0095-1137/96/$04.00
1
0
Copyright
q
1996, American Society for Microbiology
Molecular Typing of Yersinia pseudotuberculosis by Using an
IS200-Like Element
MONIQUE ODAERT, PATRICK BERCHE,
ANDMICHEL SIMONET*
Institut National de la Sante´ et de la Recherche Me´dicale U411, Faculte´ de Me´decine Necker,
75730 Paris Cedex 15, France
Received 18 March 1996/Returned for modification 1 May 1996/Accepted 12 June 1996
The IS200-like insertion sequence (IS) is a 708-bp element recently found in Yersinia pestis. Its nucleotide
sequence is 85% identical to that of IS200 recovered in most Salmonella enterica isolates. It is also present in
multiple copies in Y. pseudotuberculosis. In contrast, this IS is found in some (biotype 1B strains) but not other
Y. enterocolitica strains and is absent in the nonpathogenic yersiniae: Y. frederiksenii, Y. kristensenii, Y.
interme-dia, Y. bercovieri, and Y. mollaretii. The number and locations of the ISs in the Y. pseudotuberculosis genome vary
among strains, resulting in a high degree of polymorphism, but IS fingerprints are stable after multiple
subcultures of clinical isolates. The discriminative power of IS typing is better than that of ribotyping and
almost as good as that of the time-consuming method of pulsotyping. Overall, IS200-like is a useful molecular
marker in determining the epidemiology of Y. pseudotuberculosis infections.
The genus Yersinia includes three pathogenic species: Y.
pestis (the causative agent of bubonic plague), Y. enterocolitica,
and Y. pseudotuberculosis, the last two being responsible mainly
for enteritis and mesenteric lymphadenitis, which are
some-times complicated with septicemia. Seven environmental,
non-pathogenic species, Y. frederiksenii, Y. kristensenii, Y.
interme-dia, Y. bercovieri, Y. mollaretii, Y. aldovae, and Y. rohdei, have
been described (37). Y. pestis causes epidemic or pandemic
infections, while Y. pseudotuberculosis and Y. enterocolitica are
mostly involved in sporadic infections. However, several
out-breaks involving the two enteropathogenic species, Y.
entero-colitica (especially in North America) and Y. pseudotuberculosis
(mostly in Japan), have been reported (3, 15, 21, 27–29, 32–35).
In addition to serotyping, biotyping, and phage typing, a variety
of DNA-based methods have been developed for Yersinia
typ-ing, including analysis of restriction fragment polymorphism in
chromosomal DNA or in the plasmid pYV (1, 4, 10, 12, 13, 17,
22, 24), a 70-kb plasmid harbored by all pathogenic strains and
responsible for bacterial virulence (for a review, see reference
5).
We recently discovered a novel insertion sequence (IS) in Y.
pestis (31). This 708-bp IS, integrated within the inv gene from
Y. pestis, has 85% nucleotide identity with IS200, an element
which was first described in Salmonella enterica (14). This IS,
designated the IS200-like element, was found in multiple
cop-ies (at least 15 to 20) within the genomes of several Y. pestis
strains. Y. pseudotuberculosis DNA hybridizes with a probe
internal to the IS200-like element, but the insertion element
was present in fewer copies than in Y. pestis in the limited
number of strains tested (31). The degree of similarity between
the Y. pestis and Y. pseudotuberculosis IS200-like elements is
unknown, but restriction fragment length polymorphism
re-sults with the PCR products of the IS elements from both
species were identical (unpublished results).
ISs can be used as molecular tools for typing pathogenic
microorganisms, including Salmonella enterica, Staphylococcus
aureus, and Mycobacterium tuberculosis (2, 19, 20, 23, 36). This
conclusion appears to be paradoxical at first sight because ISs
are mobile genetic elements; however, transposition events
occur at a very low frequency. IS100 and IS285 have been
identified in pathogenic yersiniae (9, 25, 26), but these
inser-tion elements have not previously been used as molecular
markers in epidemiological analyses of plague and other forms
of yersiniosis. The aim of this work was to study the usefulness
of IS200-like elements for discriminating among strains
asso-ciated with known serotypes of enteropathogenic yersiniae. IS
fingerprinting was performed with 20 epidemiologically
unre-lated strains of Y. pseudotuberculosis, and the results allowed
good discrimination of the strains. In contrast, this molecular
typing method is not useful for epidemiological studies of Y.
enterocolitica infections.
