Original Article
Long noncoding RNA PDIA3P promotes breast
cancer development by regulating miR-183/
ITGB1/FAK/PI3K/AKT/β-catenin signals
Xuekui Wang, Shen Gao, Huan Chen, Lihong Li, Chengyan He, Ling Fang
Department of Breast and Thyroid Surgery, China-Japan Union Hospital of Jilin University, Changchun, Jilin, China
Received December 11, 2018; Accepted January 30, 2019; Epub April 1, 2019; Published April 15, 2019
Abstract: Aberrantly expressed long noncoding RNAs (lncRNAs) play crucial roles in the process of breast cancer
(BC). This research aims to dig the possible roles and regulatory mechanism of lncRNA protein disulfide isomerase
family A member 3 pseudogene 1 (PDIA3P1) in BC. The mRNA level of PDIA3P both in BC tissues and in cells were determined, followed by the investigation of the effects of PDIA3P suppression on cell biological processes includ-ing the viability, apoptosis, migration and invasion. Furthermore, whether PDIA3P modulated the expression of
integrin β1 (ITGB1) expression by competitively sponging miR-183 and then regulated the activation of FAK/PI3K/ AKT/β-catenin pathway was explored. PDIA3P was discovered as up-regulated in BC tissues and cells. PDIA3P sup -pression markedly decreased cell viability, promoted apoptosis, and inhibited migration and invasion in MCF-7 cells. In addition, PDIA3P was found to be negatively interacting with miR-183, and PDIA3P regulated tumor growth and metastasis through negatively regulating miR-183. Moreover, ITGB1 was targeted by miR-183 and involved in tumor
growth and metastasis. Lastly, PDIA3P suppression markedly inhibited the activation of FAK/PI3K/AKT/β-catenin pathway, which was significantly reversed after simultaneous inhibition of miR-183. Our results reveal that PDIA3P may forpromote BC development by sponging miR-183 to regulate ITGB1, thus inducing the activation of FAK/PI3K/ AKT/β-catenin signals. PDIA3P may serve as a promising biomarker or target for the detection or treatment of BC.
Keywords: Breast cancer, long noncoding RNA, protein disulfide isomerase family A member 3 pseudogene 1, miR-183, integrin β1
Introduction
Breast cancer (BC) remains one of the most fre-quently diagnosed cancer in women worldwide, with an estimated 232670 newly diagnosed cases and 40000 deaths in the United States [1]. Most patients with BC are diagnosed at an advanced stage due to difficulties in early detection, greatly reducing the quality of the patients’ lives [2]. The five-year survival rate is less than 10% because of the predilection for bone metastasis [3]. For improving the clinical outcome, it is still imperative to deepen under-standing of the pathogenesis of BC.
Increasing evidence proves the significant roles of long non-coding RNAs (lncRNAs), which are more than 200 nucleotides in length, are wide-ly involved in diverse cellular processes [4, 5]. Aberrantly expressed lncRNAs are frequently observed in various tumors and may be used
for cancer diagnosis and prognosis due to their diverse functions in mediating tumor initiation and metastasis [6-8]. lncRNA DSCAM-AS1 plays a crucial role in the process of disease [9]. Other lncRNAs, such as LncRNA Inhibiting Metastasis (LIMT) [10], Growth arrest-specific 5 (GAS5) [11], and Esrp2-antisense [12] have been implicated in progression of BC.
In the current study, we detected the mRNA level of PDIA3P in cancer tissues and cells and also investigated the effects of PDIA3P sup-pression on tumor cell biological processes. Furthermore, lncRNAs are emerging as impor-tant regulators of gene expression during tumorigenesis through serving as competing endogenous RNAs (ceRNAs). We explored whether PDIA3P modulated the expression of integrin β1 (ITGB1) by competitively sponging miR-183 and then regulated the activation of FAK/PI3K/AKT/β-catenin signals. All of these findings were to disclose the detailed role and molecular mechanisms of PDIA3P in BC development.
Materials and methods
Human tissue samples
In total, 50 patients with BC from December 2012 to January 2018 were enrolled in this study. Tumor tissues and the adjacent non-tumor tissues were collected by surgical resec-tion, and then were quickly preserved in liquid nitrogen following resection and stored at -80°C. This study was proved by the ethics committee of our hospital and informed con-sents for research were obtained from all patients.
