0095-1137/09/$08.00⫹0 doi:10.1128/JCM.00910-09
Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Comparison of Fingerprinting Methods for Typing
Methicillin-Resistant
Staphylococcus aureus
Sequence Type 398
䌤
G. Rasschaert,
1* W. Vanderhaeghen,
2I. Dewaele,
1N. Janez
ˇ,
1X. Huijsdens,
3P. Butaye,
2,4and M. Heyndrickx
1Institute for Agricultural and Fisheries Research (ILVO), Technology and Food Science Unit, Brusselsesteenweg 370, 9090 Melle,
Belgium1; Veterinary and Agrochemical Research Centre, Groeselenberg 99, 1180 Brussels, Belgium2; National Institute of
Public Health and the Environment (RIVM), P.O. Box 1, 3720 BA Bilthoven, The Netherlands3; and Ghent University,
Faculty of Veterinary Medicine, Department of Bacteriology, Pathology and Poultry Diseases,
Salisburylaan 133, 9820 Merelbeke, Belgium4
Received 8 May 2009/Returned for modification 23 July 2009/Accepted 17 August 2009
This study evaluates the multiple-locus variable-number tandem-repeat assay (MLVA) and pulsed-field gel electrophoresis (PFGE) when using restriction enzymes BstZI, SacII, and ApaI to fingerprint a diverse collection of methicillin (meticillin)-resistantStaphylococcus aureus(MRSA) sequence type 398 (ST398) iso-lates. These isolates had been characterized previously by multilocus sequence typing,spatyping, and staph-ylococcal cassette chromosomemec(SCCmec) typing. Typeability and discriminatory power were analyzed, and the concordance between the various methods was determined. All MRSA ST398 isolates were typeable by the MLVA and PFGE using BstZI, SacII, and ApaI. With each method, the MRSA ST398 isolates formed a separate group from the two non-ST398 MRSA strains. PFGE, performed with any of the three restriction enzymes, had the most discriminatory power, followed by MLVA,spatyping, and SCCmectyping. The MLVA showed the highest concordance with PFGE using ApaI andspatyping. As further expressed by the Wallace coefficient, the MLVA type was poorly predicted byspa typing, whereas thespa type was well predicted by MLVA. PFGE, using a combination of all three restriction enzymes, had the highest concordance with the MLVA but had a low probability of being predicted by MLVA. PFGE, using a combination of all three restriction enzymes, was able to predict SCCmectype and MLVA type completely and had a high probability of predictingspa type. Both the MLVA and PFGE could be used to discriminate among the MRSA ST398 isolates. Although the MLVA is a faster technique, PFGE had more discriminatory power than the MLVA, especially when a combination of restriction enzymes was used.
Infections caused by methicillin (meticillin)-resistant
Staph-ylococcus aureus (MRSA) have been a problem in hospitals
and nursing homes for many decades. These MRSA isolates are therefore called health care-associated MRSA (HA-MRSA) isolates (1). Since the early 1990s, MRSA has emerged in healthy persons without risk factors for MRSA infections. These isolates are the so-called community-associated MRSA (CA-MRSA) isolates (20). In the last few years, MRSA has been isolated from livestock animals (pigs in particular) and pig farmers (5, 6, 30). These MRSA strains are called animal-associ-ated MRSA (AA-MRSA) strains. It seems that (livestock) ani-mals form a new, separate reservoir. These AA-MRSA strains all appear to belong to the new clonal complex 398 (CC398), with sequence type 398 (ST398) as the basic type, as determined by multilocus sequence typing (MLST) (29). MRSA ST398 has al-ready been isolated in Europe, Asia, and North America (32). Considering the worldwide spread of MRSA, epidemiological questions arise about its transmission within farms, among farms, and from farms to the population. Fast and inexpensive typing
methods with good discriminatory power are necessary to con-duct large-scale epidemiological studies.
Traditionally, human MRSA isolates have been typed by pulsed-field gel electrophoresis (PFGE), using SmaI as the re-striction enzyme (19). The advantages of using PFGE are good discriminatory power and good reproducibility at the interlabo-ratory level when standardized protocols are used. However, AA-MRSA is not typeable by this method, as the activity of SmaI is blocked due to methylation of the restriction site (2).
More recently, methods based on DNA sequencing, such as MLST andspatyping, are increasingly being used to discrim-inate among different MRSA strains. Given their excellent interlaboratory reproducibility, online databases have been made to collate and harmonize data from various geographic regions. The drawback of MLST, which measures sequence variation at seven housekeeping loci, is its limited use with epidemiological studies due to its weak discriminatory power, time-consuming protocols, and high costs.spatyping, based on the variation in repeats present in the X-region of staphylo-coccal protein A, has a discriminatory power that lies between those of PFGE and MLST. Within ST398, severalspatypes have been distinguished, although the number of spa types seems rather limited in most countries.
One promising method is the multiple-locus variable-num-ber tandem-repeat assay (MLVA), a PCR-based method, * Corresponding author. Mailing address: Institute for Agricultural
and Fisheries Research (ILVO), Technology and Food Science Unit, Brusselsesteenweg 370, 9090 Melle, Belgium. Phone: 32 9 272 30 26. Fax: 32 9 272 30 01. E-mail: [email protected].
䌤Published ahead of print on 26 August 2009.
3313
on May 16, 2020 by guest
http://jcm.asm.org/
based on the analysis of the number of repeats in the variable-number tandem-repeat regions of various individual genes. This method has proven to be useful for typing both
Staphy-lococcus aureusand clinical MRSA isolates with good
repro-ducibility and good discriminatory power. Because the MLVA is also simple, inexpensive, and easy to interpret, it is useful as a typing method for large-scale epidemiological studies (10, 11, 12, 15, 16, 17, 23, 24, 27).
This study aimed to investigate various methods for typing MRSA ST398 isolates. An MLVA, consisting of a selection of primers from three existing MLVA systems, was tested with a collection of MRSA ST398 isolates. In addition, PFGE with restriction enzymes other than SmaI was performed with this set of isolates. These isolates had been previously character-ized by MLST,spa typing, and staphylococcal cassette chro-mosomemec(SCCmec) typing. Typeability and discriminatory power were analyzed for all methods, and the concordance among the different methods was determined.
