20
Engineering Gene
Circuits: Foundations
and Applications
Dennis Tu Jiwon Lee Taylan Ozdere Tae Jun Lee Lingchong YouDuke University, Durham
20.1 Introduction...20-363 20.2 Designing Gene Circuits ...20-364
Modeling-Guided Rational Design.
Refining a Circuit through Evolutionary Techniques
20.3 Programming Robust Circuit Behavior...20-366
Cellular Noise.Basic Regulatory
Motifs to Resist Noise.Cell–Cell Communication
20.4 Applications ...20-370
Gene Therapy.Drug Development.
Biotechnology Applications
20.5 Outlook ...20-372
Abstract
Synthetic biology has emerged as a useful approach to decoding fundamental laws underlying biological control. Recent efforts have produced many exciting systems and generated substantial insights. These progresses highlight the potential of synthetic biology to impact diverse areas including biology, computation, engineering, and medicine.
20.1 Introduction
Biological systems often function reliably in diverse environments despite internal or external perturba-tions. This behavior is often characterized as ‘‘robustness.’’ Based on extensive studies over the last several decades, much of this robustness can be attributed to the control of gene expression through complex cellular networks [1–4]. These networks are known to consist of various regulatory modules, including feedback [5] and feed-forward [6] regulation and cell–cell communication [7]. With these basic regulatory modules and motifs, researchers are now constructing artificial networks that mimic nature to gain fundamental biological insight and understanding [8]. In addition, other artificial net-works that are engineered with novel functions will serve as building blocks for future practical appli-cations. These efforts form the foundation of the recent emergence of synthetic biology [3,9,10]. These artificial networks are interchangeably called ‘‘synthetic gene circuits’’ or ‘‘engineered gene circuits.’’ Recent accomplishments in synthetic biology include engineered switches [11–14], oscillators [15,16], logic gates [17–19], metabolic control [20], reengineered translational machinery [21], population control [22] and pattern formation [23] using natural or synthetic [24] cell–cell communication, reengineered viral genome [25], and hierarchically complex circuits built upon smaller, well-characterized 20-363
functional modules [26] In addition to programming cellular dynamics, efforts have also been made toward the construction of in vitro gene circuits using cell extracts [27,28].
As evidenced in the term ‘‘circuit,’’ gene circuits are often compared to electrical circuits. For example, one can characterize the state of a gene as ON or OFF depending on whether it is expressed or repressed. Construction of electrical circuits benefits from a large collection of well-characterized parts and modules, including resistors, capacitors, and inductors, which can be connected to generate a complex circuit with a useful function. The rapid progress in high-throughput technologies is now generating similar parts-lists, as evidenced by the establishment of numerous databases [29–32]. In particular, efforts along this line have led to the creation of a database of biological parts (http:==parts.mit.edu=) — the Registry of Standard Biological Parts—that is specifically established to facilitate gene-circuit design and implementation. This registry contains details on basic parts (mostly for bacteria) such as ribosome binding sites, transcriptional terminators, and reporter genes. As of this writing, the registry contains details on over 1500 parts. The ultimate goal is to be able to order standardized parts ‘‘off the shelf ’’ from the registry and assemble them together to engineer a functional circuit. The current registry documents mostly components from bacteria, especially E. coli. We anticipate that this or similar registries will also be expanded to include yeast, eukaryotes, and other organisms. In addition, we note that the recent development of high throughput, low-cost DNA synthesis technology [33] provides a unique opportunity to explore parts not found in nature.
20.2 Designing Gene Circuits
20.2.1 Modeling-Guided Rational Design
The first term that comes to mind when considering the creation of a circuit is ‘‘engineering’’ or ‘‘programming.’’ With this mindset, many novel synthetic gene circuits have been created through iterations of careful design, modeling, and implementation. A well-known synthetic gene circuit is the genetic toggle switch, whose name is derived from its ability to program bistable behavior (Figure 20.1a) [12]. The toggle switch circuit consists of two repressors, TetR and LacI, which mutually repress each other (Figure 20.1a). The circuit can be flipped between ‘‘high’’ and ‘‘low’’ states using chemical inducers isopropyl-b-D-thiogalactopyranoside (IPTG) and anhydrotetracycline (aTc). For example, when aTc is added to the system to inhibit tetR, lacI will be highly expressed and the reporter will be OFF. Conversely, when IPTG is added to inhibit lacI, tetR and the reporter will be highly expressed.
(a) (b) IPTG Lacl P trc-2 aTc TetR Reporter Lacl Lacl TetR PLtetO-1 Ptet Plac TetR λcl λP λcl
FIGURE 20.1 (a) The genetic toggle switch gene circuit layout. TetR represses PLtetO-1and is inhibited by inducer
aTc; LacI represses Ptrc-2 and is inhibited by inducer IPTG. (b) Repressilator gene circuit layout. TetR represses expression oflcI which in turn represses expression of LacI. Finally, LacI represses TetR expression, completing the repressilator cycle.
Mathematical modeling suggested that the circuit could demonstrate bistability for appropriate circuit parameter values. That is, for particular parameter sets, the system can reach either the stable high state or the stable low state, depending on the prior history of its dynamics. For example, if a sufficiently strong pulse of IPTG is applied to the circuit (previously at the low state), the circuit will be turned ON. Importantly, it will stay ON even if the IPTG is reduced to a low level or is even completely absent. Modeling also predicted the conditions where bistability would be favored, which included higher cooperativity in transcription repression and faster synthesis rates of the repressors. Guided by these modeling predictions, the authors used commonly available, well-characterized elements (e.g., tet and lac repressors) to successfully implement a library of functional bistable toggles.
This relatively simple circuit demonstrates a dynamic property with broad implications in biology [34]. For example, bistable behavior can ensure that a gene is fully ON or OFF without intermediary states, which can serve as the basis for cell fate decision [35] and cell differentiation [36]. Because an induction with sufficient strength and duration is required for switching, a bistable circuit may act as a noise filter, which is discussed in detail later.
Another well-known circuit is the repressilator [15], which consists of three repressors sequentially repressing each other. As shown in Figure 20.1b, the first repressor protein (TetR) inhibits the second repressor gene whose protein product (lcI) in turn inhibits the third repressor gene (lacI). Finally, the third repressor protein (LacI) inhibits the first repressor gene, completing a negative feedback loop with a long cascade.
Similar to the design process of the toggle switch [12], mathematical modeling was used to analyze the circuit design and guide its implementation. Modeling suggested that the circuit could generate sustained oscillations with concentrations of the three repressors within biologically feasible ranges. Through detailed analysis, it was found that oscillations would be favored by strong promoters, tight transcriptional repression (low ‘‘leakiness’’), cooperative repression characteristics, and comparable protein and mRNA decay rates. With this knowledge, the stability of each of the repressor proteins was modulated by using degradation tags [37]. This careful engineering eventually resulted in a circuit that generated oscillatory behavior in individual cells.
These pioneering studies illustrate that we have acquired enough biological understanding to begin programming cellular dynamics in a rational way. Nevertheless, there are many challenges ahead. For example, compared with natural biological oscillators, such as circadian oscillators [38–40], the repres-silator is unreliable in that oscillations only occur in a subpopulation of cells and these oscillations are often noisy.