Cell lines and cell transfection
Human BC cell lines MCF-7 and MDA-MB-231 as well as normal human mammary epithelial cell line MCF10A, obtained from American Type Culture Collection (ATCC), were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Gibco, USA) containing 10% fetal bovine serum (FBS, Gibco) in a 37°C humidified incubator with 5% CO2. Cells were passaged every 2-3 days for maintaining exponential growth. For cell transfection, sh-PDIA3P, sh-NC, miR-183 mimic, mimic NC, inhibitor NC, miR-183 inhibi-tor, si-NC and/or si-ITGB1 were transfected into MCF-7 cells using Lipofectamine 2000 (Invitrogen) following the manufacturer’s in- structions.
Quantitative real-time reverse transcription PCR (qRT-PCR)
Total RNA from tissues/cells was isolated using Trizol (Invitrogen), and reverse transcription to cDNA was subsequently conducted using the
Omniscript RT Kit (Qiagen). The PCR reaction mixture was then prepared, including 0.3 mM of each primer, 1 Fast SYBR® Green Master Mix (Applied Biosystems), and 50 ng of cDNA. qRT-PCR analyses were performed on a StepOnePlus™ (Applied Biosystems). For rela -tive quantification, the threshold cycle values were normalized to GAPDH, and the relative gene expression was quantitated by 2-ΔΔCt method.
Cell viability assay
Cell viability was assessed using MTT assay (Roche Applied Science, Penzberg, Germany). Cells with different transfections were grown in 96-well plates. After 24 h of transfection, cell viability in each group was assessed by mea-suring the absorbance (470 nm) with a MRX II absorbance reader (DYNEX Technologies, Chantilly, Virginia, USA).
Cell apoptosis
Cells were harvested after 48 h of different transfections. The apoptotic cells were ana-lyzed using the FITC-Annexin V Apoptosis Detection Kit (BD Biosciences) according to the manufacturer’s recommendations, cells were double stained with FITC-Annexin V and prop-idium iodide (PI), and then analyzed with a flow cytometry (FACScan; BD Biosciences) equipped with a CellQuest software (BD Biosciences). Viable cells, early/late apoptotic cells, and dead cells were distinguished, and the percent-age of apoptotic cells was calculated.
Cell migration and invasion assays
Cell migration and invasion assay was per-formed using Transwell assay. For the migration assay, 5 × 104 cells were harvested after 48 h
Western blot
The total protein from MCF-7 cells with different transfections was extracted by lysing with cell lysis buffer (Beyotime, Haimen, China). After being quantified using the BCA™ Protein Assay Kit (Pierce, Appleton, WI, USA), the proteins (30 μg per lane) were separated on 12% SDS-PAGE gels and transferred onto polyvinylidene fluo -ride (PVDF) membranes (Millipore, Billerica, MA, USA). The membranes were then incubat-ed with the primary antibodies to cas-pase-3, cleaved-cascas-pase-3, Bax, Bcl-2, pro-caspase-9, cleaved-pro-caspase-9, ITGB1, FAK, p-FAK, PI3K, p-PI3K, AKT, p-AKT, β-catenin, and β-actin (1:1000 dilution; Abcam, Cambridge, UK) overnight at 4°C and recommended sec -ondary antibodies (1:5000 dilution; Abcam) for 2 h. β-actin was used as the control. The pro -tein signals were revealed using ECL detection kit (Pierce, Rockford, IL, USA) and analyzed using an Odyssey Infrared Imaging System (LI-COR, Lincoln, NE, USA).
Luciferase reporter assay
The pMIR-REPORT-ITGB1-WT/MUT (Obio, Sh-anghai, China) were constructed and then co-transfected into MCF-7 cells with miR-183 mimic or mimic control. After 48 h of transfec-tion, luciferase activity of each group was detected using the Dual-Luciferase Reporter Assay System (Promega).
Statistical analysis
All experiments were independently repeated three times. Statistical analyses were per-formed with SPSS version 18.0 (SPSS, Chicago, IL). Statistical differences between groups were compared by Student’s t test or one-way ANOVA, and P < 0.05 was considered as statis -tically significant.
