Copyright 0 1983 by the Genetics Society of America
GENETIC METHODS FOR ANALYSIS AND MANIPULATION
O F INVERSION MUTATIONS IN BACTERIA
MOLLY B. SCHMID AND JOHN R. ROTH
Department of Biology, University of Utah, Salt Lake City, Utah 841 12
Manuscript received March 31, 1983 Revised copy accepted August 5, 1983
ABSTRACT
A number of genetic methods for the isolation, characterization and manip- ulation of large chromosomal inversions in Salmonella typhimurium are de- scribed. One inversion-carrying mutant is characterized in detail and used to demonstrate a number of unique genetic properties of bacterial inversions. 4 o n t r a r y to expectation, it was found that large inversion mutations can be repaired by generalized transduction. The repair results from the simulta- neous introduction of two wild-type transduced fragments into a single recipi- ent cell. Homologous recombination between the two transduced fragments and the two inversion breakpoints causes the inverted segment to be rein- verted. This results in regeneration of the wild-type orientation of this chro- mosome segment. Similar recombination events allow a large inversion muta- tion to be introduced into a wild-type strain; two transduced fragments from an inversion strain cause recombination events resulting in inversion of a large chromosome segment.-Genetic methods for mapping the extent of a large inversion mutation by generalized transduction are described and tested. The methods are operationally simple and allow good resolution of the two inver- sion breakpoints.
NLY a few examples of chromosomal inversions are known in bacteria.
0
T h e multitude of well-analyzed bacterial mutations makes the rarity of inversions unusual. T h e reason for this rarity is not obvious. In a simple bacterial inversion mutation, a segment of the circular chromosome is in re- verse orientation when compared with wild type. T h e inversion could naively be formed by two sites of chromosome breakage and rejoining that result in two novel junction sites or “inversion breakpoints.” T h e sequences between the two inversion breakpoints remain unaltered in a simple inversion. One would expect an inversion-carrying mutant to be viable, regardless of the site or size of the inverted region, as long as neither inversion breakpoint disrupts an essential gene. As long as such disruptable sites exist, naturally occurring homologous sequences and insertion elements could be expected to generate inversion in bacteria j u s t as they are thought to generate deletions and dupli- cations. T h e frequent isolation of transposable elements in nonessential sites suggests that a very large number of disruptable sites exist.One would expect that inversion-carrying mutants would be isolated as aux- otrophs, since an inversion breakpoint could disrupt a gene or operon. Such inversion mutants have not been found. An extensive characterization of more
518 M. B. SCHMID AND J. R. ROTH
than 1500 his auxotrophs revealed no inversions (HARTMAN et al. 1971). In- versions have not been described in the similarly extensive collections o f mu- tants in other gene systems (trp, ieu, lac, gal, hut, ara).
A few inversions have been described in bacteria; some of these involve large arcs of the chromosome and appear to be evolutionarily stable. The large inversion, by which the Escherichia coli and SalmonPlla typhimurium chromosomes differ, extends over 15% of the genetic map (CASSE, PASCAL and CHIPPAUX 1973). T h e genetic maps of these two species (including more than 1000 genes in E. coli and 700 in S. typhimurium (SANDERSON and HARTMAN 1978; BACH- MANN and Low 1980; RILEY and ANILIONIS 1978) are virtually superimposable except for this inversion. Recently, HILL and HARNISH (1981) discovered an inversion that differentiates the E. coli K-12 W3 110 strain from other E. coli
K-12 strains. This inversion extends over 20% of the chromosome, between two inversely oriented ribosomal RNA (rrn) gene clusters. Presumably, this inversion resulted from homologous recombination between two rrn sequences. T h e inversion appears stable, since strains derived from W3 1 10 also carry the inversion.
Recently, it has been shown that insertion elements cause inversion mutations with relatively high efficiency. KLECKNER, REICHARDT and BOTSTEIN (1 979) have shown that 50% of the tetracycline-sensitive mutations isolated in strains carrying the TetR determinant, TnlO, are inversions. Most of these inversions appear to be smaller than 50 kb. One breakpoint of the inversion is the site of the insertion element; the other breakpoint is presumably chosen randomly. Current transposition models propose that inversion formation and transposi- tion are alternative events (KLECKNER, REICHARDT and BOTSTEIN 1979). The reason that small inversions are preferentially isolated is not understood.
Some of the first bacterial inversion mutants isolated were the inversion Hfr’s of BERG and CURTISS (1967). These workers isolated mutants of an Hfr strain that showed an inverted direction of Hfr transfer. These inversion Hfr’s may result from recombination between insertion elements known to reside in F sequences [IS2, IS3 and IS1000
(rs)]
and insertion elements present in the bacterial chromosome. These inversions also affect fairly small chromosomal segments.T h e reason that inversions are rare among spontaneous auxotrophic muta- tions is uncertain. T h e obvious possibilities are either that inversion formation is mechanistically difficult and occurs rarely, or that inversions are formed but lead to reduced viability o r death. Since so few inversion mutations are known,
BACTERIAL INVERSION MUTATIONS ._ I9
MATERIALS AND METHODS
Bacterial strains: The genotypes and origins of strains referred to in this paper are given in Table 1. All of the strains are derived from S . typhiinurium strain LT2.
Media and growth conditions: The E medium of VOCEL and BONNER (1956) with 2% glucose was used as minimal medium. This was supplemented as needed with histidinol ( 1 mM), histidine (0.1 mM) or adenine (0.5 mM) plus thiamine (0.05 mM). The histidinol medium used for reversion of deletion strain hisOGD.203 to histidinol utilization was supplemented with all amino acids except histidine, plus nucleosides, thiamine and histidinol. DAVIS, BOTSTEIN and ROTH (1 980) described the standard concentrations of these supplements. For Hfr-mating crosses, solid minimal medium was supplemented with 10 ml of Difco nutrient broth per liter of medium.
Tryptone-based medium (8 g/liter of Bacto-tryptone, 5 g/liter of NaCI, 8.4 g/liter of glucose, 1 g/liter of yeast extract) or Difco nutrient broth with 0.5% NaCl were used as complex media. Solid media of all types contained 15 g/liter of Difco Bacto-agar. Tetracycline was added to complex medium at 15 rg/ml and to minimal medium at 10 rg/ml.
Mutant isolation: Single colonies of strain TR5992 (his-203) were isolated on complex medium, inoculated into complex liquid medium and grown at 37" with shaking. Each of these independent cultures was plated on solid E medium supplemented with histidinol, amino acids (except histidine), nucleosides and thiamine and incubated at 37". When histidinol-utilizing revertants arose (after 36-60 hr), the plates were replica plated to solid E medium supplemented only with histidinol. Any histidinol-utilizing mutants that grew poorly on unsupplemented histidinol plates were purified and retested for auxotrophy.
Transduction crosses: The high-frequency transducing phage P22 H T 1 0 5 / 1 int-201 was used in all transductions. This phage was derived by G. ROBERTS from the P22 HTlOSIl strain of SCHMIE- GER (1972). The mutant phage performs generalized transduction with increased efficiency over wild-type P22 phage. It has been estimated that 50% of the phage particles of this mutant carry transduced bacterial DNA (SUSSKIND and BOTSTEIN 1978). Standard transduction crosses were done at a multiplicity of infection between 1 and 10 at 37" unless otherwise stated.
