Cryptococcus
Isolates from Dogs and Cats in North America
Lisa M. Singer,aWieland Meyer,bCarolina Firacative,b,cGeorge R. Thompson III,dEileen Samitz,eJane E. Sykesf
Small Animal Clinical Sciences, Michigan State University College of Veterinary Medicine, East Lansing, Michigan, USAa; Molecular Mycology Research Laboratory, Center for Infectious Diseases and Microbiology, Sydney Medical School-Westmead Hospital, Marie Bashir Institute for Infectious Diseases and Biosecurity, The University of Sydney, Westmead Millennium Institute, Sydney, Australiab; Grupo de Microbiología, Instituto Nacional de Salud, Bogotá, Colombiac; Department of Medical Microbiology and Immunology, University of California Davis, Davis, California, USAd; Veterinary Medical Teaching Hospital, University of California Davis, Davis, California, USAe; Department of Medicine & Epidemiology, University of California Davis, Davis, California, USAf
Molecular types of theCryptococcus neoformans/Cryptococcus gattiispecies complex that infect dogs and cats differ regionally and with host species. Antifungal drug susceptibility can vary with molecular type, but the susceptibility ofCryptococcusisolates from dogs and cats is largely unknown.Cryptococcusisolates from 15 dogs and 27 cats were typed usingURA5restriction frag-ment length polymorphism analysis (RFLP), PCR fingerprinting, and multilocus sequence typing (MLST). Susceptibility was determined using a microdilution assay (Sensititre YeastOne; Trek Diagnostic Systems). MICs were compared among groups. The 42 isolates studied comprised molecular types VGI (7%), VGIIa (7%), VGIIb (5%), VGIIc (5%), VGIII (38%), VGIV (2%), VNI (33%), and VNII (2%), as determined byURA5RFLP. The VGIV isolate was more closely related to VGIII according to MLST. All VGIII isolates were from cats. All sequence types identified from veterinary isolates clustered with isolates from hu-mans. VGIII isolates showed considerable genetic diversity compared with otherCryptococcusmolecular types and could be di-vided into two major subgroups. Compared withC. neoformansMICs,C. gattiiMICs were lower for flucytosine, and VGIII MICs were lower for flucytosine and itraconazole. For all drugs except itraconazole,C. gattiiisolates exhibited a wider range of MICs thanC. neoformans. MICs varied withCryptococcusspecies and molecular type in dogs and cats, and MICs of VGIII iso-lates were most variable and may reflect phylogenetic diversity in this group. Because sequence types of dogs and cats reflect those infecting humans, these observations may also have implications for treatment of human cryptococcosis.
C
ryptococcosis, caused by the yeastsCryptococcus neoformansandCryptococcus gattii, is a worldwide invasive mycosis
caus-ing severe disease in humans and animals.Cryptococcusis the most
common systemic fungal pathogen of cats and can also cause
se-vere disseminated disease in dogs (1). In humans,C. neoformansis
opportunistic, usually infecting HIV/AIDS patients and other
im-munosuppressed individuals (2), whereasC. gattiiis more likely
to infect immunocompetent hosts (3). Cryptococcosis typically
occurs in cats with no obvious underlying immunodeficiency, and infections are not associated with concurrent retroviral infection
or other immunocompromised states (4,5). Among dogs,
pure-bred animals are most commonly affected, and an increased risk for breeds such as American cocker spaniels suggests an
underly-ing genetic immunodeficiency (6). In general, cryptococcosis in
humans is treated with a combination of amphotericin B and flu-cytosine (5FC), but high-dose (1,200 mg/day) fluconazole mono-therapy is often used in resource-poor health care settings, such as
among humans with HIV in sub-Saharan Africa (7). Similarly,
cats and dogs with cryptococcosis are often treated with flucona-zole monotherapy because of its relatively low cost, ease of admin-istration, and favorable pharmacokinetic properties. Response to treatment can be slow or inadequate, and reported rates of suc-cessful disease control or cure range from 7 to 100% in cats and
27% in dogs (8–11).
AmongC. gattiiandC. neoformansstrains, 8 molecular types
have been identified by PCR fingerprinting and amplified frag-ment length polymorphism analysis (AFLP): VNI, VNII, VNIII,
and VNIV forC. neoformansand VGI, VGII, VGIII, and VGIV for
C. gattii(12).C. gattiistrains that belong to molecular type VGII can be further categorized into a large number of subtypes. Of
those VGIIa, VGIIb, and VGIIc have been implicated in disease in immunocompetent humans, cats, dogs, and other animal species in British Columbia and the Pacific Northwest of the United States (3,13–15). An epidemiological study of dogs and cats with cryp-tococcosis in California showed that cats were most often infected withC. gattiimolecular type VGIII, whereas dogs were more likely
to be infected withC. neoformans, possibly reflecting host
differ-ences in susceptibility (6). Infections with strains of molecular
type VGIII in dogs have not yet been described, butC. gattii
mo-lecular type VGIII has emerged as a cause of disease in
immuno-compromised humans in southern California (16), and molecular
type VGIII was the predominantC. gattiimolecular type
identi-fied in humans from California in another recent report. Two VGIII subclusters have been identified in these individuals based on the results of multilocus sequence typing, VGIIIa and VGIIIb
(15,16). In eastern Australia, molecular type VGI predominates
amongC. gattiiisolates as a cause of disease in cats (17). VGIV is
Received3 December 2013 Returned for modification29 December 2013
Accepted26 March 2014
Published ahead of print2 April 2014
Editor:G. A. Land
Address correspondence to Jane Sykes, jesykes@ucdavis.edu.
Supplemental material for this article may be found athttp://dx.doi.org/10.1128 /JCM.03392-13.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JCM.03392-13
on May 16, 2020 by guest
http://jcm.asm.org/
considered to be a rare molecular type but has been detected in
humans and animals in sub-Saharan Africa (18).
The results of a number of studies have suggested that
differ-ences inin vitrodrug susceptibility are correlated with different
Cryptococcusspecies and molecular types (19–23). MICs of all azoles, particularly fluconazole, appear to be higher for molecular
type VGII isolates than for other C.gattiimolecular types,
primar-ily VGI, andC. neoformans(19, 21, 23). There is also growing
evidence that antifungal drug susceptibility may vary between
geographical locations for the same molecular type (19,21,24).
To date, no studies have compared antifungal drug suscepti-bility among molecular types isolated solely from dogs or cats in North America, with a focus on isolates from California. If infect-ing species or molecular type correlates with resistance to certain antifungal drugs, the initial drug of choice may differ regionally and differ between cats and dogs. Identification of drug
suscepti-bility patterns in isolates ofCryptococcusfrom dogs and cats may
also have relevance to the human population, because molecular
types ofCryptococcusthat infect animals can reflect those infecting
people that reside in the same geographic location (25). The
ob-jectives of this study were therefore to genetically characterize a
group ofCryptococcusisolates from dogs and cats from North
America using molecular methods and to determine the existence and strength of associations between antifungal drug MICs and
Cryptococcusspecies andCryptococcusmolecular types. Suscepti-bilities were determined using the Sensititre YeastOne (SYO) method (YO-9; Trek Diagnostic Systems, Inc., Cleveland, OH), because it is widely used for yeast susceptibility testing in
commer-cial diagnostic laboratories (26). We also sought to determine
whether fluconazole area-under-the-curve (AUC)/MIC ratios for the isolates in this study would be likely to achieve or exceed the
target ratio (ⱖ389.3) to prevent progressive fungal growth in the
central nervous system (CNS), which was previously determined
using a mouse meningoencephalitis model (7).