MATERIALS AND METHODS
Selection of the strains.This study used 34 MRSA strains (Table 1). MLST typing revealed that 32 isolates belong to MRSA ST398 (8; http://saureus.mlst
.net). The isolates had variousspatypes (www.ridom.de/staphtype), SCCmec
types, and origins (18, 21, 33). The strains were isolated from humans who were in contact with pigs, chickens, cows, and the farm environment. Twenty-one strains were isolated in Belgium during studies performed in 2007 and 2008 (7, 22, 31). Another 11 strains originated in The Netherlands (5, 28). The strains ATCC 33592 and ATCC 43300 were included as non-ST398 MRSA reference strains.
MLVA.Chromosomal DNA was prepared by suspending a few colonies from a tryptic soy agar plate in water containing 10% lysostaphin (1 mg/ml). After
incubation for 10 min at 37°C and centrifugation for 2 min at 14,000⫻g, DNA
was prepared further according the protocol of Flamm et al. (9).
First, the primer pairs listed in Table 2 were used separately in order to evaluate the typeability and discriminating power of each primer pair on the
collection of 34 MRSA isolates. Each PCR mixture contained 1⫻PCR buffer,
1.3 mM MgCl2, 2.5 U AmpliTaq DNA polymerase (Applied Biosystems, Foster
City, CA), 200M of each deoxynucleoside triphosphate, 1.5M of a primer
pair, and 2l of template DNA (25 ng/l) in a final volume of 25l. PCR was
performed with predenaturation at 94°C for 5 min, followed by 25 cycles of 30 s at 94°C, 30 s at 58°C, and 60 s at 72°C, with a final extension at 72°C for 5 min on a GeneAmp PCR system 9700 (Applied Biosystems, Foster City, CA). Ten microliters of PCR product was used for electrophoresis in 2% SeaKem LE
agarose with 1⫻Tris-borate-EDTA (TBE; pH 8) for 60 min at 100 V.
Only primers that discriminated among the 32 MRSA ST398 isolates were included in the final MRSA ST398 MLVA. This multiplex PCR was performed under the same conditions described above. Eighteen microliters of PCR
[image:2.585.47.548.81.431.2]prod-uct was electrophoresed in 1.5% SeaKem LE agarose with 1⫻TBE (pH 8) for
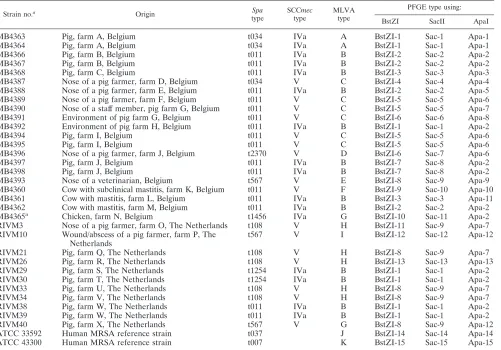
TABLE 1. MRSA ST398 isolates used in the present study grouped according their origin and types defined by the MLVA and PFGE
Strain no.a
Origin Spa
type
SCCmec
type
MLVA type
PFGE type using:
BstZI SacII ApaI
MB4363 Pig, farm A, Belgium t034 IVa A BstZI-1 Sac-1 Apa-1
MB4364 Pig, farm A, Belgium t034 IVa A BstZI-1 Sac-1 Apa-1
MB4366 Pig, farm B, Belgium t011 IVa B BstZI-2 Sac-2 Apa-2
MB4367 Pig, farm B, Belgium t011 IVa B BstZI-2 Sac-2 Apa-2
MB4368 Pig, farm C, Belgium t011 IVa B BstZI-3 Sac-3 Apa-3
MB4387 Nose of a pig farmer, farm D, Belgium t034 V C BstZI-4 Sac-4 Apa-4
MB4388 Nose of a pig farmer, farm E, Belgium t011 IVa B BstZI-2 Sac-2 Apa-5
MB4389 Nose of a pig farmer, farm F, Belgium t011 V C BstZI-5 Sac-5 Apa-6
MB4390 Nose of a staff member, pig farm G, Belgium t011 V C BstZI-5 Sac-5 Apa-7
MB4391 Environment of pig farm G, Belgium t011 V C BstZI-6 Sac-6 Apa-8
MB4392 Environment of pig farm H, Belgium t011 IVa B BstZI-1 Sac-1 Apa-2
MB4394 Pig, farm I, Belgium t011 V C BstZI-5 Sac-5 Apa-6
MB4395 Pig, farm I, Belgium t011 V C BstZI-5 Sac-5 Apa-6
MB4396 Nose of a pig farmer, farm J, Belgium t2370 V D BstZI-6 Sac-7 Apa-6
MB4397 Pig, farm J, Belgium t011 IVa B BstZI-7 Sac-8 Apa-2
MB4398 Pig, farm J, Belgium t011 IVa B BstZI-7 Sac-8 Apa-2
MB4393 Nose of a veterinarian, Belgium t567 V E BstZI-8 Sac-9 Apa-9
MB4360 Cow with subclinical mastitis, farm K, Belgium t011 V F BstZI-9 Sac-10 Apa-10
MB4361 Cow with mastitis, farm L, Belgium t011 IVa B BstZI-3 Sac-3 Apa-11
MB4362 Cow with mastitis, farm M, Belgium t011 IVa B BstZI-2 Sac-2 Apa-2
MB4365a Chicken, farm N, Belgium t1456 IVa G BstZI-10 Sac-11 Apa-2
RIVM3 Nose of a pig farmer, farm O, The Netherlands t108 V H BstZI-11 Sac-9 Apa-7 RIVM10 Wound/abscess of a pig farmer, farm P, The
Netherlands
t567 V I BstZI-12 Sac-12 Apa-12
RIVM21 Pig, farm Q, The Netherlands t108 V H BstZI-8 Sac-9 Apa-7
RIVM26 Pig, farm R, The Netherlands t108 V H BstZI-13 Sac-13 Apa-13
RIVM29 Pig, farm S, The Netherlands t1254 IVa B BstZI-1 Sac-1 Apa-2
RIVM30 Pig, farm T, The Netherlands t1254 IVa B BstZI-1 Sac-1 Apa-2
RIVM33 Pig, farm U, The Netherlands t108 V H BstZI-8 Sac-9 Apa-7
RIVM34 Pig, farm V, The Netherlands t108 V H BstZI-8 Sac-9 Apa-7
RIVM38 Pig, farm W, The Netherlands t011 IVa B BstZI-1 Sac-1 Apa-2
RIVM39 Pig, farm W, The Netherlands t011 IVa B BstZI-1 Sac-1 Apa-2
RIVM40 Pig, farm X, The Netherlands t567 V G BstZI-8 Sac-9 Apa-12
ATCC 33592 Human MRSA reference strain t037 J BstZI-14 Sac-14 Apa-14
ATCC 43300 Human MRSA reference strain t007 K BstZI-15 Sac-15 Apa-15
aMB, Molecular Bacteriology strain collection at ILVO-T&V laboratories (MB4365 originates from the work of Persoons et al.关22兴; MB4387, MB4388, and MB4389
were provided by the Department of Microbiology, Université Libre de Bruxelles, Hoˆpital Erasme, Belgium); RIVM, National Institute for Public Health and the
Environment; ATCC, American Type Culture Collection.