Strain preseruation: All strains known to carry inversions, derived from inversion-carrying strains, or potentially carrying inversions were stored in 8.0% dimethyl sulfoxide (DMSO) at -70" im- mediately after purification.
Determination of the orientation of Tn 10 insertions: The method of CHUMLEY, MENZEL and ROTH
(1979) was used to determine the orientation of TnlO insertions. S. typhimurium strains are inher- ently Lac-. An F,, 114 Lac+ episome carrying a TnlO insertion [either z$-20::TnlO(A), or zzf-
2l::TnlO(B)] was introduced into a strain carrying the TnlO element the orientation of which was to be determined, by selecting Lac+ exconjugants at 30". Hfr strains were isolated from the F' Lac+ merodiploids by selecting Lac+ derivatives at 42', Such temperature-resistant survivors have been shown to be due to integration of the F' into the site of the chromosomal TnlO element by homologous recombination. This yields an Hfr whose direction of chromosome transfer is dictated by the orientation of the TnlO elements involved. The direction of chromosome mobilization of these Hfr's is determined by conjugation crosses. A chromosomal TnlO insertion that gives an Hfr with counterclockwise transfer when an F'lac plasmid carrying zzf-2O::TnlO (TT627) inserts is referred to as an "orientation A" TnlO insertion. Chromosomal TnlO's that give rise to coun- terclockwise transfer in Hfr's that are derived from zzf-2l::TnlO (TT628) are referred to as 'orientation B" TnlO insertions.
DNA preparation: A portion of the his operon including the his control region, hisG, hisD and part of hisC has been cloned onto the single stranded phage M13 by BARNES (1978, 1979). The M13Ho168 phage of BARNES was used in all of these experiments. The double-stranded replicative form of M13Hol68 was nick-translated using a''P-dATP (DAVIS, BOTSTEIN and ROTH 1980) and used as the probe in Southern blot hybridization to identify the restriction fragments containing the hisOGDC chromosomal genes.
Chromosomal DNA was isolated I-ml overnight cultures following the rapid method of DAVIS
et al. (1980). Restriction enzymes were obtained from New England Biolabs.
520 M. B. SCHMIDANDJ. R. ROTH
TABLE 1
List of strains
Strain no. Genotype Source
TT3 17 T T 6 2 7 T T 6 2 8 T T 1 3 3 3 T T 3 7 0 7 T T 3 7 0 9 T T 3 7 1 3 T T 3 7 1 5 T T 3 7 3 1 T T 3 7 3 2 T T 3 7 3 3 T T 3 7 3 5 T T 3 7 3 6 T T 3 7 3 7 T T 3 7 3 8 T T 3 7 4 0 T T 3 8 0 6 T T 3 8 0 8 TT38 10 T T 3 8 1 1 T T 3 8 1 2 TT38 17 T T 3 8 18 T T 3 8 6 8 T T 3 8 7 0 T T 3 8 7 2 T T 4 7 7 4 T T 5 3 2 4 T T 5 3 3 0 T T 6 0 6 7 T T 6 8 6 2
T T 6 8 6 4
T T 6 8 6 5 T T 6 8 6 6 TR2931 TR5835 TR5992
purFI74I::Tnl0
strA pyrC7 /F'1t4 (ts) zzf-20::TnIO (lac+) strA pyrC7 /F'114 (ts) zzf-21::TnlO (lac+)
trp-2451::TnlO his-9 70 7
purF174I: .TnlO his-9707 cysG1510: :TnlO his-9707 metAB869: : T n I0 his-9 70 7 pheA534: :Tn 10 his-9707 c)~sA1539::TnlO his-9707 argGl828: :Tn I0 hzs-970 7 metCl975: :TnlO his-9707 tyrA555: :TnlO his-9707 glyA540: :TnlO his-9707 purC882: :Tn 10 his-9707 serA977: :Tn 10 hzs-9707 ilvA2173: :TnlO his-9707 inetE866::TnlO his-9707 ilvA597: :TnlO his-9707 metE864: : T n l 0 hzs-9707 metAB89 7: : T n I O his-9707 metF877: : T n l 0 his-9707 purDl735::TnlO his-9707 leu-I 151: :TnlO his-9707 pyrB692: :TnlO his-9707 pyA684: : T n 10 his-970 7
zee-2::TnlO IN282
p u r F I 741 : : T n l 0 IiW282 his-203 trp-2451: :TnlO IiW282 has-203
zcf778::TnlO ( T n I 0 98% linked to purB) zce-862: :TnlO his-9707
zce-864: : T n l 0 his-9707
INV282 hisC869 I : :Tn IO(A) INV282 hisC8667: : T n I O(B) his-2236 his01242 edd- gnd'
INV282 his-203 his-203 Laboratory collection Laboratory collection Laboratory collection Laboratory collection Laboratory collection This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study This study
K. L. STRAUCH, C. G . MILLER K. L. STRAUCH,
C. G. MILLER This study This study
Laboratory collection This study
Laboratory collection
Multiply marked strains used in this study are listed along with their complete genotype. T h e nomenclature used t o describe inversion strains is described in MATERIALS AND METHODS. Other strains described throughout the text and tables as single auxotrophs (his-203, leu-485, his-135) carry only the mutation listed. Strains T T 6 8 6 2 and T T 6 8 6 4 are described in STRAUCH and MILLER (1 983).
dodecyl sulfate (SDS) and 100 pg/ml of salmon sperm DNA. Washes were performed at 45" in
2X SSPE plus 0.2% SDS.
BACTERIAL INVERSION MUTATIONS 521
nature is given the three letter symbol CRR and an allele number. As more becomes known about the nature of the mutation, it is redesignated as an inversion (INV), a duplication (DUP), a deletion (DEL) or a transposition (TPN). It retains the same allele number to prevent confusion, so all rearrangement allele numbers are drawn from a single pool. Capital letters are used to distinguish these numbers from standard mutational allele numbers. The first and last genes known to be inverted are enclosed in parentheses. Any standard mutations that have been introduced into the inverted region are also listed within the parentheses. Any mutations that are carried by the strain, but not within the inverted segment, are listed after the inversion description.
RESULTS
Properties of inversion mutations: T h e expected genetic behavior of a strain carrying an inversion mutation is described as follows.
1. Normal genetic linkages should be disrupted at two distinct points on the chromosome. A strain carrying an inversion has two sites where normally oriented and inversely oriented sequences are joined. Across these two break- point sites, normal genetic linkages will be disrupted. Phage-mediated trans- duction provides a good means of analyzing this linkage disruption; if the size of the transduced fragment is smaller than the size of the chromosomal inver- sion, the inversion cannot be repaired by a single wild-type tranduced frag- ment. Thus, if an inversion breakpoint creates or destroys a phenotype, this damage will not be repairable by a single wild-type transduced fragment.