20.2.2 Refining a Circuit through Evolutionary Techniques
Despite careful design and computer simulation, building a gene circuit in vivo may still be challenging due to a lack of detailed understanding of the cellular components and how they interact with one another. The result of this knowledge gap is a circuit that may require fine tuning of circuit components or even rearranging the entire circuit topology. This process may benefit from evolutionary techniques. For example, in silico evolution is a technique using computer simulated evolution to discover the best parameter sets. This technique entails simulating a library of networks with varying parameter settings and topologies in a computer. Then, these networks are screened for the best performing ones according to a predefined objective function [41] For instance, one could select circuits that generate oscillations with a certain period. Winners from the first round of screening are ‘‘mutated’’ further by varying their parameters or topologies and computationally screened again for variants that display improved performance. Several rounds of in silico mutagenesis and screening often leads to circuits with topologies and parameter settings that ensure greatly improved performance according to a predefined objective. The optimal parameters discovered through this computer simulated evolution are useful in guiding the circuit designer to select appropriate promoters and genes. An advantage of this strategy is that it can potentially identify alternative designs that are not obvious to a ‘‘rational designer.’’ These could include circuits with the same architecture but with nonobvious parameter settings, or circuits
with completely different network architecture and different parameter settings. However, the extent to which this strategy can speed up circuit design and implementation is unclear. In fact, synthetic gene circuits are yet to be experimentally implemented based on a design resulting from this approach.
Laboratory evolution can be realized by combining random mutagenesis and subsequent high-throughput screening or selection for a desired function [42,43]. This ‘‘directed evolution’’ technique has been used successfully for years to optimize enzymes by improving or altering their efficiency and specificity [44]. In gene circuit design, directed evolution has been used to optimize metabolic pathways to produce useful metabolites [45]. This optimization is accomplished by generating components such as proteins and DNA regulatory elements with diverse kinetic properties that may not be naturally available. In other words, it allows researchers to take advantage of a broader parameter space when designing a gene circuit. In recent work, an evolved transcriptional activator (LuxR) [46] was used in a synthetic pattern formation circuit [23] to optimize the final circuit function.
Another application of directed evolution is to optimize a partially functional or nonfunctional circuit to be fully functional [47]. For example, this technique was used to evolve an initially nonfunctional genetic inverter into a fully functional one [19]. Briefly, several rounds of mutagenesis by error-prone PCR and subsequent high-throughput screening were able to achieve proper matching of the different components that were assembled to make up the inverter, allowing for rapid optimization of the circuit [19]. It is interesting to note that the functional circuit, created from directed evolution, ended up with parameters tuned completely different from what was attempted by a rational approach [17]. This aspect again highlights the advantage of evolution methods to explore a parameter space not readily accessible by rational design.
More recently, this technique was further revised by combining mutagenesis with selection instead of screening [48]. That is, the ability of a circuit to function properly is tied to the survival or death of the host cell. This selection strategy, if properly established, can drastically improve the throughput of circuit evolution. In this proof-of-concept work, Yokobayashi and Arnold were able to isolate a functional inverter from a pool with more than 3 million fold background of nonfunctional circuits. Although this work demonstrated a technique that was limited to optimization of a circuit with two output states (e.g., logic gates and toggle switches), more complex circuits may be broken down into such submodules for individual optimization.
Random mutagenesis is unlikely to alter the connectivity of a circuit; rather, it largely results in changes in one or more circuit parameters. Therefore, for directed evolution techniques to succeed, they require a starting circuit with a topology that can in principle display the desired function with proper parameter settings. Complementary to random mutagenesis, a combinatorial method will enable the generation of a large number of circuits with diverse topologies from a small number of elements. As a proof-of-concept, Guet and colleagues created 125 logic circuits using only three genes and five promoters [18]. The resulting circuits displayed diverse phenotypes including NOR, NOT IF, and NAND. Potentially, the same strategy can be applied to a larger number of elements and an even greater diversity of circuit connectivity can be created. When coupled with efficient screening or selection methods, this strategy can allow evolution of a circuit in both the topological space and the parametric space.
20.3 Programming Robust Circuit Behavior
20.3.1 Cellular Noise
Another fundamental challenge in constructing gene circuits with reliable function is to deal with cellular ‘‘noise’’ or stochastic fluctuations in cellular processes. Such noise can be due to a number of factors, including small numbers of interacting molecules, heterogeneity of the cellular environment, perturbations from the extracellular environment, and extended gene cascades [49–58].
Noise may benefit cells by providing phenotypic variations through which they adapt to changing environments [59–62]. In most cases, however, noise is detrimental because it poses a significant
challenge for creating circuits with robust function. For example, the repressilator displays noisy behavior. Only 40% of the cells in the population exhibited oscillatory behavior and these oscillations are often out of phase with each other, even between sibling cells [15]. To a lesser degree, cellular noise also cause variations in the oscillatory dynamics in a more sophisticated oscillator (metabolator) recently implemented [16]. In contrast, natural oscillators function much more reliably. For example, single-cell measurements in cyanobacteria revealed highly robust oscillations for days [63]. An intri-guing question is how natural systems are able to function with such robustness despite cellular noise.
20.3.2 Basic Regulatory Motifs to Resist Noise
Extensive theoretical and experimental studies have been conducted to elucidate the underlying mech-anisms by which biological systems achieve robustness [50,64–66]. One such mechanism is negative feedback, where the output from a system reduces its own output. In a biological context, this occurs when a protein limits its own accumulation by repressing its own synthesis [14,67] or by facilitating its own turnover [68,69].
A well-studied example is bacterial chemotaxis, a phenomenon where bacteria move toward an attractant by biasing their swimming motion [2]. This response is controlled by receptors that detect the attractant gradient. When the bacterium reaches a steady concentration of attractant, adaptation occurs. This ‘‘zeroes’’ the detector so that it can react to new gradients. This adaptation was found to be resistant to internal and external perturbations [1,2]. In-depth theoretical analysis indicates that the robust behavior of the circuit can be attributed to an integral negative feedback [70].
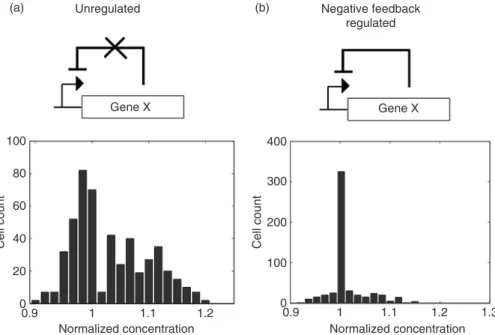
Over three decades ago, Savageau pioneered systems-level analysis of various regulatory motifs [71], and found that negative feedback can confer stability in system output and improve system response speed [67]. These predictions were recently verified experimentally. Besckei and Serrano constructed a simple synthetic circuit where transcription repressor TetR represses transcription of its own gene (Figure 20.2). Comparison of this autoregulated gene circuit with an unregulated counterpart by negative feedback indicated that the autoregulation indeed reduced noise, as measured in the distribu-tion of gene expression across the populadistribu-tion (Figure 20.2) [5]. That is, the autoregulated circuit
(a) Unregulated (b) Negative feedback
regulated
Gene X Gene X
Normalized concentration Normalized concentration Cell count Cell count
0 20 40 60 80 100 0 100 200 300 400 0.9 1 1.1 1.2 0.9 1 1.1 1.2 1.3
FIGURE 20.2 (a) An unregulated circuit as compared with (b) an autoregulated circuit by negative feedback results in a narrower distribution of gene expression as compared with an unregulated circuit with no feedback.
resulted in gene expression with a much narrower distribution compared to the unregulated circuit. To further illustrate the importance of negative feedback, the autoregulatory loop was gradually removed by adding an inducer. As the inducer concentration increased, the negative feedback circuit behavior transitioned to that of the unregulated circuit, resulting in a wide distribution of gene expression [5]. Part of the mechanism behind this behavior may be an increase in the noise frequency. This frequency shift is thought to enable the cell to more effectively filter the noise [72]. Using the same circuit, Alon and colleagues demonstrated that negative feedback also significantly improved system response speed [73].