MATERIALS AND METHODS
Bacterial strains and growth conditions.The Yersinia strains used in this study
are listed in Table 1. They were obtained from the Centre National de Re´fe´rence des Yersinia, Institut Pasteur (Paris, France). Plasmid pYV was detected in bacteria by colony-blot hybridization with a probe consisting of a 5.3-kb BamHI fragment from the Ca21dependence locus of virulence plasmid pIB1 as
previ-ously described (30). Bacteria were grown at 308C with aeration in Luria-Bertani broth or on agar.
Preparation of DNA probes.The IS200-like gene probe (421 bp) was obtained
by PCR amplification of pYV-cured Y. pestis 6/69Mc with the primers 59-TTCT TGTATCCTGGCCGT-39and 59-TGCGGTCTGGCAACT-39as previously de-scribed (31). The ribosomal 16S RNA (rrn) probe (about 1.5 kb) was obtained by PCR amplification of Y. pseudotuberculosis with the universal primer pair 59-G GTTACCTTGTTACGACTT-39and 59-AAGAGTTTGATCATGGCTCAG-39. Probes were labeled by random priming (Rediprime kit; Amersham France, Courtaboeuf, France) with [a-32P]dCTP (Amersham), and labeled DNA was
separated from unincorporated nucleotides by Sephadex G-50 column (Pharma-cia, Uppsala, Sweden) chromatography.
Southern hybridization.DNA extracted from bacterial cells as previously
described (18) was digested with appropriate restriction endonucleases according to the manufacturer’s instructions (New England Biolabs, Beverly, Mass.). Re-stricted DNA fragments were separated by electrophoresis through a 0.8% agarose gel in Tris-borate or Tris-acetate buffer and then transferred to a nylon membrane (Boehringer Gmbh, Mannheim, Germany). Probes were hybridized at 65 or 688C in buffer containing 0.1% sodium dodecyl sulfate (SDS), 63SSC (13SSC is 0.15 M NaCl plus 0.015 M sodium citrate [pH 7.0]), and 0.05% nonfat milk. After hybridization, filters were washed twice at room temperature for 30 min in 23SSC and 0.1% SDS and then twice at 65 or 688C for 45 min in 0.23
SSC and 0.1% SDS. Nylon membranes were autoradiographed by exposure to X-OMAT film (Eastman Kodak, Rochester, N.Y.) at2808C.
Pulsed-field gel electrophoresis.Bacterial DNA was embedded in agarose
plugs as previously described (7). DNA fragments generated by restriction en-donuclease digestion were separated in a 1% agarose–Tris-borate buffer gel by
* Corresponding author. Mailing address: Laboratoire de
Bac-te
´riologie-Hygie
`ne, Faculte
´ de Me
´decine Henri Warembourg, 1 Place
de Verdun, 59045 Lille, France. Phone: (33) 20444597. Fax: (33)
20529361. Electronic mail address: [email protected].
2231
on May 15, 2020 by guest
http://jcm.asm.org/
electrophoresis with a CHEF apparatus (Bio-Rad, Paris, France) at 148C and at 6 V/cm with alternating pulses at a 1208angle in a 1- to 30-s pulse-time gradient for 40 h. The gels were stained with ethidium bromide. DNAs from bacterio-phage lambda concatemers were used as size markers.
RESULTS AND DISCUSSION
The distribution of IS200-like elements in the genomes of
the pathogenic yersiniae Y. pseudotuberculosis and Y.
enteroco-litica and in the nonpathogenic species Y. frederiksenii, Y.
kris-tensenii, Y. intermedia, Y. bercovieri, and Y. mollaretii was
in-vestigated by Southern blot analysis. Total bacterial DNA was
digested with HincII, a restriction endonuclease that does not
cut within the IS element from either Y. pestis or Y.
pseudotu-berculosis, and DNA fragments were hybridized with a 421-bp
probe internal to the IS200-like element.