T h e generalized transducing phage P22 was used in these experiments. It transduces approximately 50 kb DNA fragments (EBEL-TSIPIS, BOTSTEIN and
FOX 1972). Thus, to show linkage disruption in these experiments, an inversion must be larger than 50 kb, or about 1% of the total chromosome size.
Genetic linkage of two markers located entirely within the inverted segment is expected to be normal since sequences within this region are oriented prop- erly with respect to one another. T h e exception should be near the inversion breakpoints where homology necessary for crossover events may be limited.
2. An Hfr with an origin of transfer within the inverted region should show an opposite direction of transfer. T h e opposite direction of transfer will be apparent when inheritance of markers outside of the inverted segment is ob- served; the inheritance pattern of markers within the inverted segment should be normal since the relative orientation of the Hfr origin and markers within the inverted segment has not changed.
3. Physical analysis of the two inversion breakpoint regions should demon- strate the two complementary fusion points. In these experiments, restriction digestion and hybridization to a cloned his fragment demonstrate that the his
sequences are present at two distant sites on the chromosome.
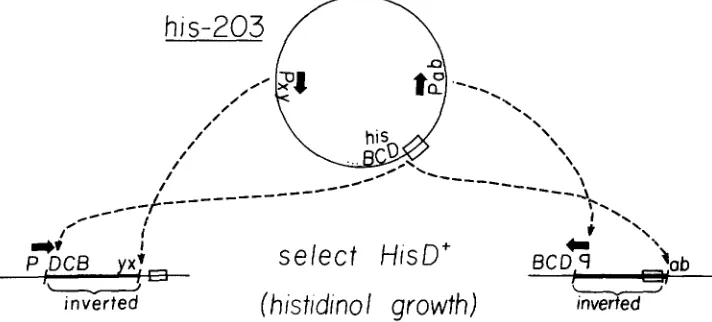
Selection of auxotrophic inversion mutations: T h e parent strain for this selection,
his-203 (TR5992), possesses an intact hisD gene that is unexpressed because the his promoter is deleted. Growth on the biosynthetic intermediate histidinol requires only a functional hisD gene, and mutants capable of histidinol utili- zation can be isolated from a strain carrying the his-203 deletion. T h e selection requires events that generate a promoter that allows expression of the hisD
522 M. B. SCHMIDANDJ. R . ROTH
first 260 base pairs of the hisG gene (S. LAM, D. BIEK and M. CIAMPI, unpub- lished results). T h e sequences in the remaining part of the hisG gene (approx- imately 650 base pairs) and the hisG-hisD intercistronic region (- 100 base pairs) are potential targets for fusion to a foreign promoter or for mutation to generate a promoter. Previous use of this selection has produced several distinct classes of mutants. Among the types of mutations isolated from the selection are deletions that extend the his-203 deletion and fuse the hisD gene to a nearby promoter, and point mutations that create a new promoter in the sequences preceding the hisD gene (AMES, HARTMAN and JACOB 1963; ST. PIERRE 1968). Unstable mutations have also been obtained from this selection; these have been characterized as tandem duplications (ANDERSON and ROTH 1978a). T h e HisD+ phenotype results from fusion of the partial
his
operon to a foreign promoter at the duplication join point.In principle, this selection should yield inversion mutations since the inver- sion breakpoint could cause fusion of hisD to a foreign promoter. Figure 1 diagrams the selection and shows two types of inversions that could lead to
hisD expression. Inversion mutations that are obtained from this selection should have one inversion breakpoint within, or very near, the his operon. T h e other breakpoint (the "non-his' breakpoint) must be near a properly ori- ented promoter to permit expression of the hisD gene across the inversion breakpoint.
A screen was designed to detect inversions in which the non-his breakpoint disrupted a biosynthetic gene or operon and caused auxotrophy. In such cases, histidinol-utilizing ( h i s D f ) mutants could be selected on medium containing histidinol and a variety of possible auxotrophic requirements. Auxotrophic inversion mutants could be identified when transferred to unsupplemented histidinol media.
BACTERIAL INVERSION MUTATIONS 523
T h e frequency of spontaneous HisD+ mutants of his-203 on supplemented E plus histidinol medium is lo-'; this frequency is not noticeably different than the frequency of mutation to histidinol growth on unsupplemented E plus histidinol medium (ANDERSON and ROTH 1978a, and M. SCHMID, unpublished results).
Out of 10,000 histidinol-utilizing mutants in 100 independent groups, only one, TR3835, has acquired a new auxotrophy. This strain requires adenine, as well as histidinol or histidine, to support growth on minimal medium. T h e requirement for adenine cannot be replaced by guanine; this growth property is characteristic of either purA or purB mutations. Analysis of this mutant has shown that the chromosome segment between
his
and purB is inverted. A number of genetic and physical properties supf*)rt this conclusion. These in- clude the demonstration that linkage is disrupted in both the his operon, and at purB; this reduced transducibility of the Pur- phenotype or the His- phe- notype; a change in the orientation of Hfr transfer within the chromosome segment between his and purB and the physical demonstration that his se- quences are separated in this strain.Linkage disruption at the inversion breakpoints: An inversion mutation that sat- isfies this selection will position a foreign promoter near the hisD gene. If the inversion breakpoint lies within the remaining part of the hisG gene, the two parts of the his operon will be separated in the inversion chromosome. A wild- type transduced fragment will be unable to repair the his operon to give a His' phenotype if the inverted segment is larger than a transduced fragment. Table
2
shows that strain TR5835 is transduced to His+ at an extremely low frequency. This is unlike the parent strain, his-203, and most of the other mutants isolated from this selection that are readily transducible to His+.Likewise, if the inversion breakpoint disrupts the PurB gene, the strain should not be transducible to Pur+. Data in Table
2
show that the Pur- phenotype of strain TR5835 is repaired at very low frequency by transduction. Thus, both phenotypes that were obtained in the mutational event yielding strain TR5835 are only rarely repaired by transduction. T h e nature of the rare transductants will be discussed later.T h e HisD+ phenotype of strain TR5835 cannot readily be inherited by a strain with a wild-type chromosome order in a generalized transduction cross. Data in Table 3 show that the HisD+ phenotype of TR5835 cannot readily be donated to the parent strain, his-203. T h e rare HisD+ recombinants arise by unique recombination events that are discussed extensively in a later section. Other markers (such as leu-485) as well as other regions of the his operon (his- 135) are donated readily in a transduction cross. This property is expected if an inversion breakpoint lies between the hisD gene and the promoter express- ing it in strain TR5835. Most HisD+ mutants from this selection (carrying a point mutation, deletion or tandem duplication) can readily donate their HisD+ phenotype. Only large inversions and large nontandem duplications should be unable to donate the HisD+ phenotype to the parent his-203 strain.