on May 16, 2020 by guest
http://jcm.asm.org/
240 min at 140 V. The DNA size standard was a 100-bp DNA ladder (Invitrogen, Carlsbad, CA). The gels were stained in ethidium bromide, visualized on an UV transilluminator, and digitally photographed. Gel patterns were analyzed with BioNumerics version 5.10 (Applied Maths, Sint-Martens-Latem, Belgium) and clustered using Dice’s coefficient (tolerance, 1%; optimization, 0.1% to 1%) and the unweighted-pair group method using average linkages (UPGMA).
PFGE.The plugs were prepared according the protocol of Struelens et al. (26), with modifications. Briefly, an overnight-grown culture was suspended in EET buffer (100 mM EDTA, 10 mM EGTA, 10 mM Tris-HCl [pH 8]) and adjusted to an optical density at 600 nm value of 0.9. The suspension was mixed with equal volumes of a 2% solution of low-melting-temperature agarose (InCert agarose; Cambrex Bio Science Rockland, Rockland, MA) in EET buffer. After solidifi-cation, the agarose plugs were incubated for 4 h at 37°C in EET buffer containing
1 mg lysozyme and 50g lysostaphin per ml. The plugs were transferred into
EET buffer containing 1% sodium dodecyl sulfate and 1 mg proteinase K per ml of buffer and incubated overnight at 50°C. Afterwards, the plugs were washed thoroughly with TE buffer (10 mM Tris-HCl, 1 mM EDTA [pH 8]).
The restriction enzyme chosen was isoschizomer XmaI, because it is a meth-ylation-insensitive version of SmaI. XmaI cuts the same recognition sequence as SmaI, but at a different position. The other restriction enzymes were chosen based on the restriction digest and PFGE program provided at http://insilico.ehu .es/ (3). DNA was digested overnight with 40 U SmaI (Invitrogen, Carlsbad, CA), SacII, XmaI (New England Biolabs, Ipswich, MA), ApaI, or BstZI (Promega, Madison, WI) at the recommended assay temperatures. DNA fragments were separated on a CHEF Mapper system (Bio-Rad Laboratories, Hemel Hemp-stead, United Kingdom) in a 1% SeaKem gold agarose gel (Lonza, Rockland,
MA). The running conditions were 6 V/cm in 0.5⫻TBE (45 mM Tris, 45 mM
boric acid, 1 mM EDTA [pH 8]) at 14°C. For separation of SmaI and XmaI DNA fragments, runs lasted 18 h, with switch times from 2.63 s to 63.8 s. ApaI fragments were separated during runs of 16 h, with switch times of 0.22 s to 17.33 s. DNA fragments digested with BstZI and SacII were separated during a run of 20 h, with an initial switch time of 0.46 s and a final switch time of 35.38 s for BstZI and a switch time of 44.69 s for SacII. Gels were stained with ethidium bromide, destained in water, and digitally captured under UV light. Gel images
were visually analyzed with BioNumerics version 5.10 (Applied Maths,
Sint-Martens-Latem, Belgium) usingSalmonellaBraenderup H9812 digested with
XbaI as a normalization reference. The similarity between the fingerprints was calculated using the Dice coefficient (with an optimization of 1% and a position tolerance of 0.5% to 1%), and they were grouped together according to their similarities using UPGMA.
Delineation of MLVA types and pulsotypes.An MLVA type was assigned on the basis of a difference of at least one band. As this method relies on the number of the tandem repeats in the variable-number tandem-repeat regions of different individual genes, even a small shift in the position of one band was considered to show another MLVA type. MLVA types are indicated by capital letters, as shown below.
For each restriction enzyme, a corresponding pulsotype was assigned, which was based on the difference in the presence or absence of at least one band. This was indicated by the name of the restriction enzyme followed by a number (e.g., BstZI-1).
Calculation of discriminatory power and concordance.The discriminatory power (D) was calculated according to the formula described by Hunter and Gaston (13, 14). The concordance of different typing techniques was calculated
with the software described by Carric¸o et al. (4). Rand’s index is an index that
shows the proportion of agreement, whereas the adjusted Rand’s index (R) shows the proportion of agreement corrected for the presence of chance agree-ment. Wallace’s coefficient (W) indicates the probability that two strains classi-fied as the same type by one method will also be classiclassi-fied as the same one when using the other method.
For these calculations, isolates originating from the same farm and having the
samespatype, SCCmectype, MLVA type, and pulsotype were considered to be
the same strain and were excluded from further analysis.