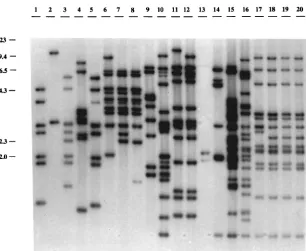
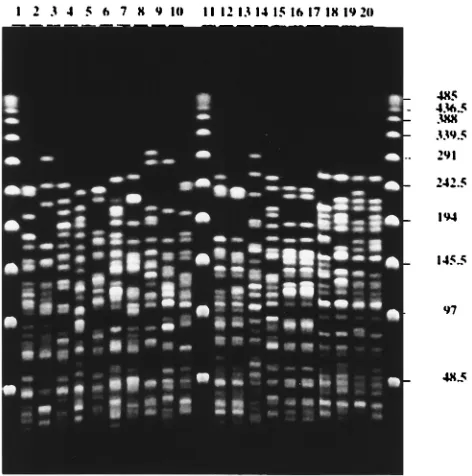
[image:2.612.55.557.82.293.2]Twenty strains of Y. pseudotuberculosis isolated from human
and animal infections in several different countries (Table 1)
were studied: 16 strains belonged to serotypes 1, 2, 3, 4, or 6,
and 4 were not typeable with specific antisera. All strains tested
contained the IS, and the number of copies and locations of the
FIG. 1. IS200-like element profiles of Y. pseudotuberculosis. Southern blot of HincII-digested DNA with an internal fragment of the IS200-like element labeled with [32P]dCTP. Lanes 1 to 5, serotype 1 strains; lanes 6 to 9, serotype 2 strains; lanes 10 to 12, serotype 3 strains; lanes 13 and 14, serotype 4 strains; lanes 15 and 16, serotype
[image:2.612.154.460.442.693.2]6 strains; lanes 17 to 20, untypeable strains. The positions and sizes (in kilobases) of HindIII fragments of bacteriophage lambda DNA are indicated on the left.
TABLE 1. Yersinia strains used in this study
Species Serotype Origin Country No. of strains
Y. pseudotuberculosis
1
Human, pig, and goat
France and Italy
5
2
Human and hare
France
4
3
Human and cow
Spain and Argentina
3
4
Human and sheep
United Kingdom
2
6
Guinea pig
Japan
2
Nontypeable
Human
France
4
Y. enterocolitica
American strains (biotype 1B)
8
Human
Norway and United States
2
13 and 18
Unknown
United States
1
13a and 13b
Unknown
United States
1
20
Unknown
United States
2
21
Unknown
United States
2
European strains (biotype non-1B)
3
Human
France and Belgium
4
9
Human
France and Belgium
3
Y. frederiksenii
Human
France
3
Y. kristensenii
Human and milk
France
3
Y. intermedia
Human and environment
France
3
Y. bercovieri
Human and environment
France
3
Y. mollaretii
Human and cow
France
3
on May 15, 2020 by guest
http://jcm.asm.org/
element in the genomes varied among strains, resulting in a
high degree of polymorphism (Fig. 1). Comparisons of the
IS200-like profiles of two parental strains harboring the
viru-lence plasmid pYV and their respective isogenic pYV-cured
derivatives indicated that most IS copies, if not all, were
in-serted into the bacterial chromosome (data not shown).
Eigh-teen different genomic DNA restriction patterns for the
IS200-like element were found among the 20 isolates. Some IS bands
were shared by several strains. Three of the four untypeable
strains (Fig. 1, lanes 18, 19, and 20) contained 13 copies of the
IS, with similar distributions occurring in the genomes. In the
fourth untypeable strain (Fig. 1, lane 17), there was one copy
in an
;
2.8-kb fragment instead of the 2.5-kb fragment
ob-served in strains 18, 19, and 20 (Fig. 1). By using restriction
endonuclease BamHI, EcoRI, or EcoRV, enzymes that do not
cut within the IS element, we were still unable to distinguish
the four untypeable strains (data not shown).
The stability of restriction patterns of Y. pseudotuberculosis
DNA for the IS200-like element was studied to assess the value
of this insertion element as a marker for typing. Five strains
containing 7 to 13 copies of the insertion element were
sub-cultured at 30
8
C on agar plates, and after 20 subcultures (i.e.,
about 600 generations), DNA of each strain was extracted and
digested with HincII and DNA fragments were hybridized with
the IS200-like probe. For all of the strains tested, we were
unable to detect any difference in IS patterns after these
mul-tiple passages (data not shown); the frequency of IS200-like
transposition is under investigation. Consequently, IS200-like
fingerprinting appears to be useful for typing Y.
pseudotuber-culosis.