An alternative approach to resisting noise and enhancing stability is increasing the rise-time in gene expression. Rise-time is the time it takes for a gene circuit to reach a steady-state level of protein expression [73]. An increased rise-time requires a longer-lasting input to fully induce a gene. This behavior can be implemented through a coherent feed-forward loop, where transcription factor A regulates B, and both jointly control the expression of C [6]. The latter stage of the feed-forward loop causes an increase in rise-time, allowing the cell to filter noise from the input by withstanding transient perturbations.
Another regulatory feature used to increase cellular robustness is positive feedback. Positive feedback occurs when the output of a network causes an increase in the same output. It can be realized by positive autoregulation [13,14], mutual activation [36,74,75] or mutual inhibition [12] (as detailed in Figure 20.1a). Positive feedback loops often amplify noise [13,64]. On the other hand, when these modules display bistable switching behavior with appropriate parameter sets, they may serve as a noise-resistant device. This is because a sufficiently strong, long-lasting input signal is required to change the state of the switch [76]. Using this feature as a noise filter to stabilize oscillations was proposed on the basis of computational analysis [77]. Subsequent experimental efforts successfully generated population-level oscillations, although these were damped oscillations [78]. In another example of noise-resistant behavior, cellular ‘‘memory’’ is controlled by a positive feedback loop. This memory allows the cell to maintain a state that depends on the initial input. Once the memory is established, the state of the cell does not change with varying input as long as the memory is sustained by the positive feedback loop [79].
20.3.3 Cell–Cell Communication
In recent years, scientists have discovered that bacteria do not exist merely as individuals, but can coordinate their behavior as a group through quorum sensing [7,80,81]. Quorum sensing is a technique by which bacteria use cell–cell communication to sense changes in cell population density. This ability has been extensively studied for its role in regulating diverse physiological functions such as biolumin-escence [80], virulence [82,83], biofilm formation [84], and programmed death [85–87].
In quorum sensing, signaling molecules are expressed by the cells and can freely diffuse through the cell membrane. At a low population density, these molecules diffuse out of the cells into the extracellular space with little accumulation of the signal within the cells. Thus, communication is weak and the cells act independent of one another. As the population increases, the signaling molecule concentration increases both inside and outside the cells. Upon reaching a threshold concentration, the signal will lead to coordinated expression of genes responsible for various functions [7,81].
Intuitively, the large number of signaling molecules outside the cells can act as a buffer and temper the fluctuations of the signaling molecule in each cell. That is, the fluctuation in the concentration of the signaling molecule is likely much smaller compared with molecules of the same concentration, but confined in individual cells. Consider a molecule with a concentration of 10 nM. If it is confined in the cell, there will be about 10 molecules=cell (assuming a cell volume of 10 15L). The fluctuation in its concentration will be about 101=2, or a relative fluctuation of 0.3. However, for a freely diffusible signaling molecule with the same concentration in a 1 mL culture, the molecular number is 1013, and the relative fluctuation is around 10 6.5, which is considerably less noisy. Therefore, if a gene is under control of such a signaling molecule, there will likely be less noise in its expression across the population.
Confirming this point, our simulations suggest that cell–cell communication can indeed reduce noise in gene expression across a population of cells. The distribution of gene expression controlled by quorum sensing is much tighter than the distribution found in the system without cell–cell communication (unpublished data). Our simulations also suggest that this effect depends only on the cell density and diffusion rate and not on the number of cells. With this evidence, quorum sensing may be a side effect of batch culture cells detecting their environmental conditions through diffusion [88].
It has been proposed that cell–cell communication may be used to facilitate more robust oscillations. Recent computer simulations have demonstrated that the repressilator oscillations can be synchronized across an entire population of cells through cell–cell communication [89,90]. The same strategy was also proposed to synchronize relaxation-type oscillators [91,92]. Although this synchronization strategy remains to be demonstrated experimentally, there is evidence that suggests that interlocked oscillators provide more robust behavior in nature [93].
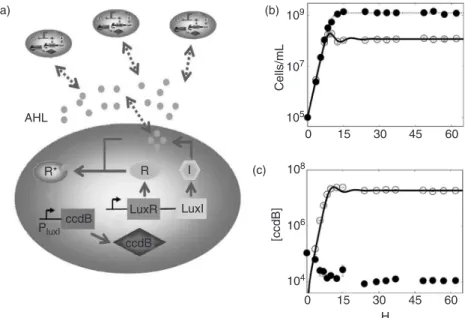
In addition to synchronizing oscillators, cell–cell communication can serve as an effective mechanism to achieve integrated and coordinated behavior among a population of cells. In a synthetic ‘‘population control circuit,’’ quorum sensing was coupled with a negative-feedback loop to regulate the cell density by controlling the death rate [22]. As the size of the population increases, the signaling molecule (AHL) builds up in the environment and inside the cells. When a particular population density is reached, the production of a killer gene (ccdB) is activated and the death rate increases. Gradually, the population density steady state is achieved when the growth rate equals the death rate (Figure 20.3). Simple mathematical models predict that the circuit can generate stable steady states [22] or sustained oscillations [94]. 0 15 30 45 60 0 15 30 45 60 Cells/mL 109 107 105 104 106 108 H [ccdB] AHL (a) (b) (c) R I LuxI LuxR ccdB ccdB R* PluxI
FIGURE 20.3 A population control circuit that detects the size of a population and activates a cell killing gene when the cell density is too high (a) Acyl-homoserine lactone (AHL) is produced by LuxI (I) and freely diffuses through the cell. At high enough concentration, AHL binds to and activates LuxR (R). Activated LuxR (R*) binds to the PluxIpromoter causing expression of the killer gene (ccdB) and cell death. Both luxRandluxIare the under
control of an inducible promoter Placpromoter. (b) Experimentally measured growth curves with the circuit ON
(open circles) and OFF (filled circles). The model predictions are in solid (ON) and dotted (OFF) lines. (c) A time course showing the concentration of killer gene (ccdB) produced in the ON and OFF cultures. This was measured by assaying the LacZ activity because theccdBwas fused tolacZa. There is noccdBconcentration line for model prediction of the OFF culture because it is always zero. (From You, L., Cox, R.S. III, Weiss, R., and Arnold, F.H., Nature428, 868, 2004.)
Experimentally, this population control circuit can be turned ON by inducing the quorum sensing module. As expected, the uninduced (OFF) culture grew exponentially until reaching a stationary phase at the maximum density the culture medium could sustain (Figure 20.3b). The ON culture initially grew similarly as the OFF culture until a threshold was reached at 7 h after circuit activation. The culture then underwent damped oscillations that were likely the result of a delay in the negative-feedback loop, which can be caused by the synthesis and transport of the signal, activation of LuxR, expression ofccdB, and the cascade leading to cell death byccdB. Even though negative feedback is usually associated with increased output stability, such long cascades can cause oscillations. In particular, recent measurements using a microchemostat demonstrated that the circuit was able to generate sustained oscillations for up to several hundred hours [94].