RESULTS
[image:3.585.44.545.80.380.2]Optimizing the MLVA.All primer pairs generated one band for at least one of the two reference strains, except the primers TABLE 2. Primers tested for discriminatory power with the MLVA
Locus Primer Primer sequence (5⬘–3⬘) Reference No. of
types
Discriminatory power (DI)
clfA clfA-F GATTCTGACCCAGGTTCAGA 23 3 0.65
clfA-R CTGTATCTGGTAATGGTTCTTT
clfB clfB-F ATGGTGATTCAGCAGTAAATCC 23 5 0.49
clfB-R CATTATTTGGTGGTGTAACTCTT
spa spa-F AGCACCAAAAGAGGAAGACAA 23 6 0.59
spa-R GTTTAACGACATGTACTCCGT
ssp sspA-F ATCMATTTYGCMAAYGATGACCA 23 1 0
sspA-R TTGTCTGAATTATTGTTATCGCC
sdr sdrCDE-F GTAACAATTACGGATCATGATG 23 —a —
sdrCDE-R TACCTGTTTCTGGTAATGCTTT
sdr sdrCDE2-F TTACGGATCATGATGATTTCA 10 5 0.50
sdrCDE2-R CAYTACCTGTTTCTGGTAATGCTT
fnbAB fnBP-F GGTCAAGCRCAAGGAACCART 10 1 0
fnBP-R AATAATCCGCCGAACAACAT
cna cna-F AAAATGACAAAATGGCAAG 10 1 0
cna-R CAGGTTTAGTTGGTGGTGTT
SIRU01 SIRU01-F CATGAGCAGTGCCTCCTTTA 11 1 0
SIRU01-R CGCCTTGTATCTTAAATTTGTTG
SIRU05 SIRU05-F AAAGCGTAAACACTCCGAACA 11 0 0
SIRU05-R CATCGGGTTCACTTCTTTCTT
SIRU07 SIRU07-F CGCAAAATTTATTTCGTCGT 11 1 0
SIRU07-R TTGGCATCATTTAGCAGACG
SIRU13 SIRU13-F GGGGGAAATTCTAAGCAACC 11 1 0
SIRU13-R CGAAATTTTCCACGTCGATT
SIRU15 SIRU15-F AATTATAGCATATTAGAGCCCCTTA 11 1 0
SIRU15-R ACGTAAAGGTCGCGACAAAA
SIRU16 SIRU16-F TCAGCTTGTGACTCTTCTTTCG 11 0 0
SIRU16-R AAGAACGTGTTGAAGGATTTTT
SIRU21 SIRU21-F AGCAGTAGTGCCGTTTGCTT 11 6 0.59
SIRU21-R GCTCAAGCACCAAAAGAGGA
a—, for this primer pair, nonspecific bands were obtained.
on May 16, 2020 by guest
http://jcm.asm.org/
forsdr, which generated multiple bands. Data for the number of types for the 32 MRSA ST398 isolates and the discrimina-tory power of each primer pair are given in Table 2. The primer pairs SIRU5 and SIRU16 generated no amplicons for the MRSA ST398 isolates. Primer pairs sspA, fnbAB, cna, SIRU1, SIRU7, SIRU13, and SIRU15 did not discriminate among the MRSA ST398 isolates. The ClfA and ClfB primer pairs gen-erated three and five different amplicons, respectively, al-though the difference in size forclfAwas minimal. Both primer pairs spa and SIRU21 had good discriminating power: both found six types among the tested MRSA ST398 isolates. For each isolate, primer pair SIRU21 consistently generated a band that was approximately 50 bp bigger than the bands generated by thespaprimers. The locus of SIRU21 has also been identified asspa(12). Last to be evaluated were the two primer pairs sdrCDE and sdrCDE2, used to detect the
poly-morphism of three individual genes, namelysdrC,sdrD, andsdrE.
Staphylococcus aureusstrains do not always possess all three of
these genes (25). The 32 MRSA ST398 strains possess only the
sdrC and sdrE genes, as tested in this study according to the protocol of Sabat et al. (25) (data not shown). Given this infor-mation, only two amplicons for each isolate can be expected when using these two primer pairs. Only the sdrCDE2 primer pair generated two bands for each MRSA ST398 isolate, and five different types were obtained.
[image:4.585.44.541.70.494.2]The following primer pairs were chosen for their discrimi-natory power and included in the ST398 MLVA: SIRU21, ClfA, ClfB, and sdrCDE2. The best results were obtained by running the sdrCDE2 primer set separately from the other three primer sets and mixing 10 l of the sdrCDE2 PCR product with 8l of the multiplex PCR product before elec-trophoresis.
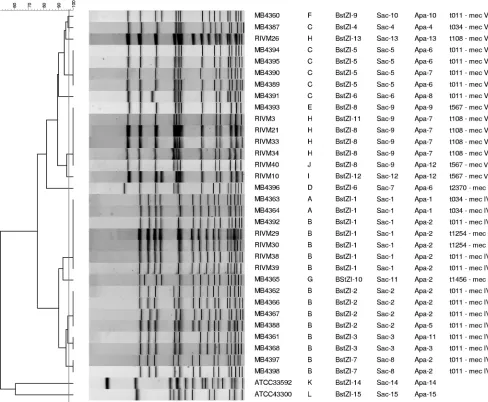
FIG. 1. Dendrogram and fingerprints for the MLVA with 32 MRSA ST398 isolates and two reference MRSA strains. The similarities between the fingerprints were calculated using the Dice coefficient (optimization, 1%; position tolerance, 0.1% to 1%), and the fingerprints were grouped according to their similarities by use of the UPGMA algorithm.
on May 16, 2020 by guest
http://jcm.asm.org/
Analysis of the MRSA ST398 strains using the MLVA.All 34 isolates were typeable using the MLVA described above. The MRSA ST398 isolates clustered together with a similarity of 33%. As some differences were so small, it was not possible to determine a cutoff value in order to automatically cate-gorize the types with the BioNumerics software. For exam-ple, MB4393 and RIVM10 were 100% related according to the BioNumerics software, but it was possible to visually distin-guish a small shift due to a smallerclfB band. Using visual categorization, 10 different MLVA types were obtained within the ST398 isolates (Fig. 1). Six MLVA types consisted of only one isolate. Two isolates belonged to MLVA type A, but these two isolates originated from the same farm. MLVA type H consisted of five isolates, all isolated in The Netherlands, whereas MLVA types B and C consisted of 13 and 6 isolates, respectively. These two last types were very similar; the only difference was a small shift of theclfAband.
Analysis of the MRSA ST398 isolates using PFGE.None of the AA-MRSA isolates was typeable by SmaI digestion, which confirmed this characteristic as being typical for MRSA CC398. However, they were all typeable using the restriction
enzymes BstZI, SacII, and ApaI. XmaI, an alternative to SmaI, as mentioned above, was also evaluated. However, the finger-prints obtained yielded several weak bands, even after optimi-zation of the restriction conditions. Because of this, XmaI was excluded from the study.
A cutoff value of 97% for delineation of the different pul-sotypes was determined for each restriction enzyme, according to the criteria of the delineation of pulsotypes as described above in Materials and Methods.
The fingerprints of the MRSA ST398 isolates obtained from PFGE using restriction enzyme BstZI clustered together with a similarity of 79%, whereas the two reference non-ST398 MRSA isolates had a similarity with the MRSA ST398 isolates of less than 57%. Based on the cutoff value of 97%, 13 BstZI pulsotypes were determined within the MRSA ST398 isolates (Fig. 2).
