COMPLEMENTATION PATTERN CHANGES AT THE pyr-3
LOCUS OF NEUROSPORA CRASSA'
S. K. DUTTA AND V. W. WOODWARD
Biology Department, Rice University, Houston, Texas
Received April 7, 1965
H E pyrimidine-3 ( p y r - 3 ) mutants of Neurospora crassa comprise one genetic
T
locus (SUYAMA, MUNKRES and WOODWARD 1959),
fall into two complementa- tion groups when the very slow growing types are scored as nongrowers (WOOD-WARD 1962; DAVIS and WOODWARD 1962), are characterized enzymatically by
the absence of in vitro aspartate transcarbamylase (ATCase) activity or the pres- ence of ATCase activity indistinguishable, in crude extracts, from that found in wild type, and are suppressed or not suppressed by the nonallelic mutant, argi- nine-12 suppressor (arg-12s) (DAVIS and WOODWARD 1962). Mutants with ATCase activity are suppressed by arg-lZs, while mutants lacking ATCase activ- ity are not. The genetic-metabolic relationships within the arginine and pyrimi- dine metabolic pathways are as shown in Figure 1.
The current hypothesis regarding the protein structured by the pyr-3 gene is that one gene specifies the primary structure of one polypeptide and that either the tertiary or quaternary (identical subunits) structure of the protein is char- acterized by two active sites, one the transcarbamylase site and the second the site catalyzing the synthesis of carbamyl phosphate (CAP) destined for pyrimi-
arg-a
-
orn
asp
-CAP
US -UMP
(9 + ATCase
*
C& + NH3 + ATP
t
I
Pyr-3FIGURE 1 .-Gene-enzyme relationships within parts of the arginine and pyrimidine metabolic pathways. CAP = carbamyl phosphate; orn = ornithine; cit = citrulline; arg = arginine; asp = aspartic acid; US = ureidosuccinic acid, or carbamyl aspartate; UMP = uridylic acid; CPKase = carbamylphosphokinase; OTCase = ornithine transcarbamylase; nrg-3 = arginine-3 mutants.
392 S. K. DUTTA A N D V. W. WOODWARD
ATC
-
mutants (majority)FIGURE 2.-Complementation map of the pyr-3 mutants. ATC- is a symbol for the pyr-3 mutants lacking in vitro ATCase activity, and ATC+ symbolizes those mutants with in vitro ATCase activity.
dine synthesis. Supporting evidence for this view stems from the complementa- tion pattern of the pyr-3 mutants, as shown in Figure 2. All of the ATC+ mutants fall within one group and they complement two of the ATC- mutants. The majority of mutants, all ATC-, fail to complement any of the pyr-3 mutants. It was suggested (DAVIS and WOODWARD 1962; WOODWARD and DAVIS 1963) that the two complementing, ATC- mutants are defective at the transferase site, the ATC+ mutants are defective at the CAP synthesizing site, and the non- complementing mutants are defective at both sites. Evidence that the transferase site can be restored by partial reverse mutation was presented earlier (WOOD-
WARD and DAVIS 1963). This paper concerns the restoration of the CAP syn-
thesizing site by partial reverse mutation of one of the noncomplementing mu- tants. A preliminary report appeared earlier (DUTTA and WOODWARD 1964).
MATERIALS A N D METHODS
Strains: Mutants prefixed KS are described by SUYAMA, MUNKRES and WOODWARD (1959). KS125, 10, 16, U) and 48 are ATC+; each was crossed to al-2, the progeny of which yielded pyr-3 al-2 double mutants. KS6, 23 and 139 are ATC-, noncomplementing mutants; each was made doubly mutant with col-4.
The arginine-requiring mutants used in this study were argJ(33444Z), nrg-3 (30300) and RU20, a mutant isolated in our laboratory and described briefly by MCDOUGAU and WCODWARD
(1964). RU20 is located on the far right arm of linkage group I, responds in growth cultures to exogenous arginine but is leaky on minimal medium and appears to be blocked prior to ornithine in the arginine biosynthetic pathway.