5 24 M. B. SCHMID AND J. R . ROTH
TABLE 2
INV282 as a transductional recipient
Chromosome Selected No. of
Recipient order Phenotype marker transductants
his-203 Normal His- His+ 4000
p u r B l 2 Normal Ad- Ad' 3000
TR5835 INV282 His-, Ad- His+ 4
TR5835 I i W 2 8 2 His-, Ad- Ad+ 5
A P22 lysate of strain LT2 was used as a transduction donor. The number of His+ or Ad+ transductants obtained using recipients carrying the his-203 or purB12 mutations provides a meas- ure of the rarity of His' or Ad+ transductants that arise in strain TR5835. Strain TR5835 shows normal phage sensitivity and is transduced for other markers at a normal frequency. These trans- ductions were performed using 0.4 plaque forming units per cell.
TABLE 3
INV282 as a transductional donor
Donors (HisD+)
Recipient Select TR5835 (IAVZ82) LT2
hisOGD203 HisD+ 3 2000
hisIE1?5 His' 300 2000
leu-485 Leu+ 1000 2000
Strain TR5835 carries the HisD' Ad- inversion IW282. Lysates of strain TR5835 and wild- type strain LT2 were used to donate their HisD+ phenotype to recipient his-203, their hisl and hisE genes to recipient his-135 or their leu genes to recipient leu-485. Transduction of his-20? to HisD' requires a rare two-fragment event when inversion strain TR5835 is used, because the donor inversion join point must be inherited. Deletion h i d E l 3 5 shows a reduced number of transductants, because recombination must occur in the restricted region between the recipient hislE135 deletion and the donor inversion breakpoint. Mutation leu-485 is unlinked to either the his or purB regions, and the number of transductants demonstrates the transduction ability of the two lysates.
placed with wild-type his sequences and the closely linked zee-2: :TnlO insertion. If the original rearrangement has not deleted his material, but has a breakpoint within IiisG, the resulting strain, TT4774, should possess all sequences of the
hisG gene, although not in a contiguous region of the chromosome. T h e strain (TT4774) retains an Ad- Hol+ His- phenotype. T h e IiW282 rearrangement results in location of the upstream part of the hisG gene and the zee-2::TnlO insertion near the normal position of the PurB locus. T h e distal portion of the
hisG gene should be at the normal chromosome site of the his genes. When inversion strain TT4774 was used as a donor, recipient hi& point mutations were transduced to His+. Cotransduction of the hisG point sites and the donor zee-2::TnlO insertion was then determined. In a normal strain, all hisG point mutations and the zee-2: :TnlO element are 50-80% cotransducible. In the ZAV282 chromosome, some points in the hisG gene will be transductionally linked to the zee-2: :TnlO insertion, whereas other hisG sites will be far removed on the chromosome and should no longer show transductional linkage to zee-
2::TnlO. T h e inversion breakpoint was identified as the position in the hisG
BACTERIAL INVERSION MUTATIONS 525
TABLE 4
Mapping the his breakpoint of INV282
Donor strain: TT4774 (ZNv282, zee-2::TnlO) Recipient No. of transductants ?6 TetR transductants
hisG858 300 50%
hisG27 76 200 50%
30%
-
57
hisG936
-
hisG428 12 0%
hid460 150 0.67% (1/150)
hisG325 200 0%
-~ ~
Strains carrying each of the hisG point mutations listed in the first column were transduced to His’ with phage grown on strain TT4774 (IhV282, zee-2::TnlO). The percentage of TetR trans- ductants was then determined. The results show the position in hisG where (cotransduction of the zee-2::TnIO insertion and hisG sequences stops in the IhV282 chromosome. The point mutations in hisG are listed in the order in which they appear on the genetic map of the normal his operon, with the last mutations near hisD. The zee-2::TnlO insertion is located outside the his operon, near hiso. Many other hisG point mutations were tested in this experiment and confirm these results. When a lysate of strain TT4774 was used, all hisG point mutations proximal to hisG936 showed linkage to ze-P::TnlO, whereas all point mutations distal to hisG428 showed complete absence of linkage.
hisG point mutation. Table 4 shows part of the data from this experiment. T h e sequences found in strain TT4774 could repair all hisG point mutations that were tested. We conclude that no large deletion of his sequences resulted from the Ziw282 mutation. T h e zee-2::TnlO element is no longer linked to
hisG point mutations distal to mutation hisG936. We conclude that the inver- sion breakpoint is within the hisG gene between mutations hisG936 and his- G428.
Hfr
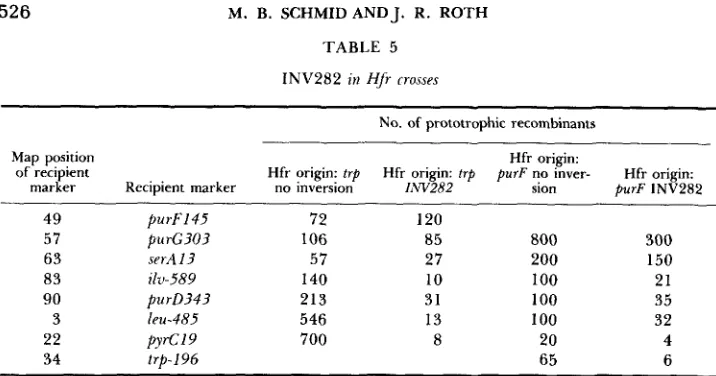
transfer between his and purB is inverted: Hfr strains carrying a chromo- somal inversion should show an order of gene transfer that reflects the new chromosome structure. T h e method of TnlO-directed Hfr formation (CHUM- LEY, MENZEL and ROTH 1979) provided a number of different Hfr origin sites in the INV282 chromosome. TnlO elements were transduced into the inver- sion chromosome by homologous recombination. TnlO insertions in the trp and purF genes served as the sites for integration of F’ts 114 TnlO episomes. Strain TT5330 carries the INV282 mutation, and a TnlO insertion in the trp genes. T h e trp operon (at 34 min) lies between the his operon (at 44 min) and the purB gene (at 25 min). In a wild-type chromosome, Hfr’s derived by insertion of F’ zzf-20::TnlO into trp-2451: :TnlO show a counterclockwise di- rection of transfer; in an INV282 chromosome, they show a clockwise direction of transfer. These directions of transfer are inferred from the data on gra- dients of transmission shown in Table 5, and diagrammed in Figure2.
Three other independently derived Hfr’s were tested that had origins at other sites in the region thought to be inverted. All of these data support the inverted nature of the region between his and purB.526 M. B. SCHMID AND J. R. ROTH
TABLE 5
INV282 in HJr rrosses
No. of prototrophic recombinants
Map position
of recipient
marker Recipient marker
49 purFl45
57 purG303
6 3 serAl3 8 3 ilv-589
90 purD343
3 leu-485
22 pyrC19
34 trp-196
Hfr origin:
Hfr origin: trp Hfr origin: trp purF no inver- Hfr origin: no inversion IN11282 sion purF INV282
72 120
106 8 5 800 300
57 27 200 150
140 10 100 21
213 31 100 35
546 13 100 32
700 8 20 4
6 5 6
Hfr strains with origins at trp were formed by the integration of F’II, carrying zzf2U::TnlO into trp-2451::TnlO; these were generated in strains TT5330 and T T 1 3 3 3 which contain trp- 2451::TnIU with and without the INV282 inversion. Hfr strains with origins at purF were gener- ated by integration of F’114 carrying zzf21::TnlO into purF1741::TnlU in strains T T 5 3 2 4 and T T 3 1 7 which contain purF174I::TnlO with and without the INV282 inversion. A set of recipient strains, each carrying only the single marker listed was used in the Hfr crosses. Donor and recipient strains were plated directly on minimal selective medium.
of the region inverted by the IiW282 mutation. Table 5 shows these data. The direction of transfer of Hfr strains with origins at purF are identical in an
I,W282 and a normal chromosome. These data strongly support the conclusion that the IiW282 mutation inverts the chromosomal region between purB at 25 min, and his at 44 min on the Salmonella genetic map.