MATERIALS AND METHODS
Fungal isolation.Cryptococcal strains were isolated from swab speci-mens, biopsies, aspirates, or body fluids from dogs and cats with crypto-coccosis. Isolates were obtained from animals with cryptococcosis that were evaluated by veterinarians at the University of California, Davis, veterinary medical teaching hospital (VMTH) or from specimens submit-ted to the investigators’ laboratory by veterinarians or veterinary diagnos-tic laboratories between 2004 and 2011. Isolates not processed immedi-ately were retrieved from storage at⫺80°C and grown for a minimum of 3 days on inhibitory mold agar(Hardy Diagnostics, Santa Maria, CA) and potato flake agar at 30°C until individual colonies were isolated. Yeast colonies were subcultured to Sabouraud dextrose agar. Identification of
Cryptococcusspp. was based on morphology observed under light micros-copy after Gram staining and use of a commercial yeast identification kit according to the manufacturer’s instructions (API 20C AUX yeast identi-fication kit; bioMérieux, Durham, NC).C. neoformanswas differentiated fromC. gattiiwithL-canavanine– glycine– bromthymol blue agar (27).
Medical record review.Information obtained from medical records included each patient’s geographic location, species, breed, sex, age at time of diagnosis, history of antifungal drug therapy, site of specimen collection, method of specimen collection, and the date thatCryptococcus
spp. were isolated in culture. Geographic locations within California were divided into north coastal (cities west of Fairfield and north of Big Sur; climate zones 1, 2, 3, and 4), north central (cities east of Fairfield and north of Merced; climate zones 11 and 12), central (cities in the central valley that are south of Merced and north of Cahente; climate zone 13), and south coastal (cities west of Palm Springs and south of Big Sur; climate
zones 8 to 10). Sites of organ or tissue involvement were categorized as nasal (within the nasal cavity or a mass protruding from the nasal cavity), cutaneous (dermal nodules, including those originating on the bridge of the nose or eyelid; other superficial dermal masses; and draining wounds of the lips), CNS (central nervous system) (involving brain, spinal cord, or meninges), eyes (including conjunctiva), lungs (including pleura or pa-renchyma), lymph nodes, urinary tract (kidneys and urine), or other tissues, as described previously (6). Disease was described as local or dis-seminated (⬎1 organ system involved), although the possibility of dis-seminated disease could not be ruled out in animals with local lesions that did not subsequently have a full necropsy performed.
Molecular genotyping.High-molecular-weight genomic DNA was extracted and purified as previously described (28). Mating types were determined by PCR using the primers Mf␣U and Mf␣L (mating type␣) and MFa2U and MFa2L (mating typea), as previously reported (29). The major molecular types of theC. neoformans/C. gattiispecies complex were initially identified byURA5restriction fragment length polymorphism analysis (RFLP). TheURA5gene was first amplified via PCR with the primers URA5 (5=-ATGTCCTCCCAAGCCCTCGACTCCG-3=) and SJO1 (5=-TTAAGACCTCTGAACACCGTACTC-3=), as described previ-ously, followed by a double digestion with the restriction enzymes Sau96I and HhaI (28). The molecular subtypes VGIIa and VGIIb were identified via triple digestion of theURA5gene with the restriction enzymes HhaI, DdeI, and BsrGI, as described previously (30). Molecular subtypes VGIIa and VGIIb were also differentiated by means of PCR fingerprinting with the primer M13, as described previously (31). The patterns were assigned via visual comparison with patterns obtained for standard strains of the major molecular types of theC. neoformans/C. gattiispecies complex, including VNI (WM 148), VNII (WM626), VNIII (WM628), VNIV (WM629), VGI (WM179), VGII (WM178), VGIII (175), and VGIV (WM779), or for representative strains of the VGIIa (CDCR265) and VGIIb (CDCR272) molecular subtypes. MLST was performed on isolates using the ISHAM (International Society of Human and Animal Mycol-ogy) MLST consensus scheme for theC. neoformans/C. gattiispecies com-plex (12). Dendrograms showing the genetic relationships amongC. gattii
isolates and amongC. neoformansisolates were constructed using a soft-ware package (MEGA 5.05; Center for Evolutionary Medicine and Infor-matics, Tempe, AZ) based on maximum likelihood analysis of the con-catenated seven ISHAM consensus MLST loci. AdditionalC. gattiistrains included in the MLST analysis are listed in Table S1 in the supplemental material (15,28,32–37).
Antifungal drug susceptibility testing.The susceptibilities of the iso-lates in this study were determined by a commercially available colorimet-ric microdilution susceptibility test(Sensititre YeastOne YO-9; Trek Di-agnostic Systems, Inc., Cleveland, OH) performed according to the manufacturer’s instructions. Briefly, yeasts were subcultured on potato flake agar and incubated at 30°C for 24 h. Fungal colonies larger than 1 mm in diameter were collected using a sterile loop and added to 5 ml of sterile demineralized water and adjusted to a cell density of 0.5 McFarland standard (1⫻106to 5⫻106cells/ml). A total of 20l of yeast suspension was transferred into 11 ml of inoculum broth for a 1.5⫻103to 8⫻103 viable CFU/ml. The purity of the diluted cell suspension was determined by plating the McFarland standard and the inoculum broth on defi-brinated sheep blood agar. Colony counts were performed after 48 to 72 h of incubation to confirm purity and inoculum strength. The ranges of drug concentrations tested in 2-fold dilutions were as follows: amphoter-icin B (AMB), 0.008 to 16g/ml; fluconazole (FLC), 0.125 to 256g/ml; itraconazole (ITC), 0.008 to 16g/ml; flucytosine (5FC), 0.03 to 64g/ ml; voriconazole (VRC), 0.008 to 16g/ml; posaconazole (POS), 0.008 to 8g/ml; and caspofungin, anidulafungin, and micafungin, 0.008 to 16
g/ml.
A total of 100l of inoculum broth was placed in each well of the manufacturer’s plate using an autoinoculator (Trek/Sensititre autoinocu-lator with nephelometer; Trek Diagnostic Systems, Inc., Cleveland, OH). Reference strains (Candida parapsilosisATCC 22019 andCandida krusei
on May 16, 2020 by guest
http://jcm.asm.org/
ATCC 6258) were used for quality control purposes. Plates were incu-bated at 35°C in ambient air. Controls were read at 24 h, andCryptococcus
species MIC endpoints were read after 72 h of incubation or until the colonies grew to the 1 mm in diameter required to interpret the assay (up to 108 h of incubation) using a software program (Sensititre Trek/SWIN system with Sensitouch; Trek Diagnostic Systems, Inc., Cleveland, OH). Duplicate susceptibility tests were performed for isolates that required incubation times that exceeded 72 h in order to confirm assay results. Fluconazole AUC/MIC ratios were calculated for each isolate using pre-viously reported pharmacokinetic data available for dogs and cats, which estimate a mean AUC of 375 mg/h/liter after an oral dose of 50 mg in cats (38) and 268 mg/h/liter after an oral dose of 10 mg/kg in dogs (39). The AUC/MIC ratios were compared with the target ratio to inhibit fungal growth that was identified in a mouse meningoencephalitis model (7).