[image:5.585.42.543.71.436.2]Using SacII, the AA-MRSA isolates clustered together with a similarity of 76%, whereas the two reference strains were an outgroup, with a similarity of 61% with the MRSA ST398 isolates (Fig. 3). Thirteen SacII pulsotypes were distinguished among the AA-MRSA isolates, with a cutoff value of 97%. The FIG. 2. Dendrogram and fingerprints for PFGE using restriction enzyme BstZI with 32 MRSA ST398 isolates and two reference MRSA strains. The similarities among the fingerprints were calculated using the Dice coefficient (optimization, 1%; position tolerance, 0.5% to 1%), and the fingerprints were grouped according to their similarities by use of the UPGMA algorithm. The vertical gray line shows the delineation level of 97%.
on May 16, 2020 by guest
http://jcm.asm.org/
ST398 cluster could be further subdivided into two major groups (cutoff value of 80%), one group of isolates with SCCmectype V, and another group of isolates with SCCmec
type IVa. The same isolates grouped together with the restric-tion enzyme SacII as those that grouped with restricrestric-tion en-zyme BstZI, with the exception of three pulsotypes. Pulsotype Sac-9 contained pulsotypes BstZI-8 and BstZI-11; thus, in this case, BstZI was more discriminating. In contrast, restriction enzyme SacII was more discriminating for pulsotypes Sac-6 and Sac-7, which shared the same fingerprints when using restriction enzyme BstZI.
The AA-MRSA isolates clustered together with a similarity of 71% when using restriction enzyme ApaI (Fig. 4). Again, the two reference strains had a similarity of 62% or less with the MRSA ST398 cluster. As with restriction enzyme SacII, the ST398 cluster could be further subdivided into two groups (cutoff value of 80%), separating isolates carrying SCCmec
type IVa and those carrying SCCmectype V. With the cutoff
value of 97%, 13 different Apa pulsotypes were distinguished. However, there was only little correlation between the pulso-types obtained by BstZI or SacII and the pulsopulso-types obtained by ApaI. For example, pulsotype Apa-2 contained four differ-ent BstZI/SacII pulsotypes. On the other hand, isolates belong-ing to pulsotype BstZI-8 or Sac-9 were further divided into three pulsotypes using ApaI.
[image:6.585.43.531.70.472.2]Discriminatory power and concordance.The discriminatory indices (DI) of the methods, and those of the combination of the methods, are shown in Table 3. The discriminatory power of SCCmectyping,spatyping, and the MLVA for ST398 were quite low, as follows: 0.51, 0.74, and 0.81, respectively. The combination ofspa typing and the MLVA resulted in more discriminatory power (D⫽0.87) than that of the MLVA used alone but was still lower than that of PFGE when using a single restriction enzyme (D⫽0.88 to 0.92). For this data set, the DI of PFGE with the restriction enzymes BstZI and ApaI was as high as the DI of PFGE using all three restriction enzymes FIG. 3. Dendrogram and fingerprints for PFGE using restriction enzyme SacII with 32 MRSA ST398 isolates and two reference MRSA strains. The similarities among the fingerprints were calculated using the Dice coefficient (optimization, 1%; position tolerance, 0.5% to 1%), and the fingerprints were grouped according to their similarities by use of the UPGMA algorithm. The vertical gray line shows the delineation level of 97%.
on May 16, 2020 by guest
http://jcm.asm.org/
together or using those enzymes in combination with the MLVA (D⫽0.97). The combination of PFGE (when using all restriction enzymes or BstZI and ApaI) andspatyping had the most discriminatory power (D⫽0.98).
The following isolates were indistinguishable by all means applied: MB4392 and RIVM38; MB4362 and MB4366; RIVM29 and RIVM30; and RIVM21, RIVM33, and RIVM34. The concordance among different typing methods is shown in Tables 4 and 5. For the MLVA, the highest concordance was with PFGE using ApaI and withspatyping. As further expressed by the Wallace coefficient, the MLVA type was completely pre-dicted by PFGE using a combination of all the restriction en-zymes (W⫽1). The MLVA type was poorly predicted only byspa
typing (W⫽0.48), whereas thespatype was well predicted by MLVA (W⫽0.68). As already indicated above, the concordance between PFGE using restriction enzyme BstZI and PFGE using SacII was very high (Rand’s index⫽0.98; R⫽0.89). PFGE, using
a combination of all three restriction enzymes, was able to predict theSCCmec type and MLVA type completely (W⫽1.00) and had a high probability of predicting thespa type (W ⫽ 0.64). SCCmectype was a poor predictor for all typing methods (W⫽ 0.06 to 0.38), although it was well predicted by MLVA (W⫽0.98) and completely predicted using all PFGE methods (W⫽1.00).
DISCUSSION
[image:7.585.43.540.74.478.2]AA-MRSA strains of CC398 are not typeable using PFGE with SmaI digestion, the gold standard for MRSA typing. For this reason, until now, sequence-based typing methods, such as MLST andspatyping, have been used for typing AA-MRSA in epidemiological studies. Although these methods have the ad-vantage of reliability and excellent interlaboratory reproduc-ibility, their discriminatory power is weak and, consequently, less useful for short-term epidemiological studies such as an FIG. 4. Dendrogram and fingerprints for PFGE using restriction enzyme ApaI with 32 MRSA ST398 isolates and two reference MRSA strains. The similarities among the fingerprints were calculated using the Dice coefficient (optimization, 1%; position tolerance, 0.5% to 1%), and the fingerprints were grouped according to their similarities by use of the UPGMA algorithm. The vertical gray line shows the delineation level of 97%.
on May 16, 2020 by guest
http://jcm.asm.org/
examination of contamination routes on farms. To our knowl-edge, this is the first study exploring alternative fingerprinting techniques for subtyping MRSA ST398.
In this study, PFGE using the restriction enzymes XmaI (an isoschizomer of SmaI), BstZI, SacII, and ApaI was tested to obtain the enzymes’ discriminatory power with a set of 32 MRSA ST398 isolates. In addition, three recently developed MLVA techniques, successfully applied toStaphylococcus au-reusand more specifically to HA-MRSA and CA-MRSA, were evaluated and optimized for use with ST398 strains. Both the initial Polish system (15, 16, 17, 23, 24) and the Swiss system (10) are based on tandem repeats in genes, whereas the British system is based on staphylococcal interspersed repeat units, called SIRUs (11, 12). In this study, only primers with discrim-inatory power were included in this optimized MLVA specific for AA-MRSA.