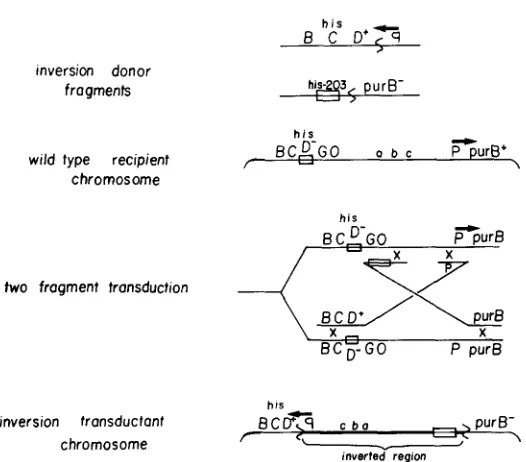
Repair ofthe inversion by transduction: At a low frequency, a wild-type donor appears capable of repairing the INV282 inversion by transduction (see Table 2). We believe that these rare transductants arise by the simultaneous inher- itance of two transduced fragments by the inversion recipient. The model for repair of an inversion by two transduced fragments is shown in Figure 3. According to this model, two wild-type transduced fragments, one from each of the chromosome regions near the inversion breakpoints, must recombine with the inversion chromosome. Recombination events must occur between each transduced fragment and the two corresponding homologous regions that are separated in the inversion chromosome. We imagine that two recipient sister chromosomes may be involved in these transductions, but other recom- bination events requiring only one recipient chromosome can equally well accommodate the events diagrammed in Figure 3. It is known that transduced fragments are capable of recombining with homologous regions that are widely separated on the chromosome (ANDERSON and ROTH 1978a,b; CHUMLEY and ROTH 1980; SCHMID 1981; and M. B. SCHMID and J. R. ROTH, unpublished results). These recombination events serve to reinvert the inverted segment and generate a wild-type chromosome (see Figure 3).
Three experiments support this model:
BACTERIAL INVERSION MUTATIONS 527
ori
porF
FIGURE 2.--Genetic maps of wild-type (WT). S. typhimurium and the INV282 mutant. The markers tested in the Hfr experiments described in Table 5 are shown. The directions of transfer of Hfr strains derived from integration of F ' I I ~ zrf20::TnIO into trp-2451::TnIO, and integration of F'114 zlf-21::TnlO, into purFl741::TnlU are shown for the two chromosomes. Hfr's with an origin of transfer in tup show opposite directions of transfer in the two strains.
mosome, there is no transductional linkage between the his and purB genes; they are separated by 20 min on the genetic map. However, when inversion strain TR5835 is transduced to His+, many of the transductants are Pur+ even though no selection was made for inheritance of the Pur+ phenotype. Likewise, when strain TR5835 is transduced to Pur+, linkage to the his genes is observed. T h e double transduction model explains the apparent linkage of these two genes in the lNV282 chromosome. In order to repair one of these genes, recombination events must involve both inversion breakpoint regions. These data are shown in Table 6. These experiments also show that the Pur- lesion of strain TR5835 is due to disruption of the purB gene. T h e donor strain carries a TnlO insertion that is 98% linked to the purB gene is a wild-type chromosome. Table 6 shows that this TnlO is inherited with high frequency when either His or Pur is repaired. This linkage demonstrates that a trans- duced fragment from the purB region is inherited in these transductants. Anal- ogous experiments with a TnIO in the purA gene show no such linkage. Data in the next section suggest that the rare His+ Ad- transductants are the result of inheritance of a single rare transduced fragment, probably one that carries a duplication join point involving the his region.
528 M. B. SCHMID AND J. R. ROTH
wild-type donor fragments
inversion recipient chromosome
inverted regim
6 C D 3
-
.,
pur6two fragment transduction
6 C D 9
his,
wild-type transductant BCDGO a 6 c P T r 6
chromosome \
FIGURE 3.-The diagram shows transductional repair of the large INV282 inversion by a donor lysate grown on a wild-type chromosome. Two wild-type donor fragments are required, one from each of the two regions disrupted by the inversion. The inversion recipient chromosome shows the ISVZ82 inversion present in strain TR5835. The his and purB regions are separated by 20 min on the genetic map. The two-fragment transduction cross is drawn using two daughter chro- mosomes. One of the recombinant products is a completely wild-type chromosome. The inversion present in the recipient has been reinverted to generate the wild-type order. The transductants that are recovered have repaired both the his and purB defects and are phenotypically Ad' His'.
fold. T h e number of His+ Ad- transductants did not increase with multiplicity of infection, suggesting that these transductants arise by a different mechanism. Similar experiments show that the frequency of His+ transductants increases as the multiplicity of infection increases. None of these data were analyzed more quantitatively since the small number of recombinants severely limits the data at low multiplicites of infection, while the complication of phage killing of the recipient cells occurs at high multiplicities of infection.
BACTERIAL INVERSION MUTATIONS 529
TABLE 6
Litiknge of PurB and His in 1NV282
~ ~
Donor: TT6067 (TnlO near pur& His+ Ad+) Recipient: TR5835 (IiW282; His- Ad-)
Selected Marker Transductant phenotypes His+
Ad+
His+ Ad'
40/50 His+ Ad+ TetR 9/50 His+ Ad+ TetS 1/50 His+ Ad- Tets
31/38 His+ Ad+ TetR 7/38 His+ Ad+ Tet'
32/41 His+ Ad+ TetR 9/41 His+ Ad+ TetS
Transductional linkage between PurB and his exists in the INV282 chromosome. A phage lysate of strain TT6067 (TnlO near purB+ in a normal chromosome) was used to transduce strain TR5835 (liw282) to either His' or Ad+ or simultaneously to His' Ad+. The unselected phenotype and the TetR or TetS phenotype were then scored.
TABLE 7
Multiplicity-depeizdetzt repair of INV282
Multi licity % Ad+ His+ Frequency of Recipient strain of ingction transductants Ad+ His+/pfu
TR5835 (INv282) 0.2 0 a . 8 x 10-9
0.4 25 4.5 x 10-9
2.1 60 18. x 10-9
2.6 95 64 x 10-9
his-203 0.02 3.7 x 10-4
0.15 5.0 x 1 0 - ~
0.7 2.0 x 1 0 - ~
2.5 2.8 x
~~ ~
A P22 lysate grown on wild-type strain LT2 was used to transduce the inversion strain (TR5835; HOP His- Ad-) to Ad+ at various multiplicities of infection. Coinheritance of His+ was then scored, and the frequency of Ad+ His+ transductants per phage plaque forming unit (pfu) was determined. The same donor lysate was used to transduce a simple his auxotroph (hzs-203) to His' at various multiplicities of infection. All of these transductants were His' Ad+ as expected.
only by inheritance of a transduced fragment from each of the two lysates. These experiments show that the transduction events leading to repair of the inversion results from simultaneous inheritance of two transduced fragments.