Induction of partial reverse mutations: Conidia of the ATC+ al-2 double mutants were spotted (spots were '/e to 3/9 inches in diameter) onto minimal agar plates supplemented with .5 g sorbose per 100 ml. Following an LD,, of ultraviolet light (UV), conidia of the noncomple- menting, ATC-col-4 double mutants were mixed, each on a separate spot of untreated conidia. Mycelia emerging from the spots derived from contamination and partial reversion of the pyr-3 locus. Wild-type and heterocaryotic mycelia were distinguished by the following criteria: (1 )
recovery, or not, of both homocaryons; (2) reconstruction of the heterocaryon with the recovered homocaryons; (3) repetition of (1) and (2) with the reconstructed heterocaryon; (4) all hetero- caryons exhibit wild-type morphology. (All heterocaryons involved A mating type.)
Each partial revertant suspected of being functionally identical with the original ATC- complementing types was tested for its ability to suppress arg-2 and arg-3 mutants.
Crossing media All crosses were made on cornmeal agar (15 g/l) .
Enzyme assays: Mycelia grown in uridine supplemented, liquid media were harvested after
C O M P L E M E N T A T I O N CHANGES IN NEUROSPORA 393 RESULTS
Table 1 shows the attempts, and the successes, made to alter by UV irradiation three noncomplementing mutants to complement various of the ATC+ mutants. Only KS6 and KS23 yielded such mutants, and these only when mixed with conidia from KS125. I n a n earlier report (WOODWARD and DAVIS 1963), it was demonstrated that KS23 could be changed by mutation from a noncomplement- ing type to an ATC+ type. The new mutants reported here were recovered in a like manner, except that the mixed inoculum included ATC+ types rather than KS43, a representative of the ATC-, complementing type.
One each of the two kinds of heterocaryons (KS6,,
+
KS125 and KS23,,+
KS125) was tested for the recovery of homocaryons; the results are summarized in Table 2. ( p r signifies partial revertant.) The nuclear ratios of the hetero- caryons changed with time, but there is little doubt that the homocaryons derive from real heterocaryons.I n an attempt to show whether or not the new homocaryons (called KS6,,
TABLE 1
Number of heterocaryons recouered from plates mixedly inoculated with conidia from ATC+ mutants (doubly mutant with al-2) and UV irradiated conidia of noncomplementing
mutants (doubly mutant with col-4)
Strains
Irradiated Nonirradiated
Number of Number
heterocaryotic of mixed colonies recovered spots sampled
KS6 col-4
+
KS16 al-2 KS6 col-4+
KS10 al-2 KS6 col-4+
KS20 al-2 KS6 col-4+
KS48 al-2 KS6 col-4+
KS125 al-2KS23 col-4
+
KS16, 10,20 and 48 al-2 KS23 col-4+
KS125 al-2KS139 col-4
+
all five al-2 strains540 540 540 54.0 540 1080 each
810 540 each
TABLE 2
Homocaryons deriued from two heterocaryons
Heterocaryon
Irradiated Nonirradiated
KS6 col-4 f KS155al-2
KSW col-4
+
KS125 al-2Sample
1 2 3
1 2 3
Number of homocaryons
colonial albino
61 41
10 7
15 3
51 12
56 8
1 0
Number of
wild-type colonies Total colonies
tested
93 7 2 157 97 79 59
195 89 175
160
143 6 0
394 S. K. DUTTA A N D V. W. WOODWARD
TABLE 3
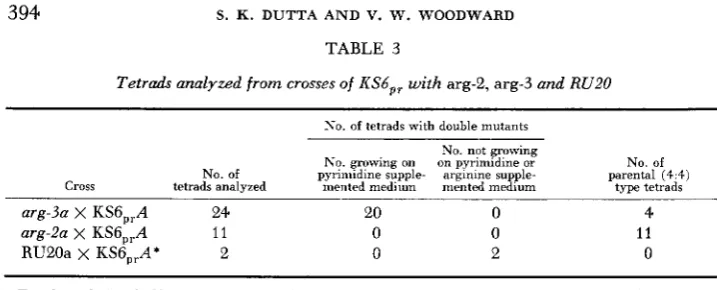
Tetrads analyzed from crosses of ICS6,, with arg-2, arg-3 and RU20
No. of tetrads with double mutants
No. not growing
No. growing on on pyrimidine or No. of No. of pyrimidine supple- arginine supple- parental (4.4)
Cross tetrads analyzed mented medium mented medium type tetrads
arg-Sa X KS6,,A 24 20 0 4
arg-2a x KS6,,A 1 1 0 0 11
RU20a x KS6,,At 2 0 2 0
Tetrads producing double mutant progeny able to grow on pyrimidine supplemented medium are evidence for s u p
* Since the double mutants from this cross require both arginine and pyrimidine, neither mutant suppresses the other. pression of arg-3 and arg-2 by KSGp,.