All isolates were typeable using both the MLVA and PFGE. However, fingerprints obtained by PFGE using XmaI had weak patterns and were thus excluded from the study. The discriminatory power of PFGE, when using any of the remain-ing three restriction enzymes, was higher than that of SCCmec
typing,spatyping, the MLVA, or spatyping in combination with the MLVA. Within PFGE, the combination of all three
restriction enzymes had higher discrimination than the use of only one restriction enzyme. The discriminatory power of PFGE using only BstZI and ApaI was equally as good as using all three restriction enzymes. The discriminatory power of the MLVA, the second best typing method, was only a bit higher than that ofspatyping, althoughspatyping was also able to discriminate among identical MLVA types in a few cases and within a pulsotype on one occasion (using all three restriction enzymes or BstZI and ApaI). In this set of AA-MRSA isolates, the MLVA was not able to distinguish among isolates with the same PFGE profile. In agreement with our results, previously published reports of MLVA typing of clinical MRSA isolates had more discriminatory power than those ofspatyping and MLST (12, 15, 16, 17). In contrast, most studies comparing the MLVA with PFGE have shown that the discriminatory power of PFGE was comparable to that of the MLVA or only slightly better. Luczak-Kadlubowska et al. (16) and Sabat et al. (23) demonstrated that their MLVA system had a comparable dis-criminatory power to that of PFGE. The same group had reported previously that PFGE was superior to the MLVA. These findings were based on the setup of the study and the selection of the MRSA isolates (17). The results of the MLVA based on staphylococcal interspersed repeat units (11) de-pended on the set of isolates: for some isolates, the MLVA was able to distinguish within the same PFGE profile, but for others, the MLVA was less discriminatory than PFGE.
[image:8.585.42.284.89.272.2]It should be kept in mind that in the publications mentioned above, the MLVA consisted of a different combination of primer sets than the MLVA described here. Furthermore, in those publications, PFGE was performed using only the re-striction enzyme SmaI. The MLVA and PFGE were also used on a wide range of MLST andspatypes belonging to HA- and CA-MRSA but not to MRSA ST398. Those studies aimed to investigate whether the faster MLVA technique was able to predict PFGE typing, MLST, orspatyping. In order to do so, relatively low similarity cutoff values between 70% and 80% were used for the MLVA and PFGE (17, 27). This contrasts with the present study, during which the MLVA and PFGE techniques were evaluated for their power to distinguish among MRSA ST398 isolates. To this end, isolates that dif-fered even slightly were considered to belong to different types. For the MLVA, some differences were small. It is thus advis-able to use capillary electrophoresis for better resolution be-tween bands.
TABLE 3. Discriminatory power of the methods, apart and in combination, evaluated with 27 MRSA ST398 strains
Methoda No. of
types No. of unique isolates No. of clustered isolates No. of clusters Cluster size DI
SCCmectyping 2 0 27 2 12–15 0.51
spatyping 7 2 25 5 2–13 0.74
MLVA 9 5 22 4 2–10 0.81
spatyping⫹MLVA 11 7 20 4 2–8 0.87
PFGE (BstZI) 13 7 20 6 2–5 0.92
PFGE (SacII) 13 8 19 5 2–6 0.91
PFGE (ApaI) 13 9 18 4 2–8 0.88
PFGE
(BstZI⫹SacII)
14 9 18 5 2–5 0.92
PFGE
(BstZI⫹ApaI)
20 16 11 4 2–4 0.97
PFGE (RE) 20 16 11 4 2–4 0.97
PFGE
(RE)⫹MLVA
20 16 11 4 2–4 0.97
PFGE (RE)⫹spa
typing
21 16 11 5 2–3 0.98
a
RE, combination of all three restriction enzymes.
TABLE 4. Concordance among typing methods according to Rand’s and adjusted Rand’s indices
Typing method
Rand’s and adjusted Rand’s index values obtained using the indicated typing methods
spa SCCmec MLVA PFGE using BstZI PFGE using SacII PFGE using ApaI
Rand’s index Adjusted Rand’s index Rand’s index Adjusted Rand’s index Rand’s index Adjusted Rand’s index Rand’s index Adjusted Rand’s index Rand’s index Adjusted Rand’s index Rand’s index Adjusted Rand’s index
SCCmec 0.54 0.07
MLVA 0.80 0.44 0.70 0.38
PFGE using BstZI 0.73 0.11 0.59 0.17 0.82 0.26
PFGE using SacII 0.74 0.14 0.60 0.19 0.83 0.30 0.98 0.89
PFGE using ApaI 0.72 0.14 0.63 0.25 0.85 0.44 0.86 0.24 0.87 0.31
PFGE using REa 0.74 0.08 0.54 0.07 0.84 0.25 0.95 0.54 0.94 0.49 0.91 0.38
a
RE, combination of all three restriction enzymes.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:8.585.43.542.595.716.2]Even when combining all methods, some isolates from dif-ferent origins were indistinguishable from each other: pig iso-lates MB4392 and RIVM38 (one isolated in Belgium, the other in The Netherlands), MB4362 and MB4366 (the first isolated from a cow, the second from a pig), RIVM29 and RIVM30 (both isolated from Dutch pigs), and finally, RIVM21, RIVM33, and RIVM34 (also found in Dutch pig herds). It may be possible to distinguish between these isolates using other techniques. Another possibility, given the large number of an-imals transported within and between Belgium and The Neth-erlands, is that those isolates are the same strain circulating in different countries and in different animal species.
The concordance among the different methods was calcu-lated according to Rand’s and adjusted Rand’s indices and Wallace coefficients. As this is calculated for only a limited set of data, it should be interpreted carefully. According to the obtained results, thespatype was best predicted by MLVA, whereas the MLVA type was poorly predicted only by spa
typing. The MLVA type was completely predicted by PFGE using a combination of all restriction enzymes (W⫽1) but not vice versa. Tenover et al. (27) also concluded that the MLVA cannot be used to predict PFGE types.