530 M. B. SCHMIDANDJ. R. ROTH
TABLE 8
Repnir .f INV282 by mixed donor lysates
No. of His+ recombinants obtained for indicated donors
Recipient LT2 TT609 I LT2
+
6091his-203 (His-) 30,000 (100% Tet") 0 30,000 (100% Tets)
TR5835 (IiVVZ82; 27/50 Ad- His' 28/50 Ad- His+ TetS Ad- His- Tet? 300 23/50 Ad+ His+ 0 57
I
22/50 Ad+ His+
I
45% TetS 55% TetRIPU-485 30,000 30,000 30,000
P22 phage lysates grown on strains LT2 and TT6091 [hzs-203, TnlU near purB+ (98% linked)] were used to transduce strains TR5835 (1NV282, hzs-203) and his-203 to His', and leu-485 to leu'. The IPU-485 marker is unlinked to either the his or purE region and is used to show the equal transduction ability of the lysates. Lysates of strain TT6091 are incapable of generating His' transductants in either TR5835 or his-203 because of the his-203 deletion present in all strains. The number of His', Ad+ and TetR or TetS transductants are shown. The second and third columns show the number of Hist transductants obtained when each of the lysates are used individually. The last column shows the number of His+ transductants obtained when a 1:l mixture of the lysates was used. The titers of the two lysates were made equal for the experiment. His+ Ad' TetS recombinants can arise either by using two transduced fragments from LT2 or by using one transduced fragment from TT6091 ( p u r e region fragment) and one transduced fragment from LT2 (his region fragment) and recombining such that the TnlO near pure is not inherited by the transductant. Normally, this TnlO insertion is 98% linked to purB. Since a 1:l mixture of lysates was used, one would expect about 49% linkage of TnlU to purB in this experiment. The observed linkage is 55%. The multiplicity of infection was 15 in this cross.
INV282 mutation was used to transduce a HisD- recipient strain to HisD+. Two different events could lead to HisD+ transductants. Straightforward re- combination events could repair the recipient hisD mutation. Alternatively, two fragment transduction events could lead to inheritance of the hisD+ inversion mutation; these transductants will acquire the Ad- phenotype as well. To in- crease the proportion of recombinants that inherit the
ZNV282
inversion, the recipient carried a deletion of hisD that has one endpoint close to, but not crossing, the INK282 his breakpoint. When this recipient (TR293 1) was trans- duced to HisD+, six of 11 transductants were phenotypically HisD+ Ad-, as expected of transductants that inherited the I N 2 8 2 inversion. The transduc- tant strains behaved identically with the parent strain TR5835; neither the His- nor the Pur- phenotypes was readily repaired by transduction. T h e re- maining five of 1 1 transductants were phenotypically HisD+ (His+) Ad+, as expected of strains resulting from simple repair of the recipient deletion.BACTERIAL INVERSION MUTATIONS
inversion donor frogments
wild type recipient chromosome
two frogment transduction - his
531
h i s -
inversion transductant BCD+<q c b a
-
.,
purB-chromosome - - c \
inverted region
FIGURE 4.-The diagram shows re-creation of a large inversion by transduction. A lysate grown on a strain carrying inversion :AV282 is used to transduce a HisD- recipient (with a normal chromosome order) to HisD+. The transduction requires the simultaneous introduction of two transduced fragments carrying the two inversion join points into a recipient cell. The wild-type recipient chromosome is drawn as a replicating fork for convenience. The t w o donor fragments pair at the his and purl? regions, separated by 20 min on the genetic map. The resulting recom- binant has inverted the region between his and purl?, is phenotypically HisD+ Ad- and is genetically indistinguishable from a strain carrying the original INV282 mutation.
532 M. B. SCHMIDANDJ. R . ROTH
donor fragment
his-9707
I)
Jvvv TnlO P a b c fl t z
,
recipient chromosome
mapping cross
-
hls-9707
-
his
TnlO
7
D+ c’ TnlO P a b cHis D’ transductant w w
z
dvb- -+FIGURE 5.-The location of the non-his inversion breakpoint can be determined by transduc- tion. The transduction donors carry the inversion mutations and a hisC::TnlO insertion in either “A” or ‘B” orientation. The transduction recipients carry the his-9707 deletion and a TnZO inser- tion in a known location. Each of these TnlO insertions lies either clockwise (to the left) or counterclockwise (to the right) of the promoter that expresses the hisD gene in the inversion strain. To inherit the HisD+ phenotype of the donor, recombination events between the donor and recipient foreign promoter sequences and the donor and recipient TnlO insertions must occur. In case 1, the recipient TnlO insertion is to the right (counterclockwise) of the foreign promoter. Recombination events here lead to a deletion between the foreign promoter and the TnlO inser- tion. These transductants will be inviable, so no HisD+ transductants will be recovered. In case 2,
the recipient TnlO insertion is to the left (clockwise) of the foreign promoter. Recombination events here lead to a duplication between the foreign promoter and TnlO. These transductants will be viable, so HisD+ transductants will be recovered.
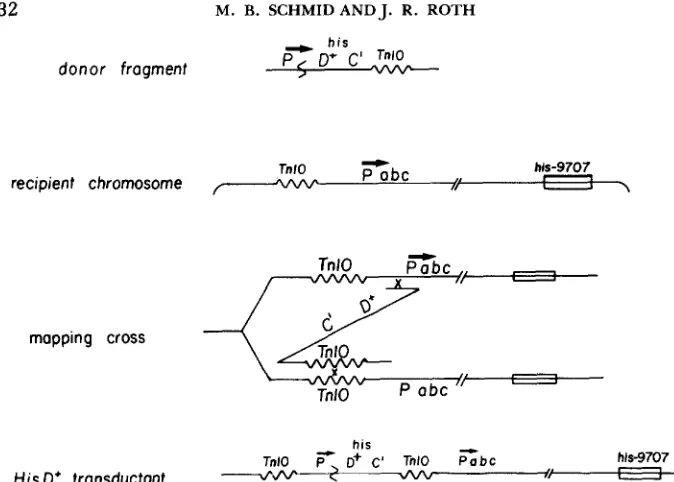
within this region, this transductant will not be viable. Thus, HisD’ transduc- tants will arise only when the TnlO and foreign promoter are in the proper location with respect to one another. In general, HisD+ transductants will arise only when the TnlU insertion is clockwise of the foreign promoter site. This provides a method of mapping the location of the foreign promoter.
Since TnlO insertions have a genetic orientation, only insertions with the same orientations will recombine readily. The donor hi&: :TnZO insertion is inverted with respect to the normal orientation of the foreign promoter. In order to get homologous recombination in this cross, the inversion donor hisC::TnlO that is known to be in “A” orientation should recombine best with a recipient TnlO known to be in “B” orientation. This helps to diagnose the donor chromosome rearrangement as an inversion.