Statistical methods.The differences in MICs for each antifungal drug tested between two groups were compared using a Mann-Whit-ney test, with a null hypothesis that there were no differences between the groups. Group comparisons for MIC data includedC. gattiiversus
C. neoformans, molecular type VGIII versus allC. gattiiisolates that did not belong to the molecular type VGIII, molecular type VGIII versus VGII isolates, molecular type VGIII versusC. neoformansisolates, and isolates from animals with a history of antifungal drug therapy versus those that had not yet been treated with antifungal drugs. The F test was used to compare the distribution of MICs between each pair of groups. A chi-square analysis was used to compare the geographic distribution of isolates from cats to that of isolates from dogs. All analyses were per-formed with a statistical software package (MEGA 5.05; Center for Evo-lutionary Medicine and Informatics, Tempe, AZ) (GraphPad Prism,
ver-TABLE 1Source and molecular types ofCryptococcusisolates in this study
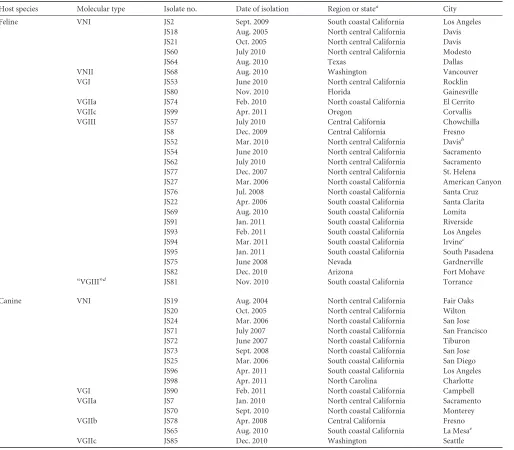
Host species Molecular type Isolate no. Date of isolation Region or statea City
Feline VNI JS2 Sept. 2009 South coastal California Los Angeles
JS18 Aug. 2005 North central California Davis
JS21 Oct. 2005 North central California Davis
JS60 July 2010 North central California Modesto
JS64 Aug. 2010 Texas Dallas
VNII JS68 Aug. 2010 Washington Vancouver
VGI JS53 June 2010 North central California Rocklin
JS80 Nov. 2010 Florida Gainesville
VGIIa JS74 Feb. 2010 North coastal California El Cerrito
VGIIc JS99 Apr. 2011 Oregon Corvallis
VGIII JS57 July 2010 Central California Chowchilla
JS8 Dec. 2009 Central California Fresno
JS52 Mar. 2010 North central California Davisb
JS54 June 2010 North central California Sacramento
JS62 July 2010 North central California Sacramento
JS77 Dec. 2007 North central California St. Helena
JS27 Mar. 2006 North coastal California American Canyon
JS76 Jul. 2008 North coastal California Santa Cruz
JS22 Apr. 2006 South coastal California Santa Clarita
JS69 Aug. 2010 South coastal California Lomita
JS91 Jan. 2011 South coastal California Riverside
JS93 Feb. 2011 South coastal California Los Angeles
JS94 Mar. 2011 South coastal California Irvinec
JS95 Jan. 2011 South coastal California South Pasadena
JS75 June 2008 Nevada Gardnerville
JS82 Dec. 2010 Arizona Fort Mohave
“VGIII”d JS81 Nov. 2010 South coastal California Torrance
Canine VNI JS19 Aug. 2004 North central California Fair Oaks
JS20 Oct. 2005 North central California Wilton
JS24 Mar. 2006 North coastal California San Jose
JS71 July 2007 North coastal California San Francisco
JS72 June 2007 North coastal California Tiburon
JS73 Sept. 2008 North coastal California San Jose
JS25 Mar. 2006 South coastal California San Diego
JS96 Apr. 2011 South coastal California Los Angeles
JS98 Apr. 2011 North Carolina Charlotte
VGI JS90 Feb. 2011 North coastal California Campbell
VGIIa JS7 Jan. 2010 North central California Sacramento
JS70 Sept. 2010 North coastal California Monterey
VGIIb JS78 Apr. 2008 Central California Fresno
JS65 Aug. 2010 South coastal California La Mesae
VGIIc JS85 Dec. 2010 Washington Seattle
a
Location of the animal’s residence at the timeCryptococcuswas isolated. bResided in southern California 3 years previously.
c
Resided in Arizona 7 months previously. dVGIV byURA5RFLP but VGIII by MLST analysis.
e
Recently moved from Vancouver, Canada.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:3.585.42.556.76.526.2]sion 4.00; GraphPad, San Diego, CA).Pvalues of⬍0.05 were considered significant.
Nucleotide sequence accession numbers.The GenBank accession numbers for the alleles in this study areKF667177toKF667240and KJ476370toKJ476394(see Tables S4 and S5 in the supplemental mate-rial).
RESULTS
Animals.A total of 42Cryptococcusisolates were obtained from 27 cats and 15 dogs. Twenty-two isolates were obtained from animals evaluated at the VMTH, and 20 isolates were obtained from dogs or cats evaluated by veterinarians outside the VMTH. Whether antifungal drug therapy had been administered before the isolate was obtained was known for 30 animals. Nine of the 30 animals had received antifungal drug therapy, and all had been treated for a week or longer without apparent response to treatment. Five animals had been treated with fluconazole, and 5 had been treated with itraconazole. Fourteen (52%) of the 27 cats were male, 12 (44%) were female, and all were neutered. Sex, age, and breed information was not available for one cat. The cats ranged in age from 1 to 12 years (median, 6 years). Twenty-two were of mixed breed, two were Siamese, one was a Maine coon, and one was a Persian. Disease sites identified included the nasal cavity, skin, CNS, lungs, myocardium, and mediastinum. The most common site of infection was the nasal cavity, in 11/27 (40%) of the cats. At least 8 of the 27 cats had disseminated disease.
Of the 15 dogs, 7 (47%) were male and 8 (53%) were female. One male and one female dog were intact, and the remaining dogs were neutered. The dogs ranged in age from 1.5 to 8 years (me-dian, 3 years). Breeds represented were recorded for all but one
dog, and included Labrador retrievers or their crosses (6), and one
each of Shar-Pei, Shetland sheepdog, Rhodesian ridgeback, Do-berman, basset hound, vizsla, German shepherd, and Yorkshire terrier. Sites of infection in the dogs included the nasal cavity, eye, CNS, kidneys, liver, lungs, gastrointestinal tract, and thyroid gland. The most common site of infection in dogs was the CNS, in 6/15 (40%) cases. At least 9 of the 15 dogs had disseminated dis-ease.
