. Nature 461: 367–372.
Tachibana, M., P. Amato, M. Sparman, J. Woodward, D. M. Sanchis
et al., 2013 Towards germline gene therapy of inherited
mito-chondrial diseases. Nature 493: 627–631.
Tuda, M., K. Kagoshima, Y. Toquenaga, and G. Arnqvist,
2014 Global genetic differentiation in a cosmopolitan pest of
stored beans: effects of geography, host-plant usage and anthro-pogenic factors. PLoS One 9: e106268.
Tuppen, H. A., E. L. Blakely, D. M. Turnbull, and R. W. Taylor,
2010 Mitochondrial DNA mutations and human disease.
Bio-chim. Biophys. Acta 1797: 113–128.
Verspoor, R. L., and P. R. Haddrill, 2011 Genetic diversity,
popu-lation structure andWolbachiainfection status in a worldwide
sample ofDrosophila melanogasterandD. simulanspopulations.
PLoS One 6: e26318.
Vogel, G., 2014 FDA considers trials of‘three-parent embryos.’
Science 343: 827–828.
Wang, T., H. Sha, D. Ji, H. L. Zhang, D. Chenet al., 2014 Polar
body genome transfer for preventing the transmission of
in-herited mitochondrial diseases. Cell 157: 1591–1604.
Wolf, D. P., N. Mitalipov, and S. Mitalipov, 2015 Mitochondrial
replacement therapy in reproductive medicine. Trends Mol.
Med. 21: 68–76.
Yamada, M., V. Emmanuele, M. J. Sanchez-Quintero, B. Sun, G.
Lalloset al., 2016 Genetic drift can compromise mitochondrial
replacement by nuclear transfer in human oocytes. Cell Stem
Cell 18: 749–754.
Zhang, J., H. Liu, S. Luo, A. Chavez-Badiola, Z. Liu et al.,
2016 First live birth using human oocytes reconstituted by
spindle nuclear transfer for mitochondrial DNA mutation
caus-ing Leigh syndrome. Fertil. Steril. 106: e375–e376.
Appendix
The Frequency of DMNIs in MR and Normal Reproduction
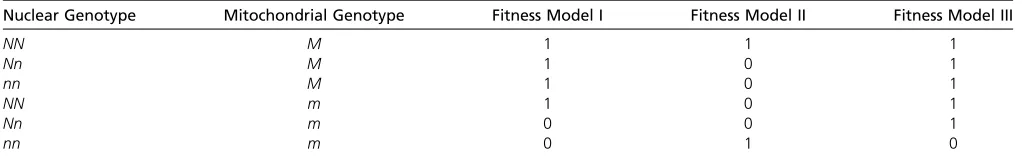
It would seem likely that the biggest difference in the frequency of DMNIs between MR and normal reproduction will occur when the mother cannot pass on the alleles during normal reproduction that interact negatively. For this to happen the interaction must be lethal. Let us consider two scenarios that fulfill this condition. In thefirst, we imagine there is a mitochondrial mutationm, which is neutral unless it is in a nuclear background with the nuclear allelen, in which case the allele is lethal,i.e.there is dominant lethal interaction (model I in Table A1). Consider an individualmallele. Under normal sex, the chance that anm
allele will encounter annallele in the next generation is approximatelyp, the frequency of thenallele in the population, because we know the mother must have been NNm, otherwise she would have been dead. This is only an approximation, because we have ignored thenalleles that have been removed because they found themselves in a zygote that also had them
allele; this is a reasonable approximation, because themand/ornalleles are likely to be at low frequency since the interaction is lethal. In contrast, under MR the chance that themallele encounters annallele is2p(12p) +p2. If thenallele is rare this is 2pand it is,2pif the allele is common. Hence, at most there is twice the chance of generating a genotype with the lethal combination during MR compared to normal reproduction.
In the second scenario, let us imagine that the two genotypesNNMandnnmare viable, and that all other genotypes are inviable (model II in Table A1). We might think of this as a“rescue”model in which thenandmmutations are lethal relative to the wild-type genotypeNNMunless they cooccur, in which case they rescue each other. Under this model all females (and males) are eitherNNMornnm. Consider anMallele under this model. The chance that this allele encounters annallele during normal reproduction is approximatelyp, the frequency of thenallele, and the chance that it encounters it during MR is2p
(12p) +p2, which is 2por less. Now consider anmallele; the chance that it encounters anNallele is(12p) during normal reproduction and 2p(12p) + (12p)2during MR. Overall, the chance that there will be a lethal interaction during normal reproduction isfp+ (12f)(12p) and during MR it isf(2p(12p) +p2) + (12f)(2p(12p) + (12p)2), wherefis the frequency ofMallele. Logic dictates that the chance of a lethal interaction during MR must be less than twice the chance under normal sex, since both 2p(12p) +p2,2pand 2p(12p) + (12p)2,2(12p) (as above). However, over much of the parameter range (i.e.values ofpandf) the effect is attenuated, particularly when the mitochondrial variant segregates at a higher frequency than the nuclear variant.
In both of these models, we have assumed that the interaction is dominant and lethal; if the mutation is either partially dominant or recessive, or more weakly selected, then the LD between the nuclear and mitochondrial variants will be less strong and the difference in the frequency of DMNIs between MR and normal reproduction will be less, because the mother will have some chance of passing on the interacting alleles to her offspring. It is instructive, for example, to consider the case where the interaction is lethal but recessive; for example, themallele is only lethal on annnbackground (model III in Table A1). It is evident, in this case, that some females that carry themallele will also carry thenallele and hence pass on thenallele to her offspring. Consider anmallele and assume that both thenandmalleles are rare. The chance that themallele is in a female who is heterozygous for thenallele is2p(12p) and the chance that themallelefinds itself in an oocyte with thenallele is
p(12p), hence the probability of generating a zygote that isnnmisp2(12p). The probability that we form annnmgenotype given anmallele using MR is simplyp2. Hence, to afirst approximation, the probability of a DMNI during MR relative to normal sex is 1/(12p). Ifpis 1% the increase in the frequency of DMNIs due to MR would be1%.
Table A1 Thefitness of the six genotypes under several models of selection
Nuclear Genotype Mitochondrial Genotype Fitness Model I Fitness Model II Fitness Model III
NN M 1 1 1
Nn M 1 0 1
nn M 1 0 1
NN m 1 0 1
Nn m 0 0 1
GENETICS
Supporting Information
www.genetics.org/lookup/suppl/doi:10.1534/genetics.116.196436/-/DC1
Mitochondrial Replacement Therapy: Are Mito-nuclear
In-teractions Likely To Be a Problem?
Adam Eyre-Walker