Results
PDIA3P is upregulated in BC tissues and cells
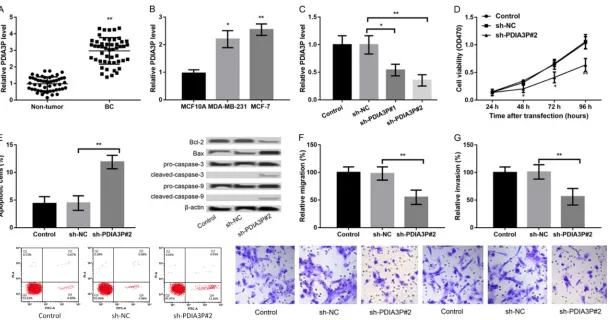
PDIA3P was up-regulated in BC tissues in rela-tion to that in adjacent normal tissues (P < 0.01, Figure 1A), indicating a possible onco-genic role of PDIA3P in BC. Moreover, PDIA3P was also highly expressed in both MCF-7 and MDA-MB-231 cells in comparison with that in normal mammary MCF10A cells (P < 0.05, Figure 1B). We found that PDIA3P expression in MCF-7 cells was higher than MDA-MB-231
cells, so MCF-7 cells were selected for following experiments.
Suppression of PDIA3P decreases cell viability, promotes apoptosis, and inhibits migration and invasion
PDIA3P was suppressed in MCF-7 cells, fol-lowed by detection of the effects of PDIA3P suppression on cell viability, apoptosis, migra-tion and invasion. In comparison with sh-NC transfection, transfection with sh-PDIA3P#1 or sh-PDIA3P#2 resulted in the significantly decreased expression of PDIA3P in MCF-7 cells (P < 0.05, Figure 1C), and sh-PDIA3P#2 was selected for subsequent experiments due to the stronger transfection efficiency. Subse-quently, we found that the cell viability of sh-PDIA3P#2-transfected groups was significantly reduced compared with sh-NC transfected or control cells (P < 0.05, Figure 1D). Next, flow cytometry showed that the percentage of apop-totic cells was clearly increased after transfec-tion with sh-PDIA3P#2 relative to sh-NC (P < 0.01, Figure 1E). Moreover, the expression of apoptosis-related proteins also exhibited con-sistent changes after different transfection. Lastly, the transwell assay was performed to study the effects of PDIA3P suppression on cell migration and invasion. It was revealed that the number of migrated or invaded cells was dis-tinctly decreased following PDIA3P suppression (P < 0.05, Figure 1F, 1G). Taken together, these results suggested that suppression of PDIA3P decreased cell viability, promoted apoptosis, and inhibited migration and invasion.
PDIA3P regulates tumor growth and metasta-sis through negatively regulating miR-183
medi-Figure 1. LncRNA PDIA3P expression is upregulated in breast cancer (BC) tissues and cells. A: qRT-PCR showed the PDIA3P expression in BC tissues and adjacent
non-tumor tissues. B: qRT-PCR showed the PDIA3P expression in BC cells (MCF-7 and MDA-MB-231 cells) and normal mammary MCF10A cells. *P < 0.05 and **P < 0.01 compared to corresponding control. C-G. Suppression of PDIA3P in MCF-7 cells decreased cell viability, promoted apoptosis, and inhibited migration and
invasion. C: qRT-PCR showed the PDIA3P expression in MCF-7 cells after transfection with sh-NC, sh-PDIA3P#1 or sh-PDIA3P#2. D: MTT assay showed the MCF-7 cell viability after different transfections. E: Flow cytometry showed that the percentage of apoptotic MCF-7 cells after different transfections and western blot revealed the expression of apoptosis-related proteins after different transfections. F, G: Transwell assays showed the migration and invasion of MCF-7 cells after different
[image:4.792.95.705.69.389.2]ated by miR-183, the miR-183 was dysregulat-ed in MCF-7, followdysregulat-ed by detection of the effects of co-transfection of sh-PDIA3P#2 and miR-183 inhibitor in MCF-7 cells. As shown in Figure 2D, miR-183 expression was prominently over-expressed and suppressed in MCF-7 cells by transfection of miR-183 mimic and miR-183 inhibitor (P < 0.001), respectively. Subsequently, we found that cotransfection of sh-PDIA3P#2 and miR-183 inhibitor significantly reversed the effects of sh-PDIA3P#2 alone by enhancing cell viability (P < 0.05, Figure 2E), decreasing
apop-tosis (P < 0.05, Figure 2F), and prompting mi gration and invasion (P < 0.05, Figure 2G, 2H).