Isolate epidemiology.Twenty-seven isolates were identified as
C. gattii, and 15 wereC. neoformans. The molecular types isolated
and their geographic origins are shown inTable 1, and the MLST
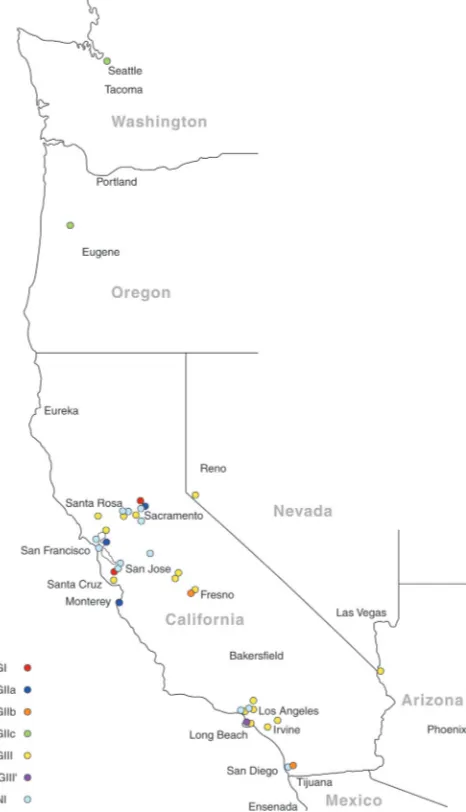
results are shown in Tables S2 and S3 in the supplemental mate-rial. The most prevalent molecular types were VGIII (16/42 [38%]) and VNI (14/42 [33%]). All of the VGIII isolates origi-nated from cats and were distributed throughout northern,
cen-tral, and southern California (Fig. 1). The most common
molec-ular type identified in dogs was VNI. There was no significant difference in the geographic location (by climate zone) for isolates obtained from dogs and those obtained from cats. All of the mo-lecular types identified were found in California except VGIIc and VNII. With the exception of a VGII isolate from a dog that had recently traveled to southern California from Vancouver in British Columbia, Canada, VGII isolates were not identified south of
Fresno (Fig. 1). Analysis of MLST results revealed that theC. gattii
andC. neoformansisolates from dogs and cats clustered with
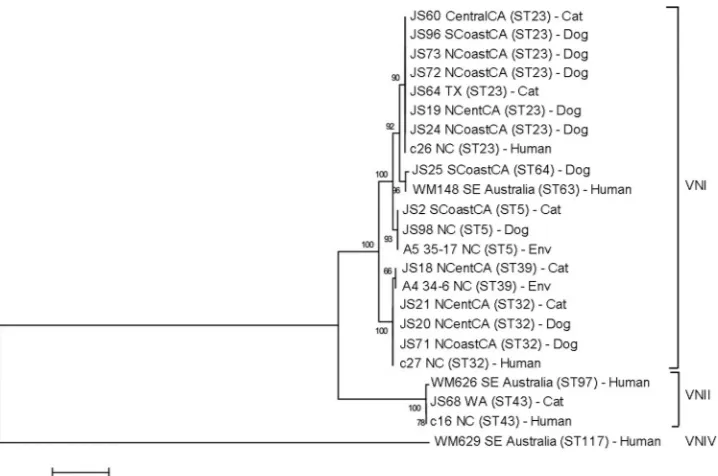
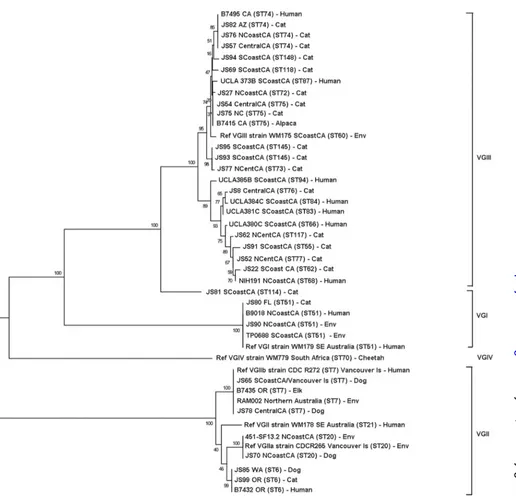
iso-lates from human patients (Fig. 2and3). The most prevalentC.
neoformanssequence type was ST23. VGIII isolates were
geneti-cally heterogeneous (Fig. 3; also, see Table S3 in the supplemental
material), and both mating types␣andawere identified. One of
the canine isolates identified as VGIV byURA5RFLP, strain JS81,
was more closely related to VGIII according to MLST.
Compari-son of the URA5gene sequence of this strain with that of the
reference VGIII and VGIV strains revealed 3 SNPs (single-nucle-otide polymorphisms) between strain JS81 and the reference VGIII strain and 30 SNPs between strain JS81 and the reference VGIV strain. The misclassification of this strain as a VGIV strain resulted from the existence of a SNP at the location of the Sau96I restriction site, generating an additional fragment, which made it identical to the VGIV restriction pattern.
Antifungal susceptibility testing.For 37 isolates, MICs could be determined at 72 h. Susceptibilities for four isolates (two VGIIa, one VGIIb, and one VGIII isolate) could not be clearly determined until 84 h of incubation, and susceptibilities for one VGIII isolate could not be clearly determined until 108 h of incu-bation.
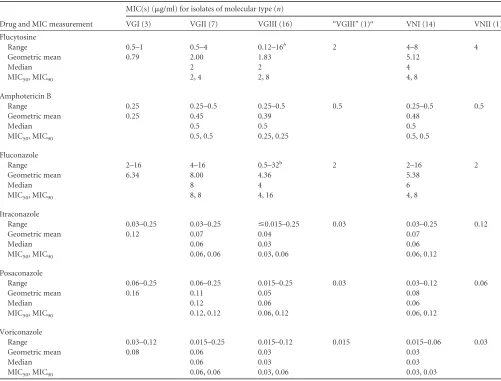
The MIC ranges, MIC50, MIC90, and geometric mean and
me-dian MICs forC. gattiiandC. neoformansof AMB, FLC, ITC, 5FC,
FIG 1Map showing the distribution ofCryptococcusmolecular types identi-fied in the current study from the western United States (n⫽38). The 4 isolates from other states are not shown. ‘VGIII’ indicates that the isolate was VGIV by
URA5RFLP but VGIII by MLST analysis.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:4.585.307.540.71.477.2]VRC, and POS are shown inTable 2. Six isolates had FLC MICs of
ⱖ16g/ml; three of these were VGIII isolates, and there was one
isolate each of VGI, VGIIc, and VNI. Only two isolates had FLC
MICs of 32 g/ml, and both were VGIII isolates. Flucytosine
MICs were lower amongC. gattiithanC. neoformansisolates (P⬍
0.001). The MICs of 5FC and ITC were lower among VGIII
iso-lates than they were amongC. neoformansisolates (P⫽0.02 for
both comparisons). Itraconazole MICs for molecular type VGIII
isolates were lower than those for isolates of all otherC. gattii
molecular types (P⫽0.01). Posaconazole MICs for molecular
type VGIII isolates appeared to be lower than those for isolates of
all otherC. gattiimolecular types and lower for VGIII isolates than
for VGII isolates, butPvalues only approached the cutoff for
significance for these observations (P⫽0.052 and 0.056,
respec-tively). There were no difference in MICs among isolates from dogs and cats and no difference in MICs of FLC or ITC between isolates from animals that had been treated with FLC or ITC and those that had not.
For all drugs except ITC,C. gattiiexhibited a wider variation in
MICs thanC. neoformansisolates (P⫽0.01, 0.04, 0.003, and 0.001
for AMB, 5FC, FLC, and POS, respectively, andP⬍0.001 for
VRC), which was also true forC. gattiimolecular type VGIII
com-pared withC. neoformansisolates (P⫽0.02, 0.007, and 0.004 for
AMB, 5FC, and POS, respectively, andP⬍0.001 for both FLC and
VRC). VGIII isolates exhibited a wider variation in MICs for FLC
and 5FC than isolates of all otherC. gattiimolecular types (P⬍
0.001 andP⫽0.01, respectively) and a wider variation in MICs for
FLC, 5FC, and VRC than VGII isolates (P⫽0.02, 0.02, and 0.01,
respectively) (Table 2).
For cats, the median FLC AUC/MIC ratio for allC. neoformans
isolates in this study was 70.3 (range, 23.4 to 187.5), and for dogs, it was 50.3 (range, 16.8 to 134). For cats, the median FLC AUC/
MIC ratio for allC. gattiiisolates in this study was 93.8 (range, 11.7
to 750), and for dogs, it was 50.3 (range, 8.4 to 536). A ratio that exceeded 389.3 was identified for only one isolate, which was from a cat.