and KS23,,) are functionally similar to KS43, one of the complementing, ATC- strains, the following tests were made: ( a ) All the new mutants were tested for
in vitro ATC activity. Both KS6,, and KS23,, were ATC negative. (b) KS6,, was crossed to arg-2, arg-3 and RU20. (KS43 suppresses the arginine requirement of
arg-2 and arg-3
,
while RU20 suppresses the pyrimidine requirement of all ATC+ mutants.) The results are seen in Table 3. KS6,, suppresses arg-3 as does KS43; since the pyr-3 locus and arg-2 are closely linked, few recombinants are expected and none was found. RU20 fails to suppress KS6,,, further indicating its likeness to KS43.The fact that KS125 was the only ATC+ mutant with which the partial re- vertants at first complemented motivated attempts to determine whether or not the mutation involved the restoration of function or merely simulated such an effect. All of the pyr-3 mutants have been outcrossed to shed heterocaryon in- compatibility genes, but on forming double mutants with al-2 and col-4, new in-
T A B L E 4
Growth, in mm, of various heterocaryons cultured in race tubes. The heterocaryons were formed with and without al-2 as a marker gene
Days after inoculation
Heterocaryon 2 4 6 8 10
KS125
+
KS43 KS125+
KS43 al-2 KS125 al-2+
KS43 KS125 al-2+
KS43 al-2 KS48+
KS43KS48
+
KS43 al-2 KS48 aI-2+
KS43 KS48 al-2+
KS43 al-2 KS10'+
KS43 KSlO+
KS43 al-2 KSlO al-2+
KS43 KS 10 al-2+
KS43 al-2+
230tr
+
+
270+
155tr 20
0 tr
0 0
0 0
+
170+
1 4 00 0
0 0
430 6 0 505 300 110
+
0 10 390 305 0 30
230
655 845 420 545 320
+
1000 tr
90 257 560
0 tr
170 340
COMPLEMENTATION CHANGES I N NEUROSPORA 395
TABLE 5
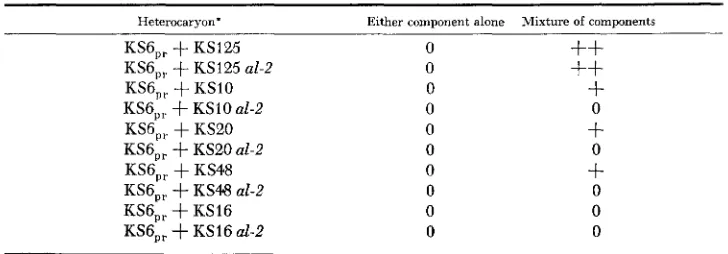
Heterocaryon formation between partial revertant, KS6,,, and ATC+ mutants with and
without the al-2 marker. The scoring was done 10 days after inoculation
Heterocaryon.
KS6,,
+
KS125 KS6,,+
KS125 al-2 KS6,,.+
KSIO KS6,,+
K S 1 0 a l - 2KS6,,
+
KS20 KS6,,+
KS20 al-2KS6,,
+
KS48 KS6,, f KS48 al-2 KS6,,+
KS16 KS6,,+
KS16 d - 2Either component alone Mixture of components
0
++
0
++
0
+
0 0
0
+
0 0
0
+
0 0
0 0
0 0
* Essentially the same results were obtained with KS23,,.
compatibility (PITTINGER and BRAWNER 1961 ; WILSON, GARNJOBST and TATUM 1961 ) genes may have been introduced. Further testing of the mutants involved revealed: (1) With the exceptions of KS125 a2-2, none of the ATC+ al-2 mutants formed heterocaryons with KS43. (2) All of the ATC+, nonalbino mutants formed vigorous heterocaryons with KS43. (3) KS43 al-2 formed vigorous there- after heterocaryons with KS125 and KS48. (4) When all of the strains were marked with al-2, KS43 formed heterocaryons with all ATC+ mutants. These data are summarized in Table 4, and point to the conclusion that incompatibility factors alter the complementation data.
KS6,, and KS23,, formed heterocaryons (Table
5 )
with KS10, KS20 and KS48,in addition to KS125. The failure of KS6,, and KS23,, to form heterocaryons with KS16 was probably due to undetermined incompatibility factors.