In conclusion, both the MLVA and PFGE can be used to discriminate among isolates belonging to the newly emerging MRSA ST398 type. All isolates were typeable using both of the fingerprinting techniques and had better discriminatory power than the commonly used techniques. Although the MLVA is a faster technique, PFGE had more discriminatory power than the MLVA, especially when two restriction enzymes (i.e., BstZI and ApaI) were used. Both techniques can be used for epidemiological tracing of the MRSA ST398 isolates.
ACKNOWLEDGMENTS
We are grateful to M. Struelens from the Laboratoire de Re´fe´rence MRSA—Staphylocoques, Department of Microbiology, Universite´ Li-bre de Bruxelles, Hoˆpital Erasme, Brussels, Belgium, for providing strains MB4387, MB4388, and MB4389. We thank Miriam Levenson for critical reading of the manuscript.
REFERENCES
1.Barrett, F. F., R. F. McGehee, Jr., and M. Finland.1968. Methicillin-resis-tantStaphylococcus aureusat Boston City Hospital. Bacteriologic and
epi-demiologic observations. N. Engl. J. Med.279:441–448.
2.Bens, C. P. M., A. Voss, and C. H. W. Klaassen.2006. Presence of a novel
DNA methylation enzyme in methicillin-resistantStaphylococcus aureus
iso-lates associated with pig farming leads to uninterpretable results in standard
pulsed-field gel electrophoresis analysis. J. Clin. Microbiol.44:1875–1876.
3.Bikandi, J., R. S. Millan, A. Rementeria, and J. Garaizar.2004.In silico
analysis of complete bacterial genomes: PCR, AFLP-PCR and endonuclease
restriction. Bioinformatics20:798–799.
4.Carric¸o, J. A., C. Silva-Costa, J. Melo-Cristino, F. R. Pinto, H. de Lencastre, J. S. Almeida, and M. Ramirez.2006. Illustration of a common framework for relating multiple typing methods by application to macrolide-resistant
Streptococcus pyogenes. J. Clin. Microbiol.44:2524–2532.
5.de Neeling, A. J., M. J. M. van den Broek, E. C. Spalburg, M. G. van Santen-Verheuvel, W. D. C. Dam-Deisz, H. C. Boshuizen, A. W. van de Giessen, E. van Duijkeren, and X. W. Huijsdens.2007. High prevalence of
methicillin resistantStaphylococcus aureusin pigs. Vet. Microbiol.122:366–
372.
6.Denis, O., C. Suetens, M. Hallin, B. Catry, I. Ramboer, M. Dispas, G. Willems, B. Gordts, P. Butaye, and M. J. Struelens.2009. Methicillin-resis-tantStaphylococcus aureusST398 in swine farm personnel, Belgium. Emerg.
Infect. Dis.15:1098–1101.
7.Dewaele, I., I. De Man, A. Stael, P. Delputte, P. Butaye, G. Vlaemynck, L. Herman, M. Heyndrickx, and G. Rasschaert. 2008. Methicillin-resistant
Staphylococcus aureus(MRSA) on Belgian pig farms, abstr. S:8:5, p. 23. Abstr. ASM Conf. Antimicrob. Resist. Zoonotic Bacteria Foodborne Pathog.
8.Enright, M. C., N. P. J. Day, C. E. Davies, S. J. Peacock, and B. G. Spratt.
2000. Multilocus sequence typing for characterization of methicillin-resistant
and methicillin-susceptible clones ofStaphylococcus aureus. J. Clin.
Micro-biol.38:1008–1015.
9.Flamm, R. K., D. J. Hinrichs, and M. F. Thomashow.1984. Introduction of
pAM1 intoListeria monocytogenesby conjugation and homology between
nativeL. monocytogenesplasmids. Infect. Immun.44:157–161.
10.Francois, P., A. Huyghe, Y. Charbonnier, M. Bento, S. Herzig, I. Topopski, B. Fleury, D. Lew, P. Vaudaux, S. Harbarth, W. van Leeuwen, A. van Bel-kum, D. S. Blanc, D. Pittet, and J. Schrenzel.2005. Use of an automated multiple-locus, variable-number tandem repeat-based method for rapid and
high-throughput genotyping ofStaphylococcus aureusisolates. J. Clin.
Mi-crobiol.43:3346–3355.
11.Hardy, K. J., B. A. Oppenheim, S. Gossain, F. Gao, and P. Hawkey.2006. Use of variations in staphylococcal interspersed repeat units for molecular
typing of methicillin-resistantStaphylococcus aureusstrains. J. Clin.
Micro-biol.44:271–273.
12.Hardy, K. J., D. W. Ussery, B. A. Oppenheim, and P. M. Hawkey.2004. Distribution and characterization of staphylococcal interspersed repeat units
(SIRUs) and potential use for strain differentiation. Microbiology150:4045–
4052.
13.Hunter, P.1990. Reproducibility and indices of discriminatory power of
microbial typing methods. J. Clin. Microbiol.28:1903–1905.
14.Hunter, P. R., and M. A. Gaston.1988. Numerical index of the discrimina-tory ability of typing systems: an application of Simpson’s index of diversity.
J. Clin. Microbiol.26:2465–2466.
15.Karynski, M., A. J. Sabat, J. Empel, and W. Hryniewicz.2008. Molecular
surveillance of methicillin-resistantStaphylococcus aureusby multiple-locus
variable number tandem repeat fingerprinting (formerly multiple-locus
vari-able number tandem repeat analysis) andspa-typing in hierarchic approach.
Diagn. Microbiol. Infect. Dis.62:255–262.
16.Luczak-Kadlubowska, A., A. Sabat, A. Tambic-Andrasevic, M. Payerl-Pal, J. Krzyszton-Russjan, and W. Hryniewicz.2008. Usefulness of multiple-locus VNTR fingerprinting in detection of clonality of community- and
hospital-acquiredStaphylococcus aureusisolates. Antonie van Leeuwenhoek94:543–
553.
17.Malachowa, N., A. Sabat, M. Gniadkowski, J. Krzyszton-Russjan, J. Empel, J. Miedzobrodzki, K. Kosowska-Shick, P. C. Appelbaum, and W. Hryniewicz.
2005. Comparison of multiple-locus variable-number tandem-repeat analysis
with pulsed-field gel electrophoresis,spatyping, and multilocus sequence
typing for clonal characterization ofStaphylococcus aureusisolates. J. Clin.
Microbiol.43:3095–3100.