DISCUSSION
In this study, we showed clearly that in California, the spectrum of
major molecular types of theCryptococcus neoformans/C. gattii
species complex infecting cats differs from that infecting dogs. The
most common molecular type to infect cats wasC. gattiimolecular
type VGIII, whereas dogs were more commonly infected by
mo-lecular type VNI (C. neoformansvar.grubii); no dogs were infected
by molecular type VGIII. This is in sharp contrast with the Pacific Northwest region of the United States and British Columbia,
where infection byC. gattiimolecular type VGII has
predomi-nated regardless of host species (14,34). Our study also confirmed
the widespread distribution ofC. gattiimolecular type VGIII in
cats from California. The reason for the predilection of this mo-lecular type for cats and not dogs is unclear, but it may relate to differences in host immunity, virulence properties of this organ-ism, and/or factors that put cats at increased risk of exposure to VGIII strains. Regional differences did not seem to be an explana-tion given that there was no difference in the geographic distribu-tion of isolates from dogs and cats, although analysis of a larger number of isolates would be required to confirm this observation.
Based on our phylogenetic analysis, bothC. neoformansandC.
gattiistrains isolated from dogs and cats in this study were closely
related to those isolated from humans. The most prevalentC.
neoformanssequence type, ST23, is a prevalent type found previ-ously in the United States, Asia, Africa, and Europe in both
envi-ronmental and clinical specimens (40–43). Other sequence types
identified in our study have been also reported before from Asia (ST5 and ST32), Africa (ST5, ST23, ST39, and ST43), and the
United States (ST32) (40–45). Molecular type VNI, VNII, VGI,
FIG 2Dendrogram showing phylogenetic relationships betweenC. neoformansisolates from the current study and 8 reference strains. Bootstrap values (%) are shown at each branch point. A total of 4,004 bp were aligned. NCoastCA, north coastal California; NCentCA, north central California; SCoastCA, south coastal California; CentralCA, central California; Env, environmental; SE, southeastern.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:5.585.114.473.65.303.2]and VGII isolates from dogs and cats in this study had sequence types that were identical or closely related to those of the reference VGI and VGII strains, respectively, supporting the more clonal
relationship of these populations as described previously (15,16).
Historically, VGIII has been one of the least prevalent molec-ular types found in humans and animals. In southeastern Austra-lia,C. gattiiisolates from dogs and cats are predominantly
molec-ular type VGI, whereas molecmolec-ular type VGII has been identified in
dogs and cats from Western Australia (4,10). The most prevalent
molecular type found in human patients with HIV/AIDS isC.
neoformansmolecular type VNI (2). However, infections withC. gattiimolecular type VGIII were recently identified in human
pa-tients with HIV/AIDS from southern California (16). A lower
number of molecular type VGIII isolates have been isolated from
FIG 3Dendrogram showing phylogenetic relationships betweenC. gattiiisolates from the current study and 20 reference strains. Molecular type VGIII isolates are clustered at the top of the dendrogram and can be divided into two major subgroups. Bootstrap values (%) are shown at each branch point. A total of 4,196 bp were aligned. NCoastCA, north coastal California; NCentCA, north central California; SCoastCA, south coastal California; CentralCA, central California; Env, environmental; SE, southeastern; Ref, reference.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:6.585.48.564.59.560.2]humans in other states, including those in the Pacific Northwest,
the south, Michigan, and Alaska (15), although the travel history
of many of the infected individuals in these states was unknown. Analysis of VGIII isolates infecting humans in the United States has shown a genetically diverse population of isolates that includes
both mating types␣andaand that can be separated into VGIIIa
and VGIIIb subclusters (15,16). The strains infecting cats in this
study also appeared to fall into two major subclusters, with both
mating types␣andabeing represented. A small number of feline
molecular type VGIII isolates in our study had matching sequence types (ST); these included 3 isolates from distant locations with ST74 (Arizona, coastal northern California, and central Califor-nia) and 2 isolates with ST145 from southern California. One isolate from southern California that was identified as VGIV by
URA5RFLP (JS81) appeared to be more closely related to VGIII.
We chose to use the colorimetric SYO method as opposed to standard broth microdilution susceptibility testing using methods defined by the Clinical Laboratories Standards Institute (CLSI), because the SYO method is widely used by clinical laboratories for
yeast susceptibility testing (26) and can be implemented in a
com-mercial veterinary diagnostic laboratory setting because it has a
convenient format and is affordable for pet owners. A comparison
of the SYO method to the CLSI method forCryptococcusfound
that essential agreements were 63.6% (FLC), 75% (ITC), 86.4%
(VRC), 77.3% (POS), 88.6% (5FC), and 86.4% (AMB) (46).
Given the relatively low number of isolates in this study, it was not possible to accurately determine epidemiological cutoff values
(ECVs), but 95% of VNI isolates in our study had MICs ofⱕ0.12
g/ml for POS and ITC,ⱕ0.06g/ml for VOR, andⱕ8g/ml for
FLC; this compared with statistical ECVs that included 95% of the
wild-type isolates of 0.25g/ml for POS, ITC, and VOR and 8
g/ml for FLC using broth microdilution (47). In our study, 95%
of VGIII isolates had MICs ofⱕ0.12g/ml for ITC and 32g/ml
for FLC; this compared with statistical ECVs of 0.5g/ml for ITC
and 8g/ml for FLC (47). The 1- to 2-fold-dilution discrepancies
in these numbers could reflect variations in methods used or true differences in susceptibility in the collection of isolates studied and/or reflect the relatively low numbers of isolates in our study. In the future, larger numbers of isolates should be used to
deter-mine ECVs using the SYO method, as has been done forCandida
(26).
[image:7.585.41.542.76.457.2]In general, for any of the tested antifungal drugs,C. gattiiVGIII
TABLE 2MICs for 27C. gattiiand 15C. neoformansisolates from dogs and cats
Drug and MIC measurement
MIC(s) (g/ml) for isolates of molecular type (n)
VGI (3) VGII (7) VGIII (16) “VGIII” (1)a VNI (14) VNII (1)
Flucytosine
Range 0.5–1 0.5–4 0.12–16b 2 4–8 4
Geometric mean 0.79 2.00 1.83 5.12
Median 2 2 4
MIC50, MIC90 2, 4 2, 8 4, 8
Amphotericin B
Range 0.25 0.25–0.5 0.25–0.5 0.5 0.25–0.5 0.5
Geometric mean 0.25 0.45 0.39 0.48
Median 0.5 0.5 0.5
MIC50, MIC90 0.5, 0.5 0.25, 0.25 0.5, 0.5
Fluconazole
Range 2–16 4–16 0.5–32b 2 2–16 2
Geometric mean 6.34 8.00 4.36 5.38
Median 8 4 6
MIC50, MIC90 8, 8 4, 16 4, 8
Itraconazole
Range 0.03–0.25 0.03–0.25 ⱕ0.015–0.25 0.03 0.03–0.25 0.12
Geometric mean 0.12 0.07 0.04 0.07
Median 0.06 0.03 0.06
MIC50, MIC90 0.06, 0.06 0.03, 0.06 0.06, 0.12
Posaconazole
Range 0.06–0.25 0.06–0.25 0.015–0.25 0.03 0.03–0.12 0.06
Geometric mean 0.16 0.11 0.05 0.08
Median 0.12 0.06 0.06
MIC50, MIC90 0.12, 0.12 0.06, 0.12 0.06, 0.12
Voriconazole
Range 0.03–0.12 0.015–0.25 0.015–0.12 0.015 0.015–0.06 0.03
Geometric mean 0.08 0.06 0.03 0.03
Median 0.06 0.03 0.03
MIC50, MIC90 0.06, 0.06 0.03, 0.06 0.03, 0.03
a
VGIV byURA5RFLP but VGIII by MLST analysis.
bVariance significantly wider than for molecular types other than VGIII.
on May 16, 2020 by guest
http://jcm.asm.org/
isolates had lower MICs thanC. neoformansisolates or all otherC. gattiiisolates combined. Of perhaps greater significance,C. gattii
isolates exhibited a significantly wider range of MICs thanC.
neo-formansisolates. The same was true for the VGIII isolates
com-pared with the isolates of all otherC. gattiimolecular types. Of the
6 isolates with FLC MICs ofⱖ16g/ml, 3 were VGIII isolates, and
FLC MICs of 32g/ml were recorded only among VGIII isolates.