Importantly, this circuit also demonstrated that a certain degree of noise can be useful for program-ming robust dynamics via cell–cell communication. Specifically, cell-to-cell variations are essential for generating robust dynamics in the population control circuit. If cells performed identically, they would all die upon reaching the critical density that leads to a sufficient level of the killer protein. Precisely because of cell-to-cell variations, some cells died while others were left to survive and the overall population dynamics could be robustly regulated.
In addition to auto-communication within a single population, cell–cell communication has also been used to program communication between different populations and to coordinate pattern forma-tion. Basu and colleagues created a pulse generator that is activated by a signal from nearby ‘‘sender’’ cells. An interesting aspect of this interaction is the spatiotemporal behavior. Specifically, the pulse-generating cells were able to differentiate between communication from nearby and far-away sender cells [95]. This spatiotemporal behavior led to the development of a pattern formation circuit. Similar to the previous circuit, sender cells were used to produce an intercellular signal but this time the receiver cells responded to the gradient by producing a bulls-eye pattern around the chemical source [23].
20.4 Applications
In addition to decoding the ‘‘design principles’’ of biological systems, synthetic gene circuits can also find wide applications in diverse areas. These applications include innovative gene regulation systems [11,20,24,26,96–99], gene therapy [100], drug development [101], cellular computation [102], stem cell reprogramming [103], and bioremediation [104]. Here we discuss a few examples in more detail.
20.4.1 Gene Therapy
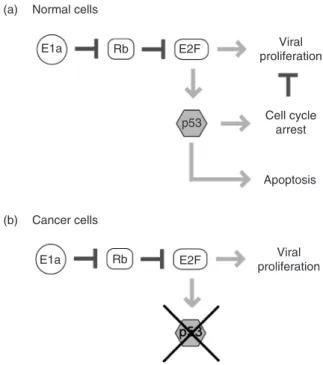
An important application area for synthetic biology is the development of innovative gene circuits for therapeutic purposes. Recent years have witnessed encouraging advancements in the field of cancer gene therapy in the creation of novel adenoviral vectors. For example, an oncolytic adenovirusdl1520, first coined by its creators [105], has shown potential use in specific targeting of tumor cells. Pioneered by McCormick and colleagues [106], ONYX-015 selectively propagates in cells with inactivated p53, a common defect in tumor cells (Figure 20.4b). On the other hand, normal cells have active p53 and ONYX-015 causes apoptosis (Figure 20.4a). Both pathways force the cell to enter the S-phase by increasing the amount of free E2F. In normal cells, the combination of entering the S-phase and a viral infection results in p53 inducing a cell cycle arrest or an apoptosis to prevent proliferation. Cancer cells with nonfunctional p53, however, will allow replication of viral particles, leading to selective killing of these cells.
The concept of selective targeting is further expanded upon by Johnson and colleagues [107]. Taking advantage of the mutations in the pRb pathway, common to most human cancers, they restrict adenoviral infection to tumor cells with high efficiency and specificity. Their strategy is based on identical dependence of both human cells and human adenovirus on the loss or mutation of Rb for uncontrolled proliferation. The E2F promoter is known to be abnormally active in tumor cells. Thus, the engineered human adenovirus ONYX-411, whose promoter of E1A or E1B has been replaced by the E2F promoter, only replicates in human tumor cells with Rb-pathway defects [107].
Although ONYX-015 is in its phase III of clinical development and the applicability of ONYX-411 is being explored with promising results, there is much room for improvement. First, effective killing of tumor cells through highly specific and efficient viral integration is most desirable. In fact, the issues of efficiency in gene integration, specificity, and effectiveness are common to many fields of gene therapy. Another limitation is variability in vector interactions with the hosts. In optimizing therapeutic benefits in various disease treatments, synthetic biology plays a key role by providing ways to design robust gene circuits that will function with increased reliability.
So far, specificity has been improved by various promoter design techniques [108–111] and increased efficiency can be achieved by the better understanding of vector integrations into cells [112–114]. Furthermore, interdisciplinary efforts and the adoption of gene delivery techniques, such as electropor-ation and ultrasound, contribute to the growth of gene therapy [115]. These technologies and method-ologies will provide ways to design more sophisticated genetic circuits that will confer improved efficiency, specificity, and effectiveness.
20.4.2 Drug Development
Many metabolites such as drugs are of natural origin but are difficult to extract from their native hosts. This rarity makes them expensive to produce in the quantities necessary to treat patients. In an effort to produce these rare drugs cheaply and in large quantities, microbes have been engineered by inserting the genes necessary to produce the drug or its precursor [20,101,116]. Bacteria are suitable hosts because they grow rapidly to produce large quantities of the drug, and it is easier to extract the product from bacteria as compared with plants [117]. In these efforts, the major challenge is to balance the expression of the drug with the host’s natural metabolic processes such that the host is not adversely affected [101]. For example, a successful attempt to produce the precursor of antimalarial drugs in bacteria was accomplished by transferring the genes fromArtimisia annua, the native plant, toE. coli. However,
(a) (b) Normal cells Cancer cells Viral proliferation Cell cycle arrest Apoptosis Viral proliferation E1a E1a Rb Rb E2F E2F p53
FIGURE 20.4 Onyx-015 viral vector circuit design for cancer gene therapy. In normal cells, (a) expression of viral vector gene E1a leads to increased E2F. An increase in E2F results in viral proliferation and increased p53. Excess p53 leads to cell cycle arrest and apoptosis, destroying the virus. On the other hand, in cancerous cells (b) p53 is inactive (marked out with an X). Viral proliferation occurs selectively in cancerous cells.
initial yields were limited by the synthase, a key enzyme in the production of the precursor. Conse-quently, the production of the synthase and other upstream molecules were increased by optimizing existing genes and introducing biosynthetic pathways fromS. cerevisiae. Using a specialized pathway from S. cerevisiae, rather than the existing pathway in E. coli, circumvented the inhibitory elements found in yeast while also bypassing those inE. coli. In the final engineeredE. colistrain that incorporated genes from different organisms, final yields of the artemisinin precursor amorphadiene were increased >10,000-fold with room for further optimization [117]. The benefits of being able to produce large quantities of rare drugs that are in high demand are quite obvious. The success of this work has recently attracted major investment from the industry [118], which will encourage broader efforts to engineer bacteria for drug and metabolite production.
20.4.3 Biotechnology Applications
By bringing discovery and utilization of novel genetic components together with a rational design approach, synthetic biology opens up tremendous possibilities for environmental engineering and biomedical discovery [119] However, use of recombinant DNA techniques invites risks commonly associated with conventional genetic engineering such as horizontal gene transfer to other species and introduction of new allergens and toxins to the environment [120,121]. Furthermore, the potential to realize desired behavior in organisms through rational design might translate into greater risks that are malevolent or inadvertent in nature [122]. A reasonable way to deal with these risks would be to make the containment of the proposed genetic circuitry an essential part of the design process. This way, the effectiveness of the containment could be made to match the estimated risk [123]. For example, containment in bacteria can be achieved through a built-in safeguard mechanism that can be imple-mented using the quorum sensing based population control discussed earlier [22].