18.Milheiric¸o, C., D. C. Oliveira, and H. Lencastre. 2007. Multiplex PCR
[image:9.585.43.545.81.187.2]strategy for subtyping the staphylococcal cassette chromosomemectype IV
TABLE 5. Wallace’s coefficient values among typing methods
Typing method
Wallace’s coefficient value obtained using the indicated typing method
spa SCCmec MLVA PFGE using
BstZI PFGE using SacII PFGE using ApaI PFGE using REa
spa 0.56 0.48 0.14 0.17 0.2 0.08
SCCmec 0.30 0.38 0.16 0.19 0.25 0.06
MLVA 0.68 0.98 0.24 0.29 0.42 0.17
PFGE using BstZI 0.46 1.00 0.57 0.96 0.39 0.39
PFGE using SacII 0.50 1.00 0.59 0.84 0.44 0.34
PFGE using ApaI 0.45 1.00 0.67 0.26 0.33 0.26
PFGE using REa 0.64 1.00 1.00 1.00 1.00 1.00
aRE, combination of all three restriction enzymes.
on May 16, 2020 by guest
http://jcm.asm.org/
in methicillin-resistant Staphylococcus aureus: ‘SCCmecIV multiplex.’ J.
Antimicrob. Chemother.60:42–48.
19.Murchan, S., M. E. Kaufmann, A. Deplano, R. de Ryck, M. Struelens, C. E. Zinn, V. Fussing, S. Salmenlinna, J. Vuopio-Varkila, N. El Solh, C. Cuny, W. Witte, P. T. Tassios, N. Legakis, W. van Leeuwen, A. van Belkum, A. Vindel, I. Laconcha, J. Garaizar, S. Haeggman, B. Olsson-Liljequist, U. Ransjo, G. Coombes, and B. Cookson.2003. Harmonization of pulsed-field gel electro-phoresis protocols for epidemiological typing of strains of methicillin-resis-tantStaphylococcus aureus: a single approach developed by consensus in 10 European laboratories and its application for tracing the spread of related
strains. J. Clin. Microbiol.41:1574–1585.
20.Naimi, T. S., K. H. LeDell, K. Como-Sabetti, S. M. Borchardt, D. J. Boxrud, J. Etienne, S. K. Johnson, F. Vandenesch, S. Fridkin, C. O’Boyle, R. N. Danila, and R. Lynfield.2003. Comparison of community- and health
care-associated methicillin-resistantStaphylococcus aureusinfection. JAMA290:
2976–2984.
21.Oliveira, D. C., and H. Lencastre.2002. Multiplex PCR strategy for rapid
identification of structural types and variants of themecelement in
methi-cillin-resistantStaphylococcus aureus. Antimicrob. Agents Chemother.46:
2155–2161.
22.Persoons, D., S. Van Hoorebeke, K. Hermans, P. Butaye, A. de Kruif, F. Haesebrouck, and J. Dewulf.2009. Emerg. Infect. Dis.15:452–453. 23.Sabat, A., J. Krzyszton-Russjan, W. Strzalka, R. Filipek., K. Kosowska, W.
Hryniewicz, J. Travis, and J. Potempa.2003. New method for typing Staph-ylococcus aureusstrains: multiple-locus variable-number tandem repeat anal-ysis of polymorphism and genetic relationships of clinical isolates. J. Clin.
Microbiol.41:1801–1804.
24.Sabat, A., N. Malachowa, J. Miedzobrodzki, and W. Hryniewicz.2006.
Com-parison of PCR-based methods for typingStaphylococcus aureusisolates.
J. Clin. Microbiol.44:3804–3807.
25.Sabat, A., D. C. Melles, G. Martirosian, H. Grundmann, A. van Belkum, and W. Hryniewicz.2006. Distribution of the serine-aspartate repeat
protein-encodingsdrgenes among nasal carriage and invasiveStaphylococcus aureus
strains. J. Clin. Microbiol.44:1135–1138.
26.Struelens, M. J., A. Deplano, C. Godard, N. Maes, and E. Serruys.1992. Epidemiologic typing and delineation of genetic relatedness of
methicillin-resistant Staphylococcus aureus by macrorestriction analysis of genomic
DNA by using pulsed-field gel electrophoresis. J. Clin. Microbiol.30:2599–
2605.
27.Tenover, F. C., R. R. Vaughn, L. K. McDougal, G. E. Fosheim, and J. E. McGowan.2007. Multiple-locus variable-number tandem-repeat assay
anal-ysis of methicillin-resistantStaphylococcus aureusstrains. J. Clin. Microbiol.
45:2215–2219.
28.van Belkum, A., D. C. Melles, J. K. Peeters, W. B. van Leeuwen, E. van Duijkeren, X. W. Huijsdens, E. Spalburg, A. J. de Neeling, and H. A. Ver-brugh on behalf of the Dutch Working Party on Surveillance and Research of MRSA-SOM.2008. Methicillin-resistant and -susceptibleStaphylococcus aureussequence type 398 in pigs and humans. Emerg. Infect. Dis.14:479– 483.
29.van Loo, I., X. Huijsdens, E. Tiemersma, A. de Neeling, N. van de Sande-Bruinsma, D. Beaujean, A. Voss, and J. Kluytmans.2007. Emergence of
methicillin-resistant Staphylococcus aureus of animal origin in humans.
Emerg. Infect. Dis.13:1834–1839.
30.Voss, A., F. Loeffen, J. Bakker, C. Klaassen, and M. Wulf.2005.
Methicillin-resistantStaphylococcus aureusin pig farming. Emerg. Infect. Dis.11:1965–
1966.
31.Willems, G., M. Dispas, O. Denis, M. Hallin, C. Suetens, B. Gordts, M. Struelens, and P. Butaye.2007. Characterization of MRSA from pigs in Belgium, p. 18. Abstr. 2nd Symp. Antimicrob. Resist. Anim. Environ. (ARAE).
32.Wulf, M. W. H., and A. Voss.2008. MRSA in livestock animals—an epidemic
waiting to happen? Clin. Microbiol. Infect.14:519–521.
33.Zhang, K., J. A. McClure, S. Elsayed, T. Louie, and J. M. Conly.2005. Novel multiplex PCR assay for characterization and concomitant subtyping of
staphylococcal cassette chromosomemectypes I to V in methicillin-resistant
Staphylococcus aureus. J. Clin. Microbiol.43:5026–5033.