In other studies, clinical and environmental isolates ofC. gattii
had higher MICs thanC. neoformansisolates (48), and VGII
iso-lates had higher MICs than the isoiso-lates of all otherC. gattii
mo-lecular types (19). VGII isolates in this study appeared to have
higher MICs for most antifungal drugs than the isolates of all other
C. gattiimolecular types, although the relatively low number of VGII isolates in this study limited our ability to document signif-icance and to compare our results with those of other investiga-tors.
Although combination therapy with AMB and FLC or 5FC has
been recommended for animals with disseminated disease (1,8),
the use of AMB and 5FC has been limited by expense, and there is a high incidence of cutaneous adverse effects in dogs treated with
5FC (49). High FLC MICs identified in some strains ofC. gattii
might not correlate with treatment failurein vivo, and clinical
breakpoints that define susceptibility versus resistance have not
been clearly defined forCryptococcusspp. in humans or animals. A
history of FLC or ITC therapy (without clinical response) was not associated with higher MICs of these drugs in this study. Extrap-olation of data from a mouse model of cryptococcal meningoen-cephalitis to HIV-infected humans suggested that high-dose FLC monotherapy might achieve or exceed the target AUC/MIC ratio
(ⱖ389.3) required to inhibit fungal growth across the expected
distribution of MICs for C. neoformans in only two-thirds of
treated patients (37). With the assumption that results obtained
with the SYO method correlate with those using CLSI methods, we estimated that the FLC AUC/MIC ratio for cats and dogs with
meningoencephalitis caused by theC. neoformansisolates in this
study would also not be predicted to approach this target. If the
same target is applied forC. gattiimeningoencephalitis,
progres-sive cryptococcal growth would be expected in all of the dogs and
⬎90% of the cats in this study had they been treated with 10 mg/kg
FLC daily. This target would also not have been attainable for 12C.
gattiiisolates from dogs and cats for which susceptibilities were
reported using CLSI methods (20). Thus, as suggested for
hu-mans, FLC monotherapy may not be an appropriate treatment, at least for animals with meningoencephalitis, and this might ex-plain the poor responses to treatment in some patients in the face of relatively low MICs.
In summary, this study shows that (i) the molecular types that infect dogs and cats in California differ, but isolates from both dogs and cats genetically resemble those infecting humans; (ii) VGIII isolates that infect cats from California show a high genetic diversity and can be separated into two major subclusters (as doc-umented in humans); (iii) significant differences in the distribu-tion of MICs exist among cryptococcal molecular types that infect dogs and cats, with molecular type VGIII isolates exhibiting a wider range of MICs than other molecular types; and (iv) the use of fluconazole at currently employed dosing regimens may not be
adequate to effectively inhibit growth ofCryptococcusin dogs and
cats with meningoencephalitis. The variability in antifungal drug susceptibility identified among VGIII isolates may relate to the high level of genetic diversity that existed in this group. This is the
first time that a possible correlation between genetic diversity and
variable antifungal drug susceptibility within aCryptococcus
mo-lecular type has been identified. The data reported in this study should also be useful for monitoring for the appearance of more resistant strains using the SYO method.
ACKNOWLEDGMENTS
We thank the following people for supplying strains listed in Table S1 in the supplemental material: Sharon C. A. Chen, June Kwon-Chung, Val-erie Davis, David Ellis, Murray Fyfe, Texter Howard, Thomas Mitchell, and Tania J. Pfeiffer. We also thank LeAnn Lindsay for her technical support.
This work was supported by an NH&MRC project grant APP1031943 to Wieland Meyer and a grant from the University of California, Davis Center for Companion Animal Health (2010-34-R). Carolina Firacative was supported by a Ph.D. scholarship “Becas Francisco José de Caldas” from COLCIENCIAS Colombia.
George R. Thompson has consulted for Astellas, the manufacturer of isavuconazole.
REFERENCES
1.Sykes JE, Malik R.2014. Cryptococcosis, p 599 – 612.InSykes JE (ed), Canine and feline infectious diseases. Elsevier, St. Louis, MO.
2.Fries BC, Cox GM.2011. Cryptococcosis in AIDS, p 515–525.InHeitman J, Kozel TR, Kwon-Chung KJ, Perfect J, Casadevall A (ed),Cryptococcus— from human pathogen to model yeast. ASM Press, Washington, DC. 3.Bartlett KH, Kidd SE, Kronstad JW.2008. The emergence of
Cryptococ-cus gattiiin British Columbia and the Pacific Northwest. Curr. Infect. Dis. Rep.10:58 – 65.http://dx.doi.org/10.1007/s11908-008-0011-1.
4.O’Brien CR, Krockenberger MB, Wigney DI, Martin P, Malik R.2004. Retrospective study of feline and canine cryptococcosis in Australia from 1981 to 2001: 195 cases. Med. Mycol.42:449 – 460.http://dx.doi.org/10 .1080/13693780310001624547.
5.Trivedi SR, Malik R, Meyer W, Sykes JE.2011. Feline cryptococcosis: impact of current research on clinical management. J. Feline Med. Surg.
13:163–172.http://dx.doi.org/10.1016/j.jfms.2011.01.009.
6.Trivedi SR, Sykes JE, Cannon MS, Wisner ER, Meyer W, Sturges BK, Dickinson PJ, Johnson LR.2011. Clinical features and epidemiology of cryptococcosis in cats and dogs in California: 93 cases (1988 –2010). J. Am. Vet. Med. Assoc. 239:357–369. http://dx.doi.org/10.2460/javma.239.3 .357.
7.Sudan A, Livermore J, Howard SJ, Al-Nakeeb Z, Sharp A, Goodwin J, Gregson L, Warn PA, Felton TW, Perfect JR, Harrison TS, Hope WW.
2013. Pharmacokinetics and pharmacodynamics of fluconazole for cryp-tococcal meningoencephalitis: implications for antifungal therapy and in vitro susceptibility breakpoints. Antimicrob. Agents Chemother.57:
2793–2800.http://dx.doi.org/10.1128/AAC.00216-13.
8.O’Brien CR, Krockenberger MB, Martin P, Wigney DI, Malik R.2006. Long-term outcome of therapy for 59 cats and 11 dogs with cryptococco-sis. Aust. Vet. J.84:384 –392.http://dx.doi.org/10.1111/j.1751-0813.2006 .00040.x.
9.Sykes JE, Sturges BK, Cannon MS, Gericota B, Higgins RJ, Trivedi SR, Dickinson PJ, Vernau KM, Meyer W, Wisner ER.2010. Clinical signs, imaging features, neuropathology, and outcome in cats and dogs with central nervous system cryptococcosis from California. J. Vet. Intern. Med.24:1427–1438.http://dx.doi.org/10.1111/j.1939-1676.2010.0633.x. 10. McGill S, Malik R, Saul N, Beetson S, Secombe C, Robertson I, Irwin P.
2009. Cryptococcosis in domestic animals in Western Australia: a retro-spective study from 1995–2006. Med. Mycol.47:625– 639.http://dx.doi .org/10.1080/13693780802512519.