One practical application area for synthetic biology, where aforementioned risks should be minim-ized, is environmental biotechnology. Microorganisms are commonly used for pollution reduction (bioremediation) and can provide safe and cost-effective alternatives to existing physicochemical methods [124]. However, risks and concerns associated with the release of recombinant microorganisms into the environment [125] currently limit their widespread use. Therefore, researchers are designing containment systems that will minimize horizontal gene transfer and uncontrolled proliferation of recombinant microorganisms (for a good review see Ref. [104]). In order to realize the full potential of synthetic biology in this area and answer safety concerns, microorganisms should be equipped with not only biodegradation pathways but also conditional lethality systems that will ensure control over the genetic material being released. One striking example of coupling of a biodegradation pathway with a containment system is achieved by engineering a streptavidin based lethality system that is controlled by TOL plasmid for aromatic hydrocarbon metabolism [126] In this system, the TOL pathway, which is capable of breaking down toxic aromatic compounds, is used to control the expression of a streptavidin gene, which binds to biotin, an essential coenzyme for microorganisms. Depletion of the environmental inducer of the TOL pathway,m-methylbenzoate, led to streptavidin expression and eventually cell death with remarkable efficiencies around 10 7–10 8mutant escape rates [126].
Containment is an important issue for synthetic constructs; moreover, synthetic biology can address this issue and tackle environmental pollution problems by introducing new perspectives and techniques. Although the use of genetic engineering for biodegradation is not novel [127], design ideas from basic circuits to improve robustness against mutations, and attain control at the level of populations rather than at the level of individuals [22,128] can improve killing efficiencies of containment systems.
20.5 Outlook
We have presented recent progress in the nascent field of synthetic biology and highlighted a few application areas. We believe that synthetic biology has great potential for numerous applications that go well beyond what we can envision today. Nevertheless, many challenges still remain. A major hurdle is
the elucidation of control mechanisms that can ensure robust function of synthetic gene circuits. To this end, synthetic biologists will benefit from rapid advances in systems biology [65,129–132]. On one hand, efforts in systems biology will continue to expand the list of building blocks that serve as the foundation of synthetic gene circuits. On the other, they will likely generate substantial lessons for optimal circuit design from naturally biological systems [133–135].
Another challenge lies in the measurement and monitoring of intracellular and intercellular dynamics with high precision. For de novo engineering of gene circuits, it has become clear that precise characterization of circuit dynamics critically depends on well-controlled experimental conditions and distinct cellular markers. To address this issue, the marriage between biology and nanotechnology can be particularly promising [34].
Fine control of growth environments can be achieved using novel microfluidics devices [136,137]. For instance, Balagadde and colleagues recently devised a microchemostat [94], an automated and continu-ously operated microfluidic device with a reactor volume of 16 nL, which is an order of magnitude smaller than conventional culturing systems. When applied to the population control circuit described above, the microchemostat significantly delayed the process of mutants arising and enabled measure-ments of complex population dynamics for more than 500 h with single-cell resolution. This work has focused on observation of cell morphology in addition to cell density, but it can be readily adapted to measure gene expression dynamics reported by fluorescence or luminescence.
Complementary to culturing systems, a wide array of nanosensors has been developed to facilitate analysis of single cells and protein dynamics inside the cell [138–141]. We anticipate that these diverse micro- and nanoscale devices can be integrated for elucidating temporal–spatial dynamics of cellular processes with ever-increasing resolution.
Acknowledgments
We thank Chee Meng Tan, Jun Ozaki, and Guang Yao for their comments and suggestions on the chapter.
References
1. Barkai, N., and S. Leibler. 1997. Robustness in simple biochemical networks. Nature 387 (6636):913–917.
2. Alon, U., M.G. Surette, N. Barkai, and S. Leibler. 1999. Robustness in bacterial chemotaxis.Nature 397 (6715):168–171.
3. Hasty, J., D. McMillen, and J.J. Collins. 2002. Engineered gene circuits.Nature420 (6912):224–230. 4. Little, J.W., D.P. Shepley, and D.W. Wert. 1999. Robustness of a gene regulatory circuit.EMBO J18
(15):4299–4307.
5. Becskei, A., and L. Serrano. 2000. Engineering stability in gene networks by autoregulation.Nature 405 (6786):590–593.
6. Mangan, S., and U. Alon. 2003. Structure and function of the feed-forward loop network motif. Proc Natl Acad Sci USA100 (21):11980–11985.
7. Miller, M.B., and B.L. Bassler. 2001. Quorum sensing in bacteria.Annu Rev Microbiol55:165–199. 8. Sprinzak, D., and M.B. Elowitz. 2005. Reconstruction of genetic circuits.Nature438 (7067):443–448. 9. Benner, S.A., and A.M. Sismour. 2005. Synthetic biology.Nat Rev Genet6 (7):533–543.
10. Ferber, D. 2004. Synthetic biology. Microbes made to order.Science303 (5655):158–161.
11. Kramer, B.P., A.U. Viretta, M. Daoud-El-Baba, D. Aubel, W. Weber, and M. Fussenegger. 2004. An engineered epigenetic transgene switch in mammalian cells.Nat Biotechnol22 (7):867–870. 12. Gardner, T.S., C.R. Cantor, and J.J. Collins. 2000. Construction of a genetic toggle switch in
Escherichia coli.Nature403 (6767):339–342.
13. Becskei, A., B. Seraphin, and L. Serrano. 2001. Positive feedback in eukaryotic gene networks: cell differentiation by graded to binary response conversion.EMBO J20 (10):2528–2535.
14. Isaacs, F.J., J. Hasty, C.R. Cantor, and J.J. Collins. Prediction and measurement of an autoregulatory genetic module.Proc Natl Acad Sci USA100 (13):7714–7719.
15. Elowitz, M.B., and S. Leibler. 2000. A synthetic oscillatory network of transcriptional regulators. Nature403 (6767):335–338.
16. Fung, E., W.W. Wong, J.K. Suen, T. Bulter, S.G. Lee, and J.C. Liao. 2005. A synthetic gene-metabolic oscillator.Nature435 (7038):118–122.
17. Weiss, R., G.E. Homsy, and T. Knight, Jr.1999. Toward in vivo digital circuits. InDimacs workshop on evolution as computation, 275–295. Princeton: Springer.
18. Guet, C.C., M.B. Elowitz, W. Hsing, and S. Leibler. 2002. Combinatorial synthesis of genetic networks.Science296 (5572):1466–1470.
19. Yokobayashi, Y., R. Weiss, and F.H. Arnold. 2002. Directed evolution of a genetic circuit.Proc Natl Acad Sci USA99 (26):16587–16591.
20. Farmer, W.R., and J.C. Liao. 2000. Improving lycopene production inEscherichia coliby engineer-ing metabolic control.Nat Biotechnol18 (5):533–537.
21. Rackham, O., and J. Chin. 2005. A network of orthogonal ribosome.mRNA pairs.Nat Chem Biol 1:159–166.
22. You, L., R.S. Cox, 3rd, R. Weiss, and F.H. Arnold. 2004. Programmed population control by cell– cell communication and regulated killing.Nature428 (6985):868–871.
23. Basu, S., Y. Gerchman, C.H. Collins, F.H. Arnold, and R.A. Weiss. A synthetic multicellular system for programmed pattern formation.Nature434 (7037):1130–1134.
24. Bulter, T., S.G. Lee, W.W. Wong, E. Fung, M.R. Connor, and J. C. Liao. 2004. Design of artificial cell– cell communication using gene and metabolic networks.Proc Natl Acad Sci USA101 (8):2299–2304. 25. Chan, L.Y., S. Kosuri, and D. Endy. 2005. Refactoring bacteriophage T7. Mol Syst Biol 1 (1):
msb4100025-E1.