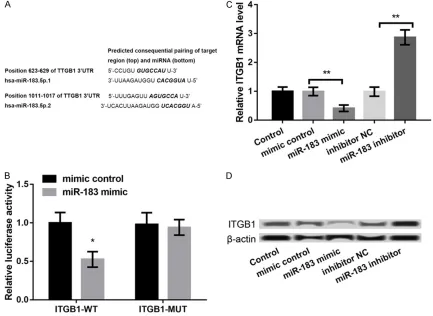
ITGB1 is a target of miR-183
[image:6.612.88.521.185.501.2]ITGB1 is one member of the integrins that are crucial in cancers [15], we predicted the inter-action between miR-183 and ITGB1 using TargetScan software (version 7.1). There was a complementary pairing relationship between miR-183 and ITGB1 (Figure 3A). Moreover, luciferase activity assay further confirmed that Figure 2. PDIA3P regulated tumor growth and metastasis through negative regulating miR-183. A: qRT-PCR showed the 183 expression in MCF-7 cells after transfection with sh-NC, or sh-PDIA3P#2. B: qRT-PCR showed the miR-183 expression in BC tissues and adjacent non-tumor tissues. C: qRT-PCR showed the PDIA3P expression in BC cells (MCF-7 and MDA-MB-231 cells) and normal mammary MCF10A cells. D: qRT-PCR showed the miR-183 expres-sion in MCF-7 cells after transfection with miR-183 mimic, mimic control, miR-183 inhibitor, and inhibitor control. E: MTT assay showed the MCF-7 cell viability after different transfections. F: Flow cytometry showed that the percent-age of apoptotic MCF-7 cells after different transfections and western bolt revealed the expression of apoptosis-related proteins after different transfections. G, H: Transwell assays showed the migration and invasion of MCF-7
cells after different transfections. *P < 0.05, **P < 0.01, and ***P < 0.001 compared to corresponding control.
Figure 3. ITGB1 was a target of miR-183. A: The complementary pairing relationship between miR-183 and ITGB1
according to the information of TargetScan software. B: Luciferase activity assay confirmed that miR-83 mimic in -hibited the luciferase activity of ITGB1-WT. C, D: qRT-PCR and western blot showed the ITGB1 expression in MCF-7
miR-83 mimic significantly inhibited the lucifer -ase activity of ITGB1-WT (P < 0.05), but showed no effects on ITGB1-MUT (Figure 3B). These data indicated that ITGB1 was a target of miR-183. Furthermore, the expression of ITGB1 at both mRNA and protein levels was markedly downregulated in miR-183 mimic group and dramatically upregulated in miR-183 inhibitor group (P < 0.05, Figure 3C and 3D), confirming that ITGB1 expression was negatively regulated by miR-183.
miR-183 regulates tumor growth and metasta-sis through targeting ITGB1
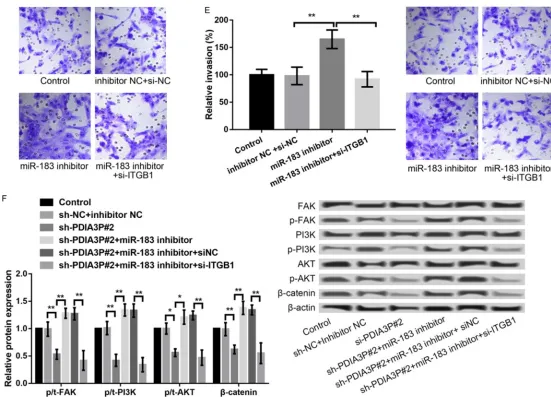
To verify the regulatory interaction between miR-183 and ITGB1 in BC cells, ITGB1 was sup-pressed in MCF-7 cells by transfection with si-ITGB1. The level of ITGB1 was significantly decreased in si-ITGB1-transfected cells in com-parison with that in si-NC-transfected cells (P < 0.01, Figure 4A). Moreover, in relation to trans-fection with miR-183 inhibitor alone, co-trans-fection with miR-183 inhibitor and si-ITGB1 in MCF-7 cells significantly decreased cell viability (P < 0.05, Figure 4B), promoted apoptosis (P < 0.001, Figure 4C) and suppressed migration (P < 0.0A, Figure 4D) and invasion (P < 0.01, Figure 4E). These data indicated that miR-183 regulates tumor growth and metastasis through targeting ITGB1.