We then compared the discriminative capacity of IS typing
with two other molecular typing methods, pulsotyping and
ribotyping. Y. pseudotuberculosis was typed by pulsed-field gel
electrophoresis of NotI-restricted DNA; NotI is a rare-cutting
enzyme (one site per 100 kb of Yersinia DNA) (16). This
method was previously reported to be very efficient for Y.
pseudotuberculosis typing (12). The resulting macrorestriction
patterns of the 20 Y. pseudotuberculosis strains each contained
about 20 fragments ranging in size from
;
40 to
;
300 kb (Fig.
2). The 20 strains could be classified into 19 pulsotypes
accord-ing to these electrophoretic patterns. Interestaccord-ingly, the four
untypeable strains that exhibited identical IS200-like profiles
displayed different macrorestriction patterns (Fig. 2, lanes 17,
18, 19, and 20). However, the two serotype 6 strains (Fig. 2,
lanes 15 and 16) had the same pulsotype but different IS types
(Fig. 1, lanes 15 and 16). Thus, the analytical performance of
IS typing was similar to that of pulsotyping, which is a
time-consuming method; however, the two methods were
comple-mentary since they allowed discrimination of all 20 strains used
in the study.
Finally, restriction fragment length polymorphism patterns
of the rDNA regions of Y. pseudotuberculosis were studied by
HindIII digestion and probing with the 16S rRNA probe. This
restriction endonuclease does not cut within the 1.5-kb probe
and was previously shown by Picard-Pasquier et al. (24) to
discriminate Y. pseudotuberculosis strains well (five ribotypes
were individualized among nine unrelated strains). Bands
con-taining the rrn gene were 7 to
.
12 kb (Fig. 3), and 14 ribotypes
were obtained among the 20 strains. One strain of serotype 1
(Fig. 3, lane 4) and one of serotype 2 (lane 7) exhibited the
[image:3.612.60.296.70.308.2]FIG. 2. NotI restriction profiles of Y. pseudotuberculosis. DNA was digested with NotI, and fragments were separated by pulsed-field gel electrophoresis as described in Materials and Methods. Lanes 1 to 5, serotype 1 strains; lanes 6 to 9, serotype 2 strains; lanes 10 to 12, serotype 3 strains; lanes 13 and 14, serotype 4 strains; lanes 15 and 16, serotype 6 strains; lanes 17 to 20, untypeable strains. The positions and sizes (in kilobases) of bacteriophage lambda DNA concate-mers are indicated on the right.
FIG. 3. 16S rRNA profiles of Y. pseudotuberculosis. Southern blot of HindIII-digested DNA with a 16S rRNA probe labeled with [32P]dCTP. Lanes 1 to 5, serotype
1 strains; lanes 6 to 9, serotype 2 strains; lanes 10 to 12, serotype 3 strains; lanes 13 and 14, serotype 4 strains; lanes 15 and 16, serotype 6 strains; lanes 17 to 20, untypeable strains. Size markers (1-kb DNA ladder) are indicated on the left.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:3.612.119.495.558.697.2]same 16S rRNA profile, and one strain of serotype 2 (lane 6)
and two strains of serotype 3 (lanes 11 and 12) similarly fell
into the same riboclass. Moreover, the two serotype 6 strains,
which were not discriminated by pulsotyping, displayed the
same rRNA pattern (lanes 15 and 16). Finally, the four
un-typeable strains (lanes 17 to 20) were separated into two
groups (lanes 17 and 20 and lanes 18 and 19) on the basis of
their 16S rRNA pattern. Consequently, the discriminatory
power of IS typing is better than that of ribotyping. In addition,
analysis of rRNA pattern is not easy because of the fact that
fragments containing the rrn genes were mostly of large size
and thus poorly separated by agarose gel electrophoresis.