26. Kobayashi, H., M. Kaern, M. Araki, K. Chung, T.S. Gardner, C.R. Cantor, and J. J. Collins. 2004. Programmable cells: Interfacing natural and engineered gene networks.Proc Natl Acad Sci USA101 (22):8414–8419.
27. Noireaux, V., R. Bar-Ziv, and A. Libchaber. 2003. Principles of cell-free genetic circuit assembly. Proc Natl Acad Sci USA100 (22):12672–12677.
28. Isalan, M., C. Lemerle, and L. Serrano. 2005. Engineering gene networks to emulate Drosophila embryonic pattern formation.PLoS Biol3 (3):e64.
29. Bader, G.D., D. Betel, and C.W. Hogue. 2003. BIND: the Biomolecular Interaction Network Database.Nucleic Acids Res31 (1):248–250.
30. Xenarios, I., L. Salwinski, X.J. Duan, P. Higney, S.M. Kim, and D. Eisenberg. 2002. DIP, the Database of Interacting Proteins: A research tool for studying cellular networks of protein inter-actions.Nucleic Acids Res30 (1):303–305.
31. Keseler, I.M., J. Collado-Vides, S. Gama-Castro, J. Ingraham, S. Paley, I.T. Paulsen, M. Peralta-Gil, and P.D. Karp. 2005. EcoCyc: A comprehensive database resource forEscherichia coli.Nucleic Acids Res33 (Database issue):D334–337.
32. Wixon, J., and D. Kell. 2000. The Kyoto encyclopedia of genes and genomes—KEGG. Yeast17 (1):48–55.
33. Tian, J., H. Gong, N. Sheng, X. Zhou, E. Gulari, X. Gao, and G. Church. 2004. Accurate multiplex gene synthesis from programmable DNA microchips.Nature432 (7020):1050–1054.
34. Philip, B. 2005. Synthetic biology for nanotechnology.Nanotechnology16 (1):R1.
35. Ptashne, M. 2004.A genetic switch: Phage lambda revisited. 3rd ed. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
36. Ferrell, J.E. Jr. 2002. Self-perpetuating states in signal transduction: Positive feedback, double-negative feedback and bistability.Curr Opin Cell Biol14 (2):140–148.
37. Andersen, J.B., C. Sternberg, L.K. Poulsen, S.P. Bjorn, M. Givskov, and S. Molin. 1998. New unstable variants of green fluorescent protein for studies of transient gene expression in bacteria. Appl Environ Microbiol64 (6):2240–2246.
38. Novak, B., and J.J. Tyson. 2004. A model for restriction point control of the mammalian cell cycle.J Theor Biol230 (4):563–579.
39. Panda, S., J.B. Hogenesch, and S.A. Kay. 2002. Circadian rhythms from flies to human.Nature417 (6886):329–335.
40. Harmer, S.L., S. Panda, and S.A. Kay. 2001. Molecular bases of circadian rhythms.Annu Rev Cell Dev Biol17:215–253.
41. Francois, P., and V. Hakim. 2004. Design of genetic networks with specified functions by evolution in silico.Proc Natl Acad Sci USA101 (2):580–585.
42. Arnold, F.H. 2001. Combinatorial and computational challenges for biocatalyst design.Nature409 (6817):253–257.
43. Yuan, L., I. Kurek, J. English, and R. Keenan. 2005. Laboratory-directed protein evolution. Micro-biol Mol Biol Rev69 (3):373–392.
44. Farinas, E.T., T. Bulter, and F.H. Arnold. 2001. Directed enzyme evolution.Curr Opin Biotechnol12 (6):545–551.
45. Umeno, D., A.V. Tobias, and F.H. Arnold. 2005. Diversifying carotenoid biosynthetic pathways by directed evolution.Microbiol Mol Biol Rev69 (1):51–78.
46. Collins, C.H., F.H. Arnold, and J.R. Leadbetter. 2005. Directed evolution of Vibrio fischeri LuxR for increased sensitivity to a broad spectrum of acyl-homoserine lactones.Mol Microbiol55 (3):712– 720.
47. Hasty, J. 2002. Design then mutate.Proc Natl Acad Sci USA99 (26):16516–16518.
48. Yohei, Y., and H.A. Frances. 2005. A dual selection module for directed evolution of genetic circuits.Nat Comput—Int J4 (3):245–254.
49. Blake, W.J., M, KAErn, C.R. Cantor, and J.J. Collins. 2003. Noise in eukaryotic gene expression. Nature422 (6932):633–637.
50. Rao, C.V., D.M. Wolf, and A.P. Arkin. 2002. Control, exploitation and tolerance of intracellular noise.Nature420 (6912):231–237.
51. McAdams, H.H., and A. Arkin. 1999. It’s a noisy business! Genetic regulation at the nanomolar scale.Trends Genet15 (2):65–69.
52. McAdams, H.H., and A. Arkin. 1997. Stochastic mechanisms in gene expression.Proc Natl Acad Sci USA94 (3):814–819.
53. Thattai, M., and A. van Oudenaarden. 2001. Intrinsic noise in gene regulatory networks.Proc Natl Acad Sci USA98 (15):8614–8619.
54. Ozbudak, E.M., M. Thattai, I. Kurtser, A.D. Grossman, and A. van Oudenaarden. Regulation of noise in the expression of a single gene.Nat Genet31 (1):69–73.
55. Levsky, J.M., and R.H. Singer. 2003. Gene expression and the myth of the average cell.Trends Cell Biol13 (1):4–6.
56. Elowitz, M.B., A.J. Levine, E.D. Siggia, and P.S. Swain. 2002. Stochastic gene expression in a single cell.Science297 (5584):1183–1186.
57. Guptasarma, P. 1995. Does replication-induced transcription regulate synthesis of the myriad low copy number proteins ofEscherichia coli?Bioessays17 (11):987–997.
58. Colman-Lerner, A., A. Gordon, E. Serra, T. Chin, O. Resnekov, D. Endy, C.G. Pesce, and R. Brent. 2005. Regulated cell-to-cell variation in a cell-fate decision system.Nature437 (7059):699–706. 59. Raser, J.M., and E.K. O’Shea. 2004. Control of stochasticity in eukaryotic gene expression.Science
304 (5678):1811–1814.
60. Korobkova, E., T. Emonet, J.M. Vilar, T.S. Shimizu, and P. Cluzel. 2004. From molecular noise to behavioural variability in a single bacterium.Nature428 (6982):574–578.
61. Weinberger, L.S., J.C. Burnett, J.E. Toettcher, A.P. Arkin, and D.V. Schaffer. 2005. Stochastic gene expression in a lentiviral positive-feedback loop: HIV-1 Tat fluctuations drive phenotypic diversity. Cell122 (2):169–182.
62. A. Arkin, J. Ross, and H.H. McAdams. 1998. Stochastic kinetic analysis of developmental pathway bifurcation in phage lambda-infectedEscherichia colicells.Genetics149 (4):1633–1648.
63. Mihalcescu, I., W. Hsing, and S. Leibler. 2004. Resilient circadian oscillator revealed in individual cyanobacteria.Nature430 (6995):81–85.