11. Lester SJ, Kowalewich NJ, Bartlett KH, Krockenberger MB, Fairfax TM, Malik R.2004. Clinicopathologic features of an unusual outbreak of cryp-tococcosis in dogs, cats, ferrets, and a bird: 38 cases (January to July 2003). J. Am. Vet. Med. Assoc.225:1716 –1722.http://dx.doi.org/10.2460/javma .2004.225.1716.
12. Meyer W, Aanensen DM, Boekhout T, Cogliati M, Diaz MR, Esposto MC, Fisher M, Gilgado F, Hagen F, Kaocharoen S, Litvintseva AP, Mitchell TG, Simwami SP, Trilles L, Viviani MA, Kwon-Chung J.2009. Consensus multi-locus sequence typing scheme forCryptococcus
on May 16, 2020 by guest
http://jcm.asm.org/
mansandCryptococcus gattii. Med. Mycol.47:561–570.http://dx.doi.org /10.1080/13693780902953886.
13. Gillece JD, Schupp JM, Balajee SA, Harris J, Pearson T, Yan Y, Keim P, DeBess E, Marsden-Haug N, Wohrle R, Engelthaler DM, Lockhart SR.
2011. Whole genome sequence analysis ofCryptococcus gattiifrom the Pacific Northwest reveals unexpected diversity. PLoS One6:e28550.http: //dx.doi.org/10.1371/journal.pone.0028550.
14. Byrnes EJ, III, Li W, Lewit Y, Ma H, Voelz K, Ren P, Carter DA, Chaturvedi V, Bildfell RJ, May RC, Heitman J.2010. Emergence and pathogenicity of highly virulentCryptococcus gattiigenotypes in the north-west United States. PLoS Pathog.6:e1000850.http://dx.doi.org/10.1371 /journal.ppat.1000850.
15. Lockhart SR, Iqbal N, Harris JR, Grossman NT, DeBess E, Wohrle R, Marsden-Haug N, Vugia DJ. 2013.Cryptococcus gattiiin the United States: genotypic diversity of human and veterinary isolates. PLoS One
8:e74737.http://dx.doi.org/10.1371/journal.pone.0074737.
16. Byrnes EJ, III, Li W, Ren P, Lewit Y, Voelz K, Fraser JA, Dietrich FS, May RC, Chaturvedi S, Chaturvedi V, Heitman J. 2011. A diverse population ofCryptococcus gattiimolecular type VGIII in southern Cali-fornian HIV/AIDS patients. PLoS Pathog.7:e1002205.http://dx.doi.org /10.1371/journal.ppat.1002205.
17. Campbell LT, Currie BJ, Krockenberger M, Malik R, Meyer W, Heit-man J, Carter D.2005. Clonality and recombination in genetically differ-entiated subgroups ofCryptococcus gattii. Eukaryot. Cell4:1403–1409. http://dx.doi.org/10.1128/EC.4.8.1403-1409.2005.
18. Litvintseva AP, Thakur R, Reller LB, Mitchell TG.2005. Prevalence of clinical isolates ofCryptococcus gattiiserotype C among patients with AIDS in sub-Saharan Africa. J. Infect. Dis.192:888 – 892.http://dx.doi.org /10.1086/432486.
19. Lockhart SR, Iqbal N, Bolden CB, DeBess EE, Marsden-Haug N, Worhle R, Thakur R, Harris JR.2012. Epidemiologic cutoff values for triazole drugs inCryptococcus gattii: correlation of molecular type and in vitro susceptibility. Diagn. Microbiol. Infect. Dis.73:144 –148.http://dx .doi.org/10.1016/j.diagmicrobio.2012.02.018.
20. Iqbal N, DeBess EE, Wohrle R, Sun B, Nett RJ, Ahlquist AM, Chiller T, Lockhart SR.2010. Correlation of genotype and in vitro susceptibilities of
Cryptococcus gattiistrains from the Pacific Northwest of the United States. J. Clin. Microbiol.48:539 –544.http://dx.doi.org/10.1128/JCM.01505-09. 21. Chong HS, Dagg R, Malik R, Chen S, Carter D.2010. In vitro suscep-tibility of the yeast pathogen cryptococcus to fluconazole and other azoles varies with molecular genotype. J. Clin. Microbiol.48:4115– 4120.http: //dx.doi.org/10.1128/JCM.01271-10.
22. Trilles L, Meyer W, Wanke B, Guarro J, Lazéra M.2012. Correlation of antifungal susceptibility and molecular type within theCryptococcus neo-formans/C. gattiispecies complex. Med. Mycol.50:328 –332.http://dx.doi .org/10.3109/13693786.2011.602126.
23. Hagen F, Illnait-Zaragozi MT, Bartlett KH, Swinne D, Geertsen E, Klaassen CH, Boekhout T, Meis JF.2010. In vitro antifungal suscepti-bilities and amplified fragment length polymorphism genotyping of a worldwide collection of 350 clinical, veterinary, and environmental Cryp-tococcus gattiiisolates. Antimicrob. Agents Chemother.54:5139 –5145. http://dx.doi.org/10.1128/AAC.00746-10.
24. Chowdhary A, Randhawa HS, Sundar G, Kathuria S, Prakash A, Khan Z, Sun S, Xu J.2011. In vitro antifungal susceptibility profiles and geno-types of 308 clinical and environmental isolates ofCryptococcus neofor-mans var. grubiiandCryptococcus gattiiserotype B from north-western India. J. Med. Microbiol.60:961–967.http://dx.doi.org/10.1099/jmm.0 .029025-0.
25. Duncan C, Schwantje H, Stephen C, Campbell J, Bartlett K. 2006.
Cryptococcus gattiiin wildlife of Vancouver Island, British Columbia, Can-ada. J. Wildl. Dis.42:175–178.http://dx.doi.org/10.7589/0090-3558-42.1 .175.
26. Cantón E, Pemán J, Iñiguez C, Hervás D, Lopez-Hontangas JL, Pina-Vaz C, Camarena JJ, Campos-Herrero I, García-García I, García-Tapia AM, Guna R, Merino P, Pérez del Molino L, Rubio C, Suárez A, FUNGEMYCA Study Group.2013. Epidemiological cutoff values for fluconazole, itraconazole, posaconazole, and voriconazole for sixCandida
species as determined by the colorimetric Sensititre YeastOne method. J. Clin. Microbiol.51:2691–2695.http://dx.doi.org/10.1128/JCM.01230-13. 27. Kwon-Chung KJ, Polacheck I, Bennett JE.1982. Improved diagnostic medium for separation ofCryptococcus neoformans var. neoformans (sero-types A and D) andCryptococcus neoformansvar.gattii(serotypes B and C). J. Clin. Microbiol.15:535–537.
28. Meyer W, Castaneda A, Jackson S, Huynh M, Castañeda E. 2003. Molecular typing of IberoAmericanCryptococcus neoformansisolates. Emerg. Infect. Dis.9:189 –195.http://dx.doi.org/10.3201/eid0902.020246. 29. Halliday CL, Bui T, Krockenberger M, Malik R, Ellis DH, Carter DA.
1999. Presence of alpha and a mating types in environmental and clinical collections ofCryptococcus neoformans var. gattiistrains from Australia. J. Clin. Microbiol.37:2920 –2926.
30. MacDougall L, Kidd SE, Galanis E, Mak S, Leslie MJ, Cieslak PR, Kronstad JW, Morshed MG, Bartlett KH.2007. Spread ofCryptococcus gattiiin British Columbia, Canada, and detection in the Pacific Northwest, U. S. A. Emerg. Infect. Dis.13:42–50.http://dx.doi.org/10.3201/eid1301 .060827.