We also looked for the presence of the IS in the other
enteropathogenic Yersinia sp., Y. enterocolitica. Several
bio-types of this species have been defined by Wauters, Kandolo,
and Janssens (38). Biotype 1B strains, mainly from North
America, are highly pathogenic and responsible for major
out-breaks (especially of serotype O:8), whereas strains belonging
to other biotypes are less pathogenic or not pathogenic. We
studied 15 strains, of which 8 were from biotype 1B (American
strains). Unlike for Y. pseudotuberculosis, the IS200-like probe
hybridized with only some of the Y. enterocolitica strains (Fig.
4): only four biotype 1B strains (serotypes O:8, O:13,18, and
O:13a,13b) gave strong hybridization signals (one or two
bands). However, several faint background bands were
de-tected for most of the American strains. These faint bands
were still detected when DNA-DNA hybridization was carried
out under higher stringency conditions (68
8
C); thus, the weak
intensities of most of the bands suggest the presence of another
repetitive element in biotype 1B strains, similar but not
iden-tical to the IS200-like element; cloning and sequencing of this
element from these strains is in progress. None of the seven
non-biotype 1B strains (four of serotype O:3 and three of
serotype O:9) hybridized with the IS200-like probe. These data
are an additional argument indicating that the two groups of Y.
enterocolitica correspond to two divergent clusters, as
previ-ously suggested by Caugant et al. (6). Production of
iron-repressible proteins and the siderophore yersiniabactin under
iron starvation conditions are characteristic traits of the highly
pathogenic biotype 1B strains (8, 11), and the IS200-like
ele-ment could be an additional marker of Y. enterocolitica
patho-genicity. This proposition is reinforced by the absence of this
element in the nonpathogenic Yersinia species Y. frederiksenii,
Y. kristensenii, Y. intermedia, Y. bercovieri, and Y. mollaretii
(three strains were tested for each species [data not shown]).
ACKNOWLEDGMENTS
We thank E. Carniel, V. Escuyer, and P. Trieu-Cuot for critical
readings of the manuscript, and E. Abachin for technical assistance.
This work was supported by INSERM and the Universite
´ Rene
´
Descartes (Paris V, France).
REFERENCES
1. Andersen, J. K., and N. A. Saunders. 1990. Epidemiological typing of
Yer-sinia enterocolitica by analysis of restriction fragment length polymorphisms
with a cloned ribosomal RNA gene. J. Med. Microbiol. 32:179–187. 2. Baquar, N., A. Burnens, and J. Stanley. 1994. Comparative evaluation of
molecular typing of strains from a national epidemic due to Salmonella
brandenburg by rRNA gene and IS200 probes and pulsed-field gel
electro-phoresis. J. Clin. Microbiol. 32:1876–1880.
3. Black, R. E., R. J. Jackson, T. Tsai, M. Medvesky, M. Shayegani, J. C. Feeley,
K. I. E. McLeod, and A. M. Wakelee.1978. Epidemic Yersinia enterocolitica
infection due to contaminated chocolate milk. N. Engl. J. Med. 298:76–79. 4. Blumberg, H. M., J. A. Kiehlbauch, and I. K. Wachsmuth. 1991. Molecular epidemiology of Yersinia enterocolitica O:3 infections: use of chromosomal DNA restriction fragment length polymorphisms of rRNA genes. J. Clin. Microbiol. 29:2368–2374.
5. Brubaker, R. R. 1991. Factors promoting acute and chronic diseases caused by yersiniae. Clin. Microbiol. Rev. 4:309–324.
6. Caugant, D. A., S. Aleksic, H. H. Mollaret, R. K. Selander, and G. Kapperud. 1989. Clonal diversity and relationships among strains of Yersinia
enteroco-litica. J. Clin. Microbiol. 27:2678–2683.
7. Che´ron, M., E. Abachin, E. Gue´rot, M. El-Bez, and M. Simonet. 1994. Investigation of hospital-acquired infections due to Alcaligenes denitrificans subsp. xylosoxydans by DNA restriction fragment length polymorphism. J. Clin. Microbiol. 32:1023–1026.
8. de Almeida, A. M. P., A. Guiyoule, I. Guilvout, I. Iteman, G. Baranton, and
E. Carniel.1993. Chromosomal irp2 gene in Yersinia: distribution,
expres-sion, deletion and impact on virulence. Microb. Pathog. 14:9–21. 9. Filippov, A. A., P. N. Oleinikov, V. L. Motin, O. A. Protsenko, and G. B.