64. Rao, C.V., and A.P. Arkin. 2001. Control motifs for intracellular regulatory networks.Annu Rev Biomed Eng3:391–419.
65. You, L. 2004. Toward computational systems biology.Cell Biochem Biophys40 (2):167–184. 66. Kaern, M., T.C. Elston, W.J. Blake, and J.J. Collins. 2005. Stochasticity in gene expression: From
theories to phenotypes.Nat Rev Genet6 (6):451–464.
67. Savageau, M.A. 1974. Comparison of classical and autogenous systems of regulation in inducible operons.Nature252 (5484):546–549.
68. Hillen, W., and C. Berens. 1994. Mechanisms underlying expression of Tn10 encoded tetracycline resistance.Annu Rev Microbiol48:345–369.
69. Batchelor, E., T.J. Silhavy, and M. Goulian. 2004. Continuous control in bacterial regulatory circuits.J Bacteriol186 (22):7618–7625.
70. Yi, T.M., Y. Huang, M.I. Simon, and J. Doyle. 2000. Robust perfect adaptation in bacterial chemotaxis through integral feedback control.Proc Natl Acad Sci USA97 (9):4649–4653. 71. Savageau, M.A. 1976.Biochemical systems analysis. Reading, MA: Addison-Wesley.
72. Austin, D.W., M.S. Allen, J.M. McCollum, R.D. Dar, J.R. Wilgus, G.S. Sayler, N.F. Samatova, C.D. Cox, and M.L. Simpson. 2006. Gene network shaping of inherent noise spectra.Nature439 (7076):608–611.
73. Rosenfeld, N., M.B. Elowitz, and U. Alon. 2002. Negative autoregulation speeds the response times of transcription networks.J Mol Biol323 (5):785–793.
74. Ferrell, J.E. Jr., and E.M. Machleder. 1998. The biochemical basis of an all-or-none cell fate switch inXenopus oocytes.Science280 (5365):895–898.
75. Bagowski, C.P., and J.E. Ferrell. Jr. 2001. Bistability in the JNK cascade.Curr Biol11 (15):1176–1182. 76. Hasty, J., D. McMillen, F. Isaacs, and J.J. Collins. 2001. Computational studies of gene regulatory
networks: In numero molecular biology.Nat Rev Genet2 (4):268–279.
77. Barkai, N., and S. Leibler. 2000. Circadian clocks limited by noise.Nature403 (6767):267–268. 78. Atkinson, M.R., M.A. Savageau, J.T. Myers, and A.J. Ninfa. 2003. Development of genetic circuitry
exhibiting toggle switch or oscillatory behavior inEscherichia coli.Cell113 (5):597–607.
79. Acar, M., A. Becskei, and A. van Oudenaarden. 2005. Enhancement of cellular memory by reducing stochastic transitions.Nature435 (7039):228–232.
80. Ruby, E.G., and M.J. McFall-Ngai. 1992. A squid that glows in the night: Development of an animal-bacterial mutualism.J Bacteriol174 (15):4865–4870.
81. Fuqua, C., M.R. Parsek, and E.P. Greenberg. 2001. Regulation of gene expression by cell-to-cell communication: acyl-homoserine lactone quorum sensing.Annu Rev Genet35:439–468.
82. Piper, K.R., S. Beck von Bodman, and S.K. Farrand. 1993. Conjugation factor ofAgrobacterium tumefaciensregulates Ti plasmid transfer by autoinduction.Nature362 (6419):448–450.
83. Hinton, J.C., J.M. Sidebotham, L.J. Hyman, M.C. Perombelon, and G.P. Salmond. 1989. Isolation and characterisation of transposon-induced mutants of Erwinia carotovora subsp. atroseptica exhibiting reduced virulence.Mol Gen Genet217 (1):141–148.
84. Davies, D.G., M.R. Parsek, J.P. Pearson, B.H. Iglewski, J.W. Costerton, and E.P. Greenberg. 1998. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 280 (5361):295–298.
85. Steinmoen, H., E. Knutsen, and L.S. Havarstein. 2002. Induction of natural competence in Streptococcus pneumoniaetriggers lysis and DNA release from a subfraction of the cell population. Proc Natl Acad Sci USA99 (11):7681–7686.
86. Whitehead, N.A., A.M. Barnard, H. Slater, N.J. Simpson, and G.P. Salmond. 2001. Quorum-sensing in Gram-negative bacteria.FEMS Microbiol Rev25 (4):365–404.
87. Lewis, K. 2000. Programmed death in bacteria.Microbiol Mol Biol Rev64 (3):503–514.
88. Redfield, R.J. 2002. Is quorum sensing a side effect of diffusion sensing? Trends Microbiol 10 (8):365–370.
89. Garcia-Ojalvo, J., M.B. Elowitz, and S.H. Strogatz. 2004. Modeling a synthetic multicellular clock: Repressilators coupled by quorum sensing.Proc Natl Acad Sci USA101 (30):10955–10960. 90. Wang, R., and L. Chen. 2005. Synchronizing genetic oscillators by signaling molecules. J Biol
Rhythms20 (3):257–269.
91. McMillen, D., N. Kopell, J. Hasty, and J.J. Collins. 2002. Synchronizing genetic relaxation oscillators by intercell signaling.Proc Natl Acad Sci USA99 (2):679–684.
92. Kuznetsov A., M. Kaem, and N. Kopell. 2004. Synchrony in a population of hysteresis-based genetic oscillators.SIAM J Appl Math65:392–342.
93. Wagner, A. 2005. Circuit topology and the evolution of robustness in two-gene circadian oscil-lators.Proc Natl Acad Sci USA102 (33):11775–11780.
94. Balagadde, F.K., L. You, C.L. Hansen, F.H. Arnold, and S.R. Quake. 2005. Long-term monitoring of bacteria undergoing programmed population control in a microchemostat. Science 309 (5731):137–140.
95. Basu, S., R. Mehreja, S. Thiberge, M.T. Chen, and R. Weiss.2004. Spatiotemporal control of gene expression with pulse-generating networks.Proc Natl Acad Sci USA101 (17):6355–6360.
96. Chen, W., P.T. Kallio, and J.E. Bailey. 1993. Construction and characterization of a novel cross-regulation system for regulating cloned gene expression inEscherichia coli.Gene130 (1):15–22. 97. Khlebnikov, A., O. Risa, T. Skaug, T.A. Carrier, and J.D. Keasling. 2000. Regulatable
arabinose-inducible gene expression system with consistent control in all cells of a culture.J Bacteriol182 (24):7029–7034.
98. Bayer, T.S., and C.D. Smolke. 2005. Programmable ligand-controlled riboregulators of eukaryotic gene expression.Nat Biotechnol23 (3):337–343.
99. Isaacs, F.J., D.J. Dwyer, C. Ding, D.D. Pervouchine, C.R. Cantor, and J.J. Collins. 2004. Engi-neered riboregulators enable post-transcriptional control of gene expression. Nat Biotechnol22 (7):841–847.
100. Bischoff, J.R., D.H. Kirn, A. Williams, C. Heise, S. Horn, M. Muna, L. Ng, J.A. Nye, A. Sampson-Johannes, A. Fattaey, and F. McCormick. 1996. An adenovirus mutant that replicates selectively in p53-deficient human tumor cells.Science274 (5286):373–376.
101. Khosla, C., and J.D. Keasling. 2003. Metabolic engineering for drug discovery and development. Nat Rev Drug Discov2 (12):1019–1025.
102. Simpson, M.L., G.S. Sayler, J.T. Fleming, and B. Applegate. 2001. Whole-cell biocomputing.Trends Biotechnol19 (8):317–320.