31. Meyer W, Mitchell TG.1995. Polymerase chain reaction fingerprinting in fungi using single primers specific to minisatellites and simple repetitive DNA sequences: strain variation inCryptococcus neoformans. Electropho-resis16:1648 –1656.http://dx.doi.org/10.1002/elps.11501601273. 32. Pfeiffer T, Ellis D.1991. Environmental isolation ofCryptococcus
neofor-mans gattiifrom California. J. Infect. Dis.163:929 –930.http://dx.doi.org /10.1093/infdis/163.4.929.
33. Carriconde F, Gilgado F, Arthur I, Ellis D, Malik R, van de Wiele N, Robert V, Currie BJ, Meyer W.2011. Clonality and alpha-a recombina-tion in the AustralianCryptococcus gattiiVGII population—an emerging outbreak in Australia. PLoS One 6:e16936. http://dx.doi.org/10.1371 /journal.pone.0016936.
34. Kidd SE, Hagen F, Tscharke RL, Huynh M, Bartlett KH, Fyfe M, Macdougall L, Boekhout T, Kwon-Chung KJ, Meyer W.2004. A rare genotype ofCryptococcus gattiicaused the cryptococcosis outbreak on Vancouver Island (British Columbia, Canada). Proc. Natl. Acad. Sci. U. S. A.101:17258 –17263.http://dx.doi.org/10.1073/pnas.0402981101. 35. Meyer W, Mitchell TG, Freedman EZ, Vilgalys R.1993. Hybridization
probes for conventional DNA fingerprinting used as single primers in the polymerase chain reaction to distinguish strains ofCryptococcus neofor-mans. J. Clin. Microbiol.31:2274 –2280.
36. Kwon-Chung KJ, Bennett JE, Theodore TS.1978.Cryptococcus bacillis-porussp. nov.: serotype B-C ofCryptococcus neoformans. Int. J. Syst. Bac-teriol.28:616 – 620.http://dx.doi.org/10.1099/00207713-28-4-616. 37. Litvintseva AP, Thakur R, Vilgalys R, Mitchell TG.2006. Multilocus
sequence typing reveals three genetic subpopulations ofCryptococcus neo-formans var. grubii(serotype A), including a unique population in Bo-tswana. Genetics172:2223–2238.http://dx.doi.org/10.1534/genetics.105 .046672.
38. Vaden SL, Heit MC, Hawkins EC, Manaugh C, Riviere JE. 1997. Fluconazole in cats: pharmacokinetics following intravenous and oral ad-ministration and penetration into cerebrospinal fluid, aqueous humour and pulmonary epithelial lining fluid. J. Vet. Pharmacol. Ther.20:181– 186.http://dx.doi.org/10.1111/j.1365-2885.1997.tb00093.x.
39. Humphrey MJ, Jevons S, Tarbit MH.1985. Pharmacokinetic evaluation of UK-49,858, a metabolically stable triazole antifungal drug, in animals and humans. Antimicrob. Agents Chemother.28:648 – 653.http://dx.doi .org/10.1128/AAC.28.5.648.
40. Cogliati M, Zamfirova RR, Tortorano AM, Viviani MA, Fimua Cryp-tococcosis Network.2013. Molecular epidemiology of Italian clinical
Cryptococcus neoformansvar.grubiiisolates. Med. Mycol.51:499 –506. http://dx.doi.org/10.3109/13693786.2012.751642.
41. Khayhan K, Hagen F, Pan W, Simwami S, Fisher MC, Wahyuningsih R, Chakrabarti A, Chowdhary A, Ikeda R, Taj-Aldeen SJ, Khan Z, Ip M, Imran D, Sjam R, Sriburee P, Liao W, Chaicumpar K, Vuddhakul V, Meyer W, Trilles L, van Iersel LJ, Meis JF, Klaassen CH, Boekhout T.
2013. Geographically structured populations ofCryptococcus neoformans
varietygrubiiin Asia correlate with HIV status and show a clonal popula-tion structure. PLoS One 8:e72222. http://dx.doi.org/10.1371/journal .pone.0072222.
42. Choi YH, Ngamskulrungroj P, Varma A, Sionov E, Hwang SM, Carri-conde F, Meyer W, Litvintseva AP, Lee WG, Shin JH, Kim EC, Lee KW, Choi TY, Lee YS, Kwon-Chung KJ.2010. Prevalence of the VNIc geno-type ofCryptococcus neoformansin non-HIV-associated cryptococcosis in the Republic of Korea. FEMS Yeast Res.10:769 –778.http://dx.doi.org/10 .1111/j.1567-1364.2010.00648.x.
43. Wiesner DL, Moskalenko O, Corcoran JM, McDonald T, Rolfes MA, Meya DB, Kajumbula H, Kambugu A, Bohjanen PR, Knight JF, Boul-ware DR, Nielsen K.2012. Cryptococcal genotype influences immuno-logic response and human clinical outcome after meningitis. mBio
3:e00196 –12.http://dx.doi.org/10.1128/mBio.00196-12.
on May 16, 2020 by guest
http://jcm.asm.org/
44. Mihara T, Izumikawa K, Kakeya H, Ngamskulrungroj P, Umeyama T, Takazono T, Tashiro M, Nakamura S, Imamura Y, Miyazaki T, Ohno H, Yamamoto Y, Yanagihara K, Miyzaki Y, Kohno S.2013. Multilocus sequence typing ofCryptococcus neoformansin non-HIV associated cryp-tococcosis in Nagasaki, Japan. Med. Mycol.51:252–260.http://dx.doi.org /10.3109/13693786.2012.708883.
45. Simwami SP, Khayhan K, Henk DA, Aanensen DM, Boekhout T, Hagen F, Brouwer AE, Harrison TS, Donnelly CA, Fisher MC.2011. Low diversityCryptococcus neoformansvarietygrubiimultilocus sequence types from Thailand are consistent with an ancestral African origin. PLoS Pathog.7:e1001343.http://dx.doi.org/10.1371/journal.ppat.1001343. 46. Bertout S, Dunyach C, Drakulovski P, Reynes J, Mallié M.2011. Comparison
of the Sensititre YeastOne dilution method with the Clinical and Laboratory Stan-dards Institute (CLSI) M27–A3 microbroth dilution reference method for deter-mining MIC of eight antifungal agents on 102 yeast strains. Pathol. Biol. (Paris)
59:48–51.http://dx.doi.org/10.1016/j.patbio.2010.07.020.
47. Espinel-Ingroff A, Aller AI, Canton E, Castañón-Olivares LR, Chowd-hary A, Cordoba S, Cuenca-Estrella M, Fothergill A, Fuller J, Govender N, Hagen F, Illnait-Zaragozi MT, Johnson E, Kidd S, Lass-Flörl C, Lockhart SR, Martins MA, Meis JF, Melhem MS, Ostrosky-Zeichner L, Pelaez T, Pfaller MA, Schell WA, St-Germain G, Trilles L, Turnidge J.
2012.Cryptococcus neoformans-Cryptococcus gattiispecies complex: an in-ternational study of wild-type susceptibility endpoint distributions and epidemiological cutoff values for fluconazole, itraconazole, posaconazole, and voriconazole. Antimicrob. Agents Chemother.56:5898 –5906.http: //dx.doi.org/10.1128/AAC.01115-12.
48. Varma A, Kwon-Chung KJ.2010. Heteroresistance ofCryptococcus gattii
to fluconazole. Antimicrob. Agents Chemother.54:2303–2311.http://dx .doi.org/10.1128/AAC.00153-10.
49. Malik R, Medeiros C, Wigney DI, Love DN. 1996. Suspected drug eruption in seven dogs during administration of flucytosine. Aust. Vet. J.
74:285–288.http://dx.doi.org/10.1111/j.1751-0813.1996.tb13776.x.