BACTERIAL INVERSION MUTATIONS 533
R. ROTH, unpublished results). Duplication between virtually any two homol- ogous sequences has been found to occur during transduction. Even large duplications arise (duplications of up to 45% of the chromosome have been recovered). Duplication of certain regions of the chromosome leads to reduced viability of the transductants (SCHMID 1981; and M. B. SCHMID and J. R. ROTH, unpublished results).
Crosses using the INV282 inversion showed the validity of this mapping method. Lysates grown on strains containing a hisC::TnlO insertion in the INV282 chromosome (TT6865 and TT6866) were used to transduce a number of recipient strains. These recipient strains each carry a chromosomal TnlO insertion and a large deletion of the
his
operon (his-9707). Table 9 shows the results of some of these crosses. No HisD+ transductants are recovered except when the recipient TnlO lies clockwise of the purB region (in strain TT6862). These data also show that known A orientation T n l 0 insertions in the donor recombine best with known B insertions in the recipient, confirming that theINV282
mutation is an inversion.T h e duplication transductants that arise from these crosses should have a number of unique genetic properties. Because of the limited number of recip- ient strains that yield transductants in these crosses, demonstration of the genetic properties of these transductants is deferred to the accompanying paper (SCHMID and ROTH 1983).
Physical analysis of inversions: An inversion that fuses hisD to a foreign pro- moter causes the his sequences to reside at two distinct chromosomal locations. This should be observable by restriction fragment hybridization; a restriction enzyme that normally gives only one fragment able to hybridize with cloned
his sequences should give two hybridizable fragments when DNA from an inversion strain is digested. BARNES (1978, 1979) has cloned and sequenced the first part of the his operon. T h e cloned region includes the his control region, as well as the hisG, hisD and part of the hisC genes. Figure 6 shows the restriction fragments from wild-type (LT2, lane l ) , the parent his deletion strain (his-203, lane
2)
and from the inversion strain (TR5835, lane 3) that hybridize with the his clone. Both strains with wild-type chromosomes (his-203 and LT2) have a single hybridizing fragment, whereas the INV282 strain shows two hybridizing fragments. This demonstrates that the two parts of the his operon are separated by at least one restriction site. From the genetic evidence that has been presented, we believe that the two parts of the his operon are separated by 20 min on the genetic map. These two restriction fragments are genetically separated in the chromosome, as can be seen when transductional repair of the his-203 deletion causes one of the fragments to increase in size (and increase in hybridization ability), whereas the other fragment remains unchanged (Figure 6, lanes 3 and4).
DISCUSSION
534 M. B. SCHMID AND J.
E ,
w
I
?.
8s
10 g : r -z e
c 9b u k 2 b x i j
E
.s
8 -
I 1 1 I I I I
Y
2
83
k E o -
-
e.;'a
I I I I I I Iz e k g
bxc.j
~ 8 . 2
6 '
Y
;
82
VI
E3ggr
8 F L C 0 I I I I I I t I 8
,$....
afle.2 $.n $ 2
4
eG
3
8g
a B
$ g A
p g
2 %
-..t c4 c
% e
$6b z
?
2
8
s
;
f g
2
252
2
e
-
p 0 0 0=3
E 9
I I I I I I +
m "
L. -e -9
9r
8
.*
I I I I I I I
b..
5
C
0 '5
2 9 b d F 9 d b F 9
b,% h
8 %
I
0 U
.- $ 1
G
n.
s
.-
v
2.:
.g
g g
-
" o m % - &m m m e i m
r - - m c o m p * D J ,E 0 m
-
r3 w"
rgb b b b b b b b b b + k b *
j; e r - p - r - c o m w m
* * * " "
R. ROTH
g*b
4
;*'Zz
.-
* 0 . 3 m o b c3 &U5 2
3 o m s p g - U 3 0' C Z & Z5J c m
O 2. ru%$
m g
EF&.5
p+
*
.9."
-
-5 6z:
.E
g s 2
v1 p t l s 0 ,o+ e , 2 . & - O,9
ij
"C"ail,t y5z:%s2m
e,z E % b d > O.f 9:-, $ %." M I .e-.- U) e,
g . 5 - c I 2 . S b
'3 &U .a I O v1 p! 3
S
. 2 0
bo32
% *P "
c
-,p+
g
$ %;.g
E E 6 O32"
r a C13-b
G ' G S Z
5
$ % b b w.Q * c3 3 v s 5 L l -&Us
ozz
05
5
O Z C Vm * 2 m s (0
=
8 c b - b - t j m3" 3 9 & * e,=. m b
v1 c 8 I...? 3 b
:
6
$ 8
2 U) 2 . 50 0 , " cd
-
m Qh
-5%
2-3 3 2? - U 9 Q J e , o
. Y p . 5 h U 8 E.?
e ¶ s < s i g e r
g & g e r o 2 .* &U 0 - m g E -
-
ti; g . 5 0 2 52 %
e, 9 $ d2=
er2
22
2 M b m u 2U - + p u A S b
4
g n * er * , O R 'M O . " 2 5 " I
5
6 Z Z c i; 8 3;;.?Jy 2 c L, 2
g
Ll3 c 3 S ! ? o ' 2 3 er 8 9 Q &i:* u .8
5
.a5
e
j-.+ 13 2 tl
Y
;;i2
UM p ! $ Z < : 23
2
.gx
8 c M2 $ c 9
5
2 . 2 ere
E C Z i J - V L S Q V :
$532
g.3
+.%-
e, U;
2 . 2g z &
c 5
c UZ I c . ^53
z
9
yJ55
r"
9
k ; ; S q + r c-E
? e E - d E % , j $$ ?
-
fj 2g
E2.S
$6. g $ . g " u S E + I L .
-
3 %
g;
5
&'XZZ
$2
~ . g d ~ ~ ~ & U
o s 923.6*;:
g
g s
2 : ;
f s . 5 3 e, 3.5 p m 0 a
g p g s & 5 p b ; m p
$ L.-$ er a E k ; _ x* &U'ZJQ.&lS *i u13
*
e - . - Ala yt ' j o + = u.3 c
'
L, 8-6M U & ac U1
0 3 3 - &U
4
a22
!$2 S g
x A.-
3
g * -
O O k Z b ? g% . s M $ y s h m
0'
53
b c2
E
bn
BACTERIAL INVERSION MUTATIONS 535
1 2 3 4
FIGURE B.--Chromosomal DNA from (1) LT2, (2) his-203, (3) TR5835 (IW282, his-203) and (4) TT4774 (LW282, w+2:Tn10) was digested with restriction enzyme Hindlll. The digested DNA was probed with a-’*PdATP-labeled M 13Ho168. This M 13 derivative carries the h i d , G, D
and part of the hisC genes. Restriction enzyme Hindlll does not cut within the his region cloned on M 13hol68, yielding a single chromosomal fragment from wild-type strains that hybridizes with the probe. All INV282-carrying strains show two hybridizing fragments. The his-203 deletion has been repaired in strain, TT4774 (4). yielding a higher molecular weight band that hybridizes more extensively with the cloned his region.
the hisD gene. Since approximately 10% of the chromosome is occupied by genes that are dispensible on rich medium, it was thought that considerable opportunity for inversion was provided. Nevertheless, such inversion mutations were rare; only one auxotroph was recovered from 10,000 mutants isolated from this selection. This mutation, lNV282, is an inversion of the chromosome segment between the purB gene at 25 min on the genetic map, and the hi5
operon at 44 min on the genetic map.