103. Lemischka, I.R. 2005. Stem cell biology: A view toward the future.Ann NY Acad Sci1044:132–138. 104. Paul, D., G. Pandey, and R.K. Jain. 2005. Suicidal genetically engineered microorganisms for
bioremediation: Need and perspectives.Bioessays27 (5):563–573.
105. Barker, D.D., and A.J. Berk. 1987. Adenovirus proteins from both E1B reading frames are required for transformation of rodent cells by viral-infection and DNA transfection.Virology156 (1):107–121. 106. Bischoff, J.R., D.H. Kim, A. Williams, C. Heise, S. Horn, M. Muna, L. Ng, J. A. Nye, A.
Sampson-Johannes, A. Fattaey, and F. McCormick. 1996. An adenovirus mutant that replicates selectively in p53-deficient human tumor cells.Science274 (5286):373–376.
107. Johnson, L., A. Shen, L. Boyle, J. Kunich, K. Pandey, M. Lemmon, T. Hermiston, M. Giedlin, F. McCormick, and A. Fattaey. 2002. Selectively replicating adenoviruses targeting deregulated E2F activity are potent, systemic antitumor agents.Cancer Cell1 (4):325–337.
108. Vile, R.G., and I.R. Hart. 1993.In vitroandin vivotargeting of gene expression to melanoma cells. Cancer Res53 (5):962–967.
109. Vile, R.G., R.M. Diaz, N. Miller, S. Mitchell, A. Tuszyanski, and S.J. Russell. 1995. Tissue-specific gene expression from Mo-MLV retroviral vectors with hybrid LTRs containing the murine tyrosi-nase enhancer=promoter.Virology214 (1):307–313.
110. Hughes, B.W., A.H. Wells, Z. Bebok, V.K. Gadi, R.I. Garver Jr., W.B. Parker, and E.J. Sorscher. 1995. Bystander killing of melanoma cells using the human tyrosinase promoter to express the Escher-ichia colipurine nucleoside phosphorylase gene.Cancer Res55 (15):3339–3345.
111. Diaz, R.M., T. Eisen, I.R. Hart, and R.G. Vile. 1998. Exchange of viral promoter=enhancer elements with heterologous regulatory sequences generates targeted hybrid long terminal repeat vectors for gene therapy of melanoma.J Virol72 (1):789–795.
112. Flotte, T.R. 2004. Gene therapy progress and prospects: Recombinant adeno-associated virus (rAAV) vectors.Gene Ther11 (10):805–810.
113. Sinn, P.L., S.L. Sauter, and P.B. McCray. Jr. 2005. Gene therapy progress and prospects: develop-ment of improved lentiviral and retroviral vectors—Design, biosafety, and production.Gene Ther 12 (14):1089–1098.
114. St George, J.A. 2003. Gene therapy progress and prospects: Adenoviral vectors. Gene Ther 10 (14):1135–1141.
115. Wells, D.J. 2004. Gene therapy progress and prospects: Electroporation and other physical methods.Gene Ther11 (18):1363–1369.
116. Bailey, J.E. 1991. Toward a science of metabolic engineering.Science252 (5013):1668–1675. 117. Martin, V.J., D.J. Pitera, S.T. Withers, J.D. Newman, and J.D. Keasling. 2003. Engineering a
mevalonate pathway in Escherichia coli for production of terpenoids. Nat Biotechnol 21 (7): 796–802.
118. Herrera, S. 2005. Synthetic biology offers alternative pathways to natural products.Nat Biotechnol 23 (3):270–271.
119. McDaniel, R
AQ1 ., and R. Weiss. Forthcoming. Advances in synthetic biology: On the path from prototypes to applications.Curr Opin Biotechnol. Corrected Proof.
120. Berg, P., D. Baltimore, S. Brenner, R.O. Roblin III, and M.F. Singer. 1975. Asilomar conference on recombinant DNA molecules.Science188 (4192):991–994.
121. Ball, P. 2004. Synthetic biologystarting from scratch.Nature431 (7009):624.
122. 2004. Futures of artificial life.Nature431 (7009):613. AQ2 120. Berg, P., D. Baltimore, S. Brenner, R.O. Roblin, and M.F. Singer. 1975. Summary statement of the
Asilomar conference on recombinant DNA molecules.Proc Natl Acad Sci USA72 (6):1981–1984. 124. Pieper, D.H., and W. Reineke. 2000. Engineering bacteria for bioremediation.Curr Opin Biotechnol
11 (3):262–270.
125. Wilson, M., and S.E. Lindow. 1993. Release of recombinant microorganisms.Annu Rev Microbiol 47:913–944.
126. Kaplan, D.L., C. Mello, T. Sano, C. Cantor, and C. Smith.1999. Streptavidin-based containment systems for genetically engineered microorganisms.Biomol Eng16 (1–4):135–140.
127. Ramos, J.L., A. Wasserfallen, K. Rose, and K.N. Timmis. 1987. Redesigning metabolic routes: Manipulation of TOL plasmid pathway for catabolism of alkylbenzoates.Science235 (4788):593– 596.
128. Andre´, J.B., and G. Bernard. 2005. Multicellular organization in bacteria as a target for drug therapy.Ecol Lett8 (8):800–810.
129. Ideker, T., T. Galitski, and L. Hood. 2001. A new approach to decoding life: Systems biology.Annu Rev Genomics Hum Genet2:343–372.
130. Gilman, A., and A.P. Arkin. 2002. Genetic ‘‘code’’: Representations and dynamical models of genetic components and networks.Annu Rev Genomics Hum Genet3:341–369.
131. Palsson, B. 2000. The challenges of in silico biology.Nat Biotechnol18 (11):1147–1150. 132. Endy, D. 2005. Foundations for engineering biology.Nature438 (7067):449–453.
133. Tyson, J.J., K. Chen, and B. Novak. 2001. Network dynamics and cell physiology.Nat Rev Mol Cell Biol2 (12):908–916.
134. Kaern, M., W.J. Blake, and J.J. Collins. 2003. The engineering of gene regulatory networks.Annu Rev Biomed Eng5:179–206.
135. Wall, M.E., W.S. Hlavacek, and M.A. Savageau. 2004. Design of gene circuits: Lessons from bacteria.Nat Rev Genet5 (1):34–42.
136. Quake, S.R., and A. Scherer. 2000. From micro- to nanofabrication with soft materials.Science290 (5496):1536–1540.
137. Hong, J.W., and S.R. Quake. 2003. Integrated nanoliter systems.Nat Biotechnol21 (10):1179–1183. 138. Thorsen, T., S.J. Maerkl, and S.R. Quake. 2002. Microfluidic large-scale integration. Science 298
(5593):580–584.
139. Cullum, B.M., G.D. Griffin, G.H. Miller, and T. Vo-Dinh. 2000. Intracellular measurements in mammary carcinoma cells using fiber-optic nanosensors.Anal Biochem277 (1):25–32.
140. Vo-Dinh, T., and P. Kasili. 2005. Fiber-optic nanosensors for single-cell monitoring.Anal Bioanal Chem382 (4):918–925.
141. Vo-Dinh, T., J.P. Alarie, B.M. Cullum, and G.D. Griffin. 2000. Antibody-based nanoprobe for measurement of a fluorescent analyte in a single cell.Nat Biotechnol18 (7):764–767.
Author Queries
[AQ1] Kindly update reference 119. [AQ2] Please supply author names.