Smirnov.1995. Sequencing of two Yersinia pestis IS elements, IS285 and
IS100. Contrib. Microbiol. Immunol. 13:306–309.
10. Fukushima, H., M. Gomyoda, S. Aleksic, and M. Tsubokura. 1993. Differ-entiation of Yersinia enterocolitica serotype O:5, 27 strains by phenotypic and molecular techniques. J. Clin. Microbiol. 31:1672–1674.
11. Heesemann, J. 1987. Chromosomal-encoded siderophores are required for mouse virulence of enteropathogenic Yersinia species. FEMS Microbiol. Lett. 48:229–233.
12. Iteman, I., H. Najdenski, and E. Carniel. 1995. High genomic polymorphism in Yersinia pseudotuberculosis. Contrib. Microbiol. Immunol. 13:106–111. 13. Kapperud, G., T. Nesbakken, and K. Dommarsnes. 1991. Application of
restriction endonuclease analysis and genetic probes in the epidemiology of
Yersinia enterocolitica infection. Contrib. Microbiol. Immunol. 12:68–74.
14. Lam, S., and J. R. Roth. 1983. IS200: a Salmonella-specific insertion se-quence. Cell 34:951–960.
15. Lee, L. A., A. R. Gerber, D. R. Lonsway, J. D. Smith, G. P. Carter, N. D.
Puhr, C. M. Parrish, R. K. Sikes, R. J. Finton, and R. V. Tauxe.1990.
Yersinia enterocolitica O:3 infections in infants and children, associated with
the household preparation of chitterlings. N. Engl. J. Med. 322:984–987. 16. Lucier, T. S., and R. R. Brubaker. 1992. Determination of genome size,
macrorestriction pattern polymorphism, and nonpigmentation-specific dele-tion in Yersinia pestis by pulsed-field gel electrophoresis. J. Bacteriol. 174: 2078–2086.
17. Makino, S.-I., Y. Okada, T. Maruyama, S. Kaneko, and C. Sasakawa. 1994. PCR-based random amplified polymorphic DNA fingerprinting of Yersinia
pseudotuberculosis and its practical applications. J. Clin. Microbiol. 32:65–69.
18. Marmur, J. 1961. A procedure for the isolation of deoxyribonucleic acid from microorganisms. J. Mol. Biol. 3:208–218.
19. Millemann, Y., M.-C. Lesage, E. Chaslus-Dancla, and J.-P. Lafont. 1995. Value of plasmid profiling, ribotyping, and detection of IS200 for tracing avian isolates of Salmonella typhimurium and S. enteritidis. J. Clin. Microbiol.
33:173–179.
[image:4.612.59.295.71.298.2]20. Monzon-Moreno, C., S. Aubert, A. Morvan, and N. El Sohl. 1991. Usefulness of three probes in typing isolates of methicillin-resistant Staphylococcus FIG. 4. IS200-like profiles of Y. enterocolitica. Southern blot of
HincII-di-gested DNA with an internal fragment of the IS200-like element labeled with [32
P]dCTP. Lanes 1 to 8, biotype 1B strains; lanes 9 to 15, biotype non-1B strains. Size markers (1-kb DNA ladder) are indicated on the left.
on May 15, 2020 by guest
http://jcm.asm.org/
aureus (MRSA). J. Med. Microbiol. 35:80–88.
21. Morse, D. L., M. Shayegani, and R. J. Gallo. 1984. Epidemiologic investi-gation of a Yersinia camp outbreak linked to a food handler. Am. J. Public Health 74:589–592.
22. Najdenski, H., I. Iteman, and E. Carniel. 1994. Efficient subtyping of patho-genic Yersinia enterocolitica strains by pulsed-field gel electrophoresis. J. Clin. Microbiol. 32:2913–2920.
23. Pelkonen, S., E.-L. Romppanen, A. Siitonen, and J. Pelkonen. 1994. Differ-entiation of Salmonella serovar infantis isolates from human and animal sources by fingerprinting IS200 and 16S rrn loci. J. Clin. Microbiol. 32:2128– 2133.
24. Picard-Pasquier, N., B. Picard, S. Heeralal, R. Krishnamoorthy, and P.
Goullet.1990. Correlation between ribosomal DNA polymorphism and
elec-trophoretic enzyme polymorphism in Yersinia. J. Gen. Microbiol. 136:1655– 1666.