T w o new genetic techniques for manipulating inversions resulted from the analysis of the 1NV282 inversion. One is a method to repair large inversions by transduction with two transduced fragments; by this method, a large inver- sion mutation can be removed from a strain by transduction with wild-type phage. Conversely, a large inversion can be introduced into a wild-type strain by transduction. In these experiments, a bacteriophage P22 mutant (HTI05/
536 M. B. SCHMID AND J. R. ROTH
this reason, the multiplicity of infection can be increased greatly before cell death d u e to phage infection occurs.
One reason that inversions are rare may be the need to acquire compensat- ing mutations or rearrangements. Analysis of the frequency with which an inversion is re-created by two-fragment transduction in a wild-type strain may provide a way to detect the necessity for these compensating changes. In a two-fragment transduction, the two transduced fragments cause a larger seg- ment of the recipient chromosome to invert. Any inversion that cannot be re- created, or that is re-created only at a low frequency, may be one that is associated with vital secondary mutations or rearrangements.
A technique for mapping the breakpoint of inversion mutations was also developed and tested using inversion INV282. This technique will be useful for analysis of inversion mutations that have a phenotype associated with only one breakpoint. When this method is used, the chromosomal location of the fusion between hisD and a promoter can be found by generalized transduction. Refinement of the chromosomal location of the foreign promoter seems limited only by the number and locations of TnZO insertions.
Since the important transduced fragment in mapping the non-his inversion breakpoint originates from the foreign promoter-hisD fusion region, this method analyzes only a small part of the rearranged strain. Because of this, the method can analyze any chromosome rearrangement in which fusion of distant sequences results in a selectable phenotype. T h e method also gives some information about the relative orientations of the two chromosome segments. However, because the method only maps the location of the foreign promoter, the presence of complex rearrangements will be obscured.
Altogether, these data show that inversion mutations in S. typhimurium are quite rare. Initially, we thought it possible that the absence of known inversion mutations might be d u e to some unexpected genetic properties that led to the misclassification of these mutations. However, the basic genetic behavior of
I1W282 was not surprising. This strongly suggests that other reasons prevent the more frequent isolation of mutants carrying inversions.
grant GM 07464.
This work was supported by National Institutes of Health grant GM 27068 and by training
LITERATURE CITED
AMES, B., P. HARTMAN and F. JACOB, 1963 Chromosomal alterations affecting the regulation of histidine biosynthetic enzymes in Salmonella. J. Mol. Biol. 7: 23-42.
ANDERSON, R. P. and J. R. ROTH, 1978a Tandem chromosomal duplications in Salmonella typhi-
murium: fusion of histidine genes to novel promoters. J. Mol. Biol. 1 1 9 147-166.
ANDERSON, R. P. and J. R. ROTH, 197813 Tandem genetic duplications in Salmonella typhimurium: amplification of the histidine operon. J. Mol. Biol. 126: 561-575.
BACHMANN, B. J. and K. B. LOW, 1980 Linkage map of Escherichia coli K12, edition 6. Bacteriol. Rev. 44: 1-56.
BACTERIAL INVERSION MUTATIONS 537
BARNES, W., 1979 Construction of an M13 histidine transducing phage: a single-stranded cloning
BERG, C. M. and R. CURTISS, 1967 Transposition derivatives of an Hfr strain of Escherichia coli K12. Genetics 56: 503-525.
CASSE, F., M. PASCAL and M. CHIPPAUX, 1973 Comparison between the genetic maps of Esche- richia coli and Salinonella typhimurium: length of the inverted segment in the trp region. Mol. Gen. Genet. 124: 253-257.
CHUMLEY, F., R. MENZEL and J. R. ROTH, 1979 Hfr formation directed by TnlU. Genetics 91:
CHUMLEY, F. and J. R. ROTH, 1980 Rearrangement of the bacterial chromosome using TnlU as a region of homology. Genetics 94: 1-14.
DAVIS, R. W., D. BOTSTEIN and J. ROTH, 1980 Advanced Bacterial Genetics Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
DEMEREC M., E. ADELBERG, A. J. CLARK and P. E. HARTMAN, 1966 A proposal for a uniform nomenclature in bacterial genetics. 54: 61-76.
EBEL-TSIPIS, J., D. BOTSTEIN and M. Fox, 1972 Generalized transduction by phage P22 in Salmonella typhimurium. J. Mol. Biol. 71: 433-448.
HARTMAN, P. E., Z. HARTMAN, R. C. STAHL and B. N. AMES, 1971 Classification and mapping of spontaneous and induced mutations in the histidine operon of Salmonella. Adv. Genet. 1 6
1-34.
Inversions between ribosomal RNA genes of Escherichia
Inversions and deletions of the Salmonella chromosome generated by the translocatable tetracycline resistance element TnlU. J. Mol. Biol. 127: 89-1 15.
Evolution of the bacterial genome. Annu. Rev. Microbiol.
32: 519-560.
Linkage map of Salmonella typhimurium, Ed. V.
Chromosome rearrangements in Salmonella typhimurium. Ph.D. Thesis, Uni-
Selection and endpoint distribution of bacterial inversion
Phage P22 mutants with increased or decreased transduction abilities. Mol.
Mutation creating a new initiation point for expression of the histidine
Isolation and characterization of Salmonella typhimurium
Molecular genetics of bacteriophage P22. Microbiol. Rev.
Acetylornithase of Escherichia coli: partial purification and some
Corresponding editor: E. JON= vehicle with ne EcoRl site. Gene 5: 127-139.
639-655.
HILL, C. W. and B. W. HARNISH, 1981
KLECKNER, N., K. REICHARDT and D. BOTSTEIN, 1979
coli. Proc. Natl. Acad. Sci. USA 7 8 7069-7072.
RILEY, M., and A. ANILIONIS, 1978
SANDERSON, K. E. and P. E. HARTMAN, 1978
SCHMID, M. B., 198 1
SCHMID, M. B. and J. R. ROTH, 1983
SCHMIEGER, H., 1972
ST. PIERRE, M., 1968
STRAUCH, K. L. and C. G. MILLER, 1983
SUSKIND, M. and D. BOTSTEIN, 1978 Microbiol. Rev. 42: 471-519.
versity of Utah, Salt Lake City, Utah.
mutations. Genetics 1 0 5 539-557.
Gen. Genet. 119: 75-88.
operon in Salmonella typhimurium. J. Mol. Biol. 35: 7 1-82.
mutants lacking a tripeptidase (peptidase T). J. Bacteriol. 1 5 4 763-771.
42: 385-413.
VOGEL, H. and D. BONNER, 1956