PDIA3P mediates the process of BC cells through FAK/PI3K/AKT/β-catenin pathway
Previous evidence reported that ITGB1 is involved in the development of tumors includ-ing BC by FAK/PI3K/AKT/β-catenin pathway [16, 17], we investigated whether FAK/PI3K/ AKT/β-catenin signal was involved in the pro -cess of PDIA3P-mediated BCPDIA3P suppres-sion distinctly decreased the expressuppres-sion levels of p-FAK, p-PI3K, p-AKT and β-catenin, which was significantly reversed after simultaneous inhibition of miR-183 (Figure 4F). Moreover, the combined effects of simultaneous suppression of PDIA3P and inhibition of miR-183 on the expression of key FAK/PI3K/AKT/β-catenin-related proteins were reversed again after fur-ther suppression of ITGB1 at the same time (Figure 4F).
Discussion
The current research explored the roles of PDIA3P in the process of BC. We discovered
that PDIA3P was up-regulated in BC tissues and cells. Suppression of PDIA3P markedly decreased the cell viability, promoted apopto-sis, and inhibited migration and invasion. In addition, PDIA3P was negatively interacted with miR-183 in regulating tumor growth and metastasis. Moreover, miR-183 regulated tu- mor growth and metastasis through targeting ITGB1. Lastly, PDIA3P distinctly inhibited the activation of FAK/PI3K/AKT/β-catenin path -way, which was significantly reversed after simultaneous inhibition of miR-183 and further reversed after suppression of ITGB1 at the same time.
Increasing evidences have confirmed that lncRNAs can function as ceRNAs to modulate gene expression through sponging miRNAs, thus contributing to the development of many diseases [18, 19]. Our findings showed that PDIA3P modulated the expression of ITGB1 by competitively sponging 183. Besides, miR-183 is located on chromosome 7q32 and is dysregulated in numerous cancers [20, 21]. Lowery demonstrated that miR-183 overex-pression could suppress the cell migration [22], which was consistent with our findings. miR-183/-96/-182 cluster is crucial to the develop-ment of breast tumor [23]. Furthermore, ITGB1 was targeted by miR-183, and aberrant expres-sion of ITGB1 was also revealed in a variety of malignancies, such as prostate cancer [24], oral squamous cell carcinoma [25], and gastric cancer [26]. Moreover, a gene expression profil -ing combined with functional analysis has revealed that ITGB1 function as a promising prognostic biomarker for triple negative breast cancer [27]. The kinase LMTK3 promoted the invasion of BC cells by GRB2-mediated induc-tion of ITGB1 [28]. Given the key role of ITGB1 in BC, we speculated that PDIA3P may contrib-ute to BC development by modulating the expression of ITGB1 by competitively sponging miR-183.
Figure 4. miR-183 regulated tumor growth and metastasis through targeting ITGB1. A: qRT-PCR and western blot showed the ITGB1 expression in MCF-7 cells after transfection with si-NC or si-ITGB1. B: MTT assay showed the MCF-7 cell viability after different transfections. C: Flow cytometry showed that the percentage of apop-totic MCF-7 cells after different transfections and western bolt revealed the expression of apoptosis-related proteins after different transfections. D, E: Transwell
the combined effects of simultaneous suppres-sion of PDIA3P and inhibition of miR-183 on the expression of key FAK/PI3K/AKT/β-catenin-related proteins were reversed again after fur-ther suppression of ITGB1 at the same time. Taken together, our results reflected that PDIA3P may promote BC development by regu-lating the activation of ITGB1/FAK/PI3K/AKT/ β-catenin signals.
To sum up, our findings reflected that PDIA3P may contribute to the process of BC by spong-ing miR-183/ITGB1 through the activation of FAK/PI3K/AKT/β-catenin signals. PDIA3P may serve as a promising biomarker or targets for the detection or treatment of BC.
Acknowledgements
This work was supported by National Natural Science Foundation of China, Grant No: 81472454 and 81572082; Bethune program of Jilin University, Grant No: 2015318, Project of Science and Technology of Jilin Province, Nos 2150414015GH.
Disclosure of conflict of interest
None.
Address correspondence to: Ling Fang, Department of Breast and Thyroid Surgery, China-Japan Union Hospital of Jilin University, 126 Xiantai Avenue, Changchun 130033, Jilin, China. E-mail: lingfang- [email protected]
References
[1] Siegel R, Ma J, Zou Z and Jemal A. Cancer sta-tistics, 2014. CA Cancer J Clin 2014; 64: 104-17.
