The heterodonty in euselachian sharks from the Pennsylvanian of Nebraska
Full text
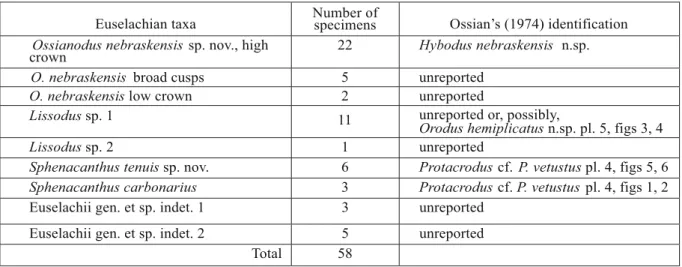
Figure
Related documents
who is previously considered as a non-resident and who arrives in the Philippines at anytime during the taxable year to reside thereat permanently shall be
organisasjonslæring, arbeidsplasslæring, uformell og formell læring, læring gjennom praksis, sosial praksis og så videre vil derfor være nyttige når man skal foreta en studie
We are not able to transfer the holding to you but, if in the unlikely event that a future payment is received, we will pass it on to you, provided we know your current address..
Figure: Activities of Bristol Food Network in the food supply chain Farmers and growers in and around Bristol Food processors, packers, wholesalers, distributors Direct sales,
• Follow up with your employer each reporting period to ensure your hours are reported on a regular basis?. • Discuss your progress with
Proprietary Schools are referred to as those classified nonpublic, which sell or offer for sale mostly post- secondary instruction which leads to an occupation..
Ravno tako smo potrdili drugo tezo: »Nevronski model za napovedovanje bo podal dovolj dobre rezultate, ki lahko pomembno pripomorejo k sprejemanju pravih odločitev.« Glede na
4.1 The Select Committee is asked to consider the proposed development of the Customer Service Function, the recommended service delivery option and the investment required8. It