DISCUSSION
The data presented in this paper support the idea that the pyr-3 locus specifies the structure of a protein with two active sites, one the aspartate transcarbamy- lase site and the other a carbamyl phosphate synthesizing site. The support is interpreted to mean that the pyr-3 mutants possess proteins with one, the other, or both inactive sites, and that mutants with both sites inactive have been par- tially reverted to the extent that one or the other site is repaired. The data pre- sented here indicate that the CAP synthesizing site of the protein has been re- stored leaving a defective transferase site on the protein that prior to partial reverse mutation was doubly defective.
Evidence for the above conclusion consists of the following: the mutants used as starting material in this study are noncomplementary, ATC-, and they do not suppress arg-3. Following irradiation and selection, mutants recovered from the starting material are complementary with ATCf strains, and they do suppress
arg-3. These are the traits which distinguish KS43, one of the mutants described as lacking transferase activity on the ATC protein.
3 96 S. K. DUTTA AND V. W. WOODWARD
possessing the ability to complement ATCf mutants fall into at least four com- plementation groups. His mutants were isolated following a different selection procedure, and they were recovered in a microconidia1 strain. This phenomenon had not been observed in our studies, but until recently only two such mutants were available. The new partial revertant, which is characterized by possessing the qualities of KS43, shows complementation with KS43. This fact complicates, but does not negate, our model of the ATC protein. If the model of the protein is correct, complementation between mutants with different defective enzyme sites implies a different mechanism of complementation than that which would ac- count for heterocaryon formation between mutants within the KS43 group. An enzyme made up of identical subunits might function if one or more of the sub- units possessed one functional site, and one or more possessed the other functional site. However, the mechanism of complementation between mutants with the same defective subunit is more difficult to rationalize since one must postulate some form of self corrective mechanism between subunits.
SUMMARY
Mutants at the pyr-3 locus fall into two complementation groups: the aspar- tate transcarbamylase positive and the ATCase negative groups. This pattern of complementation, coupled with a great deal of genetic evidence, has been in- terpreted to mean that the enzyme structured by the pyr-3 gene possesses two functional sites, one the ATCase site and the other the carbamyl phosphate synthesizing site. Noncomplementing mutants are defective at both sites, the ATCase positive type is defective at the CAP-synthesizing site, and the ATCase negative type is defective at the transferase site. Evidence for this conclusion is provided by the observation that noncomplementing mutants can be partially reverted on one or the other of the two complementing types.
LITERATURE CITED
DAVIS, R. H., 1960 An enzymatic difference among pyr-3 mutants of Neurospora crassa. Proc. Natl. Acad. Sci. U.S. 46: 677-682.
DAVIS, R. H., and V. W. WOODWARD, 1962 The relationship between gene suppression and aspartic transcarbamylase activity in pyr-3 mutants of Neurospora. Genetics 47: 1075-1083. DUTTA, S. K., and V. W. WOODWARD, 1964 The alteration of a complementation pattern among
pyr-3 mutants of Neurospora. Neurospora Newsletter 6: 13.
MCDOUGALL, K. J., and V. W. WOODWARD, 1964 Suppression of pyr-3 mutants of Neurospora. Neurospora Newsletter 6: 14.
PITTINGER, T. H., and T. G. BRAWNER, 1961 Genetic control of nuclear selection in Neurospora heterokaryons. Genetics 46: 164.5-1663.
REISSIG, J. L., 1963 Spectrum of forward mutants in the pyr-3 region of Neurospora. J. Gen. Microbiol. 30: 327-337.
SUYAMA, Y., K. D. MUNKRES, and V. W. WOODWARD, 1959 Genetic analysis of the pyr-3 locus of Neurospora crassa. The bearing of recombination and gene conversion upon intrallelic linearity. Genetica 30: 293-31 1.
Heterokaryon incompatibility in Neuro- spora crassa: microinjection studies. Am. J. Botany 4.8: 294-305.
Complementation and recombination among pyr-3 heteroalleles of Neurospora crassa. Proc. Natl. Acad. Sci. U.S. 48: 348-356.
Co-ordinate changes in complementation, suppression and enzyme phenotypes of pyr-3 mutant of Neurospora crmsa. Heredity 18: 21-25.
WILSON, J. F., L. GARNJOBST, and E. L. TATUM, 1961 WOODWARD, V. W., 1962