[2] Sidaway P. Breast cancer: LAG3 expression
in-dicates favourable outcomes. Nat Rev Clin On -col 2017; 14: 712.
[3] Bussard KM and Marini FC. Tumor-associated
osteoblasts are major mediators in facilitating bone metastatic breast cancer cell quies-cence. Cancer Res 2016; 76: 1556.
[4] Gutschner T and Diederichs S. The hallmarks of cancer: a long non-coding RNA point of view. RNA Biol 2012; 9: 703-19.
[5] Fatica A and Bozzoni I. Long non-coding RNAs: new players in cell differentiation and develop-ment. Nat Rev Genet 2014; 15: 7.
[6] Chandra Gupta S and Nandan Tripathi Y. Po-tential of long non-coding RNAs in cancer pa-tients: from biomarkers to therapeutic targets. Int J Cancer 2017; 140: 1955-1967.
[7] Bhan A, Soleimani M and Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res 2017; 77: 3965-3981.
[8] Qi P and Du X. The long non-coding RNAs, a
new cancer diagnostic and therapeutic gold mine. Mod Pathol 2013; 26: 155-65.
[9] Niknafs YS, Han S, Ma T, Speers C, Zhang C,
Wilder-Romans K, Iyer MK, Pitchiaya S, Malik R
and Hosono Y. The lncRNA landscape of breast cancer reveals a role for DSCAM-AS1 in breast cancer progression. Nat Commun 2016; 7: 12791.
[10] Sas-Chen A, Aure MR, Leibovich L, Carvalho S,
Enuka Y, Körner C, Polycarpou-Schwarz M, Lavi S, Nevo N, Kuznetsov Y, Yuan J, Azuaje F; Oslo Breast Cancer Research Consortium (OSBRE -AC)SauerTorillGeislerJürgenHofvindSolveig-
BathenTone FBorgenElinEngebråtenOlavFod
-stadØysteinGarredØysteinGeitvikGry
AarumK-åresenRolfNaumeBjørnMælandsmoGunhild MariRussnesHege GSchlichtingEllenSørlieTh-
ereseLingjærdeOle ChristianSahlbergKristine KleiviSkjervenHelle KristineFritzmanBritt, Ulit-sky I, Diederichs S, Wiemann S, Yakhini Z, Kris -tensen VN, Børresen-Dale AL, Yarden Y. LIMT is a novel metastasis inhibiting lncRNA sup-pressed by EGF and downregulated in
aggres-sive breast cancer. EMBO Mol Med 2016; 8:
1052-64.
[11] Pickard MR and Williams GT. The hormone re-sponse element mimic sequence of GAS5
ln-cRNA is sufficient to induce apoptosis in breast cancer cells. Oncotarget 2016; 7: 10104.
[12] Heilmann K, Toth R, Bossmann C, Klimo K,
Plass C and Gerhauser C. Genome-wide screen for differentially methylated long noncoding
RNAs identifies Esrp2 and lncRNA Esrp2-as
regulated by enhancer DNA methylation with prognostic relevance for human breast cancer.
Oncogene 2017; 36: 6446.
[13] Sun CC, Zhang L, Li G, Li SJ, Chen ZL, Fu YF,
Gong FY, Bai T, Zhang DY and Wu QM. The ln -cRNA PDIA3P interacts with miR-185-5p to modulate oral squamous cell carcinoma pro-gression by targeting cyclin D2. Mol Ther Nu-cleic Acids 2017; 9: 100-110.
[14] Yang X, Ye H, He M, Zhou X, Sun N, Guo W, Lin X, Huang H, Lin Y and Yao R. LncRNA PDIA3P interacts with c-Myc to regulate cell prolifera-tion via inducprolifera-tion of pentose phosphate path-way in multiple myeloma. Biochem Biophys Res Commun 2018; 498: 207-213.
[15] Desgrosellier JS and Cheresh DA. Integrins in cancer: biological implications and therapeutic opportunities. Nat Rev Cancer 2010; 10: 9-22. [16] Zeng B, Zhou M, Wu H and Xiong Z. sPP1 pro-motes ovarian cancer progression via integrin
β1/FaK/aKT signaling pathway. Onco Targets
[17] Qin J, Tang J, Jiao L, Ji J, Chen WD, Feng GK,
Gao YH, Zhu XF and Deng R. A diterpenoid compound, excisanin A, inhibits the invasive behavior of breast cancer cells by modulating
the integrin β1/FAK/PI3K/AKT/β-catenin sig -naling. Life Sci 2013; 93: 655-663.
