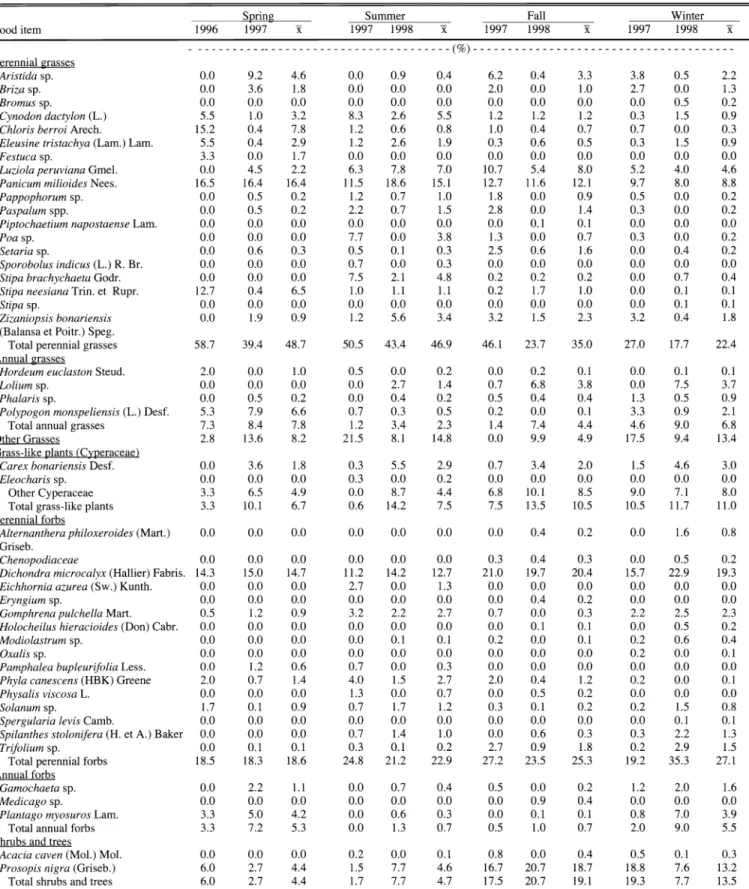
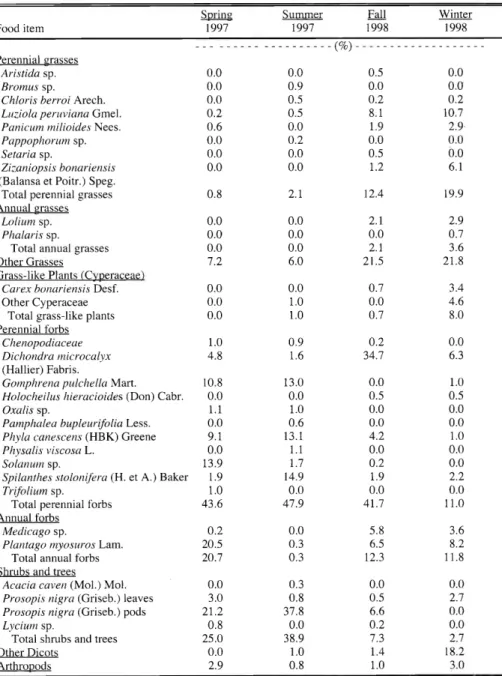
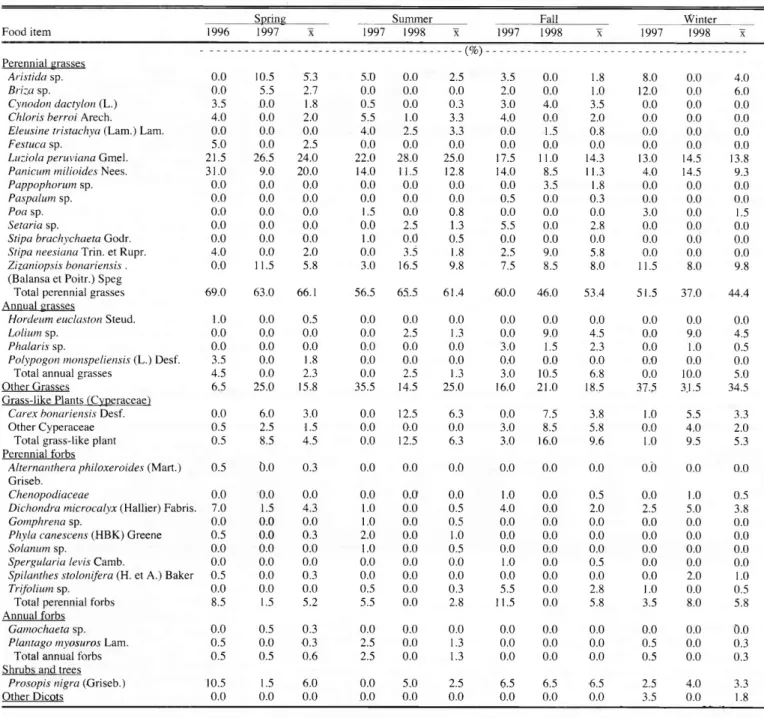
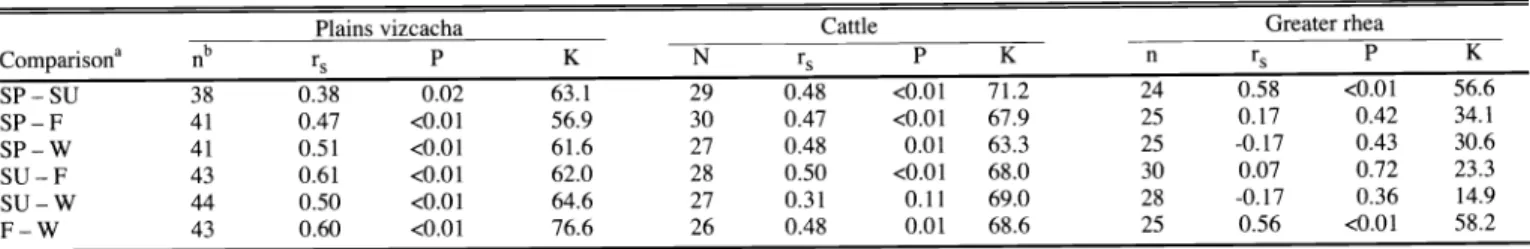
cattle plains
8
0
0
Full text
(2)
(3)
(4)
(5)
(6)
(7)
(8)
Figure
Related documents
Some prior studies have proposed using P2P approaches for the signaling phase of VoIP communication. However, a P2P approach may increase the call setup time, an important
Survey: The final renders will be presented to a number of participants as a short video sequence, containing three different simulations – Smoke, Fire and Water – from both Maya
It argues that a feminist research provides a powerful tool in assisting us to understand women’s lives, their experiences, their concerns based on
Our research conducted a data mining project on direct marketing campaigns for deposits subscriptions by using recent data of a Portuguese retail bank.. We used the Support
not provided to respondents (can be corrected by using “other” response option).. Survey Construction: Question Types.
The yearly aggregated tax income data of all, more than 8000, Italian municipalities are analyzed for a period of five years, from 2007 to 2011, to search for conformity or not
The mentoring for leadership empowerment and mentoring for personal development composite variables were both positively correlated with the dependent variable and also
Table 3-22 The effective refractive index and effective radius at three different RH conditions, and the derived growth factor (gf), κ(RH), and WUP for azelaic acid *(AzA)