25. Podladchikova, O. N., G. G. Dikhanov, A. V. Rakin, and J. Heesemann. 1994. Nucleotide sequence and structural organization of Yersinia pestis insertion sequence IS100. FEMS Microbiol. Lett. 121:269–274.
26. Portnoy, D. A., and S. Falkow. 1981. Virulence-associated plasmids from
Yersinia enterocolitica and Yersinia pestis. J. Bacteriol. 148:877–883.
27. Randall, K. J., and N. S. Mair. 1962. Family outbreak of Pasteurella
pseudo-tuberculosis infection. Lancet i:1042–1043.
28. Sanbe, K., M. Uchimura, K. Koiwai, K. Takagi, H. Yazaki, Y. Nanayama,
and M. Ohtawara.1987. Community outbreak of Yersinia pseudotuberculosis
occurred among primary school children. J. Jpn. Assoc. Infect. Dis. 61:763– 771.
29. Sato, K., K. Ouchi, and M. Taki. 1983. Yersinia pseudotuberculosis infection in children, resembling Izumi fever and Kawasaki syndrome. Pediatr. Infect. Dis. 2:123–126.
30. Simonet, M., and S. Falkow. 1992. Invasin expression in Yersinia
pseudotu-berculosis. Infect. Immun. 60:4414–4417.
31. Simonet, M., B. Riot, N. Fortineau, and P. Berche. 1996. Invasin production by Yersinia pestis is abolished by insertion of an IS200-like element within the
inv gene. Infect. Immun. 64:375–379.
32. Tacket, C. O., J. Ballard, N. Harris, J. Allard, C. Nolan, T. Quan, and M. L.
Cohen.1985. An outbreak of Yersinia enterocolitica infections caused by
contaminated tofu (soybean curd). Am. J. Epidemiol. 121:705–711. 33. Tacket, C. O., J. P. Narain, R. Sattin, J. P. Lofgren, C. Konigsberg, Jr., R. C.
Rendtorff, A. Rausa, B. R. Davis, and M. L. Cohen.1984. A multistate
outbreak of infections caused by Yersinia enterocolitica transmitted by pas-teurized milk. JAMA 251:483–486.
34. Tertti, R., K. Granfors, O.-P. Lehtonen, J. Mertsola, A.-L. Ma¨kela¨, I. Va
¨li-ma¨ki, P. Ha¨nninen, and A. Toivanen.1984. An outbreak of Yersinia
pseudo-tuberculosis infection. J. Infect. Dis. 149:245–250.
35. Toyokawa, Y., Y. Ohtomo, and T. Akiyama. 1993. Large scale outbreak of
Yersinia pseudotuberculosis serotype 5a infection at Noheji-machi in Aomori
Prefecture. J. Jpn. Assoc. Infect. Dis. 67:36–43.
36. Van Soolingen, D., P. W. M. Hermans, P. E. W. de Haas, D. R. Soll, and
J. D. A. van Embden.1991. Occurrence and stability of insertion sequences
in Mycobacterium tuberculosis complex strains: evaluation of an insertion sequence-dependent DNA polymorphism as a tool in the epidemiology of tuberculosis. J. Clin. Microbiol. 29:2578–2586.
37. Wauters, G., M. Janssens, A. G. Steigerwalt, and D. J. Brenner. 1988.
Yersinia mollaretii sp. nov. and Yersinia bercovieri sp. nov., formerly called Yersinia enterocolitica biogroups 3A and 3B. Int. J. Syst. Bacteriol. 38:424–
429.
38. Wauters, G., K. Kandolo, and M. Janssens. 1987. Revised biogrouping scheme of Yersinia enterocolitica. Contrib. Microbiol. Immunol. 9:14–21.
![FIG. 4. IS200gested DNA with an internal fragment of the IS[32P]dCTP. Lanes 1 to 8, biotype 1B strains; lanes 9 to 15, biotype non-1B strains.-like profiles of Y](https://thumb-us.123doks.com/thumbv2/123dok_us/8218466.820964/4.612.59.295.71.298/gested-internal-fragment-biotype-strains-biotype-strains-proles.webp)