[18] Jiao C, Song Z, Chen J, Zhong J, Cai W, Tian S, Chen S, Yi Y and Xiao Y. lncRNA-UCA1 enhanc-es cell proliferation through functioning as a
ceRNA of Sox4 in esophageal cancer. Oncol
Rep 2016; 36: 2960-2966.
[19] Zhang XF, Ye Y and Zhao SJ. LncRNA Gas5 acts as a ceRNA to regulate PTEN expression by sponging miR-222-3p in papillary thyroid
carci-noma. Oncotarget 2018; 9: 3519-3530.
[20] Yamada Y, Enokida H, Kojima S, Kawakami K, Chiyomaru T, Tatarano S, Yoshino H, Kawahara K, Nishiyama K and Seki N. MiR-96 and
miR-183 detection in urine serve as potential tu-mor markers of urothelial carcinoma: correla-tion with stage and grade, and comparison with urinary cytology. Cancer Sci 2011; 102: 522-9.
[21] Li J, Fu H, Xu C, Tie Y, Xing R, Zhu J, Qin Y, Sun Z and Zheng X. miR-183 inhibits
TGF-β1-induced apoptosis by downregulation of PDCD4 expression in human hepatocellular carcinoma cells. BMC Cancer 2010; 10: 354. [22] Lowery AJ, Miller N, Dwyer RM and Kerin MJ.
Dysregulated miR-183 inhibits migration in breast cancer cells. BMC Cancer 2010; 10: 502.
[23] Li P, Sheng C, Huang L, Zhang H, Huang L,
Cheng Z and Zhu Q. MiR-183/-96/-182 cluster
is up-regulated in most breast cancers and in-creases cell proliferation and migration. Breast Cancer Res 2014; 16: 473.
[24] Kurozumi A, Goto Y, Matsushita R, Fukumoto I, Kato M, Nishikawa R, Sakamoto S, Enokida H,
Nakagawa M and Ichikawa T. Tumor-suppres-sive micro RNA-223 inhibits cancer cell migra-tion and invasion by targeting ITGA 3/ITGB 1 signaling in prostate cancer. Cancer Sci 2016; 107: 84-94.
[25] Hunt S, Jones AV, Hinsley EE, Whawell SA and Lambert DW. MicroRNA-124 suppresses oral squamous cell carcinoma motility by targeting ITGB1. FEBS Lett 2011; 585: 187-92.
[26] Han TS, Hur K, Xu G, Choi B, Okugawa Y, Toi
-yama Y, Oshima H, Oshima M, Lee HJ and Kim
VN. MicroRNA-29c mediates initiation of gas-tric carcinogenesis by directly targeting ITGB1. Gut 2015; 64: 203-214.
[27] Klahan S, Huang WC, Chang CM, Wong HS,
Huang CC, Wu MS, Lin YC, Lu HF, Hou MF,
Chang WC. Gene expression profiling com -bined with functional analysis identify integrin beta1 (ITGB1) as a potential prognosis bio-marker in triple negative breast cancer. Phar-macol Res 2016; 104: 31-7.
[28] Xu Y, Zhang H, Lit LC, Grothey A, Athanasiadou
M, Kiritsi M, Lombardo Y, Frampton AE, Green AR, Ellis IO, Ali S, Lenz HJ, Thanou M, Stebbing J, Giamas G. The kinase LMTK3 promotes inva -sion in breast cancer through GRB2-mediated
induction of integrin β1. Sci Signal 2014; 7:
ra58.
[29] Yeh YC, Lin HH and Tang MJ. A tale of two
col-lagen receptors, integrin β1 and discoidin do -main receptor 1, in epithelial cell differentia-tion. Am J Physiol Cell Physiol 2012; 303: C1207-17.
[30] Yang J, Hou Y, Zhou M, Wen S, Zhou J, Xu L, Tang X, Du YE, Hu P, Liu M. Twist induces epi-thelial-mesenchymal transition and cell
motili-ty in breast cancer via ITGB1-FAK/ILK signaling