EFFECTS OF DOSE ON THE INDUCTION OF DOMINANT-LETHAL MUTATIONS AND HERITABLE TRANSLOCATIONS WITH
ETHYL METHANESULFONATE IN MALE MICE1
W. M. GENEROSO, W. L. RUSSELL, SANDRA W. HUFF,
SANDRA K. STOUT A N D D. G . GOSSLEE2
Biology Division, Oak Ridge National Laboratory, Oak Ridge, Tennessee 37830
Manuscript received January 9, 1974
ABSTRACT
Genetic damage by ethyl methanesulfonate (EMS) in male mice was measured a t doses ranging from 50 to 300 mg/kg with dominant-lethal mu- tations and reciprocal translocations as endpoints. No appreciable increase
in dominant-lethal mutations was detected following a dose of 100 m g h g . Dominant lethals induced b y EMS were convincingly detected only after a dose of 150 mg/kg, but in the translocation experiment an increase i n the genetic effect was detectable at the 50 rng/kg dose. It is likely that dominant lethals had also been induced at the 50 and 100 m g h g doses, but were no€ detected due to the relative insensitivity of the dominant-lethal procedure. Thus, for detection of low levels of EMS-induced chromosome breakage, trans- locations are a much more reliable endpoint than are dominant-lethal muta- tions. A procedure for large-scale screening of induced translocations is de- scribed.-The dominant-lethal dose-response curve, plotted on the basis of
living embryos as a percentage of the control value, is clearly not linear as it
is markedly concave downward. Similarly, the translocation dose-response curve showed a more rapid increase in the number of translocations with dose than would be expected o n the basis d dosesquare kinetics. It is clear f o r both of these endpoints that the effectiveness of EMS in inducing chromosome breakage is proportionately much lower at low doses.
I N T R O D U C T I O N
EXPOSURE of human populations to chemicals occurs predominantly at chronic levels. For a proper evaluation of the genetic hazards to humans from chemicals, it is necessary to study in mammals the extent to which a variety of mutagenic chemicals differ in their efficacy at low, as well as high, doses. I n other words, what are the shapes of the dose-effect curves for a variety of chemicals?
In
the case of mammalian germ cells, information on this question is meager. The
present study is the first in a series to test certain known mutagenic compounds extensively over a range of doses.
The study of chemical induction of chromosomal aberrations in the mouse is important in view of the many human genetic defects that result from various chromosome anomalies. So far, evaluation of chemica1;y induced chromosomaI 1 Research jointly sponsored by the National Center for Toxicological Research and by the United States Atomic
a Mathematics Research Staff, Computer Sciences Division, Union Carbide Corporation, Nuclear Division. Energy Commission under contract with the Union Carbide Corporation.
742 w. M. GENEROSO et al.
aberrations as genetic hazards has been made primarily from dominant-lethal tests in mice or rats and from cytogenetic studies of somatic and germ cells of certain mammals. Although useful, these two test systems do not measure trans- missable genetic effects, and it is not known what their results mean in terms of hazards. Obviously, the most important mutagenic effects are the permanent, transmissable ones. Thus, there is a need to obtain information on heritable chromosomal aberrations that directly represent hazards. An important aspect of the present study is that it permits comparison of the effects of different doses of
EMS
on the induction of dominant lethals on the one hand and heritable re- ciprocal translocations on the other, and thus a direct cotmparison olf their relative efficiency in measuring induced chromosomal aberrations.MATERIALS A N D M E T H O D S
Twelve-week-old (101
x
C3H)Fl male mice were injected intraperitoneally with either a 50, 100, 150, 200, 250, or 300 mg/kg dose of EMS in Hanks’ balanced salt solution (HBSS). Control mice were given a comparable volume of HBSS. The frequencies of dominant-lethal mutations were measured in germ cells treated as early spermatozoa and late spermatids, while heritable translocations were measured only in germ cells treated in the early spermatozoa stage.In the dominant-lethal experiment, males were mated with (101
x
C3H)F, virgin females dur- ing 6.5-9.5 days after injection with EMS. The females were killed during pregnancy for uterine analysis. In the translocation experiment, males were mated with normal (SECx
C57BL)F, females during 6.5-7.5 days after injection and F, male offspring were tested for translocation heterozygosity. Random-bred T-stock females were also used in limited numbers to produce F,progeny.
F, males were tested for complete or partial sterility by caging each one with a IO- to 12-
week-old (SEC
x
C57BL)F1 female that was allowed to produce at least four litters. The litters were examined soon after birth and discarded immediately. Since the majority of females mate immediately after parturition, the litters were spaced about 20 days apart. The criteria used for full fertility of F, males were arbitrarily drawn on the basis of previous knowledge of the fer- tility of normal (SECx
C57BL)F1 females (GENFXOSO, STOUT and HUFF 1971). All F, males were declared fully fertile if a t least three litters out of four were comprised of 10 or more young,or if the total number of young for the four litters was 40 or more. F, males that were either sterile o r exhibited fertility lower than the above criteria were further tested by mating each one to three (C3H
x
C57BL)F, females. These females were killed 17 days after putting them with the male. If none of the three females was pregnant, sterility of the male was c o n f i i e d . Ifless than all three were pregnant, or if one or more females were in the early stages in pregnancy, in which classification of embryos is difficult, additional females were added. A minimum of three pregnancies was analyzed before a decision on fertility was made. The criteria used for full fertilitJr of F, males in this test was also drawn arbitrarily. These were based on the r e p m ductive nature of (C3H x C57BL)Fl females mated with normal (101 x C3H)F, males (Table 1). F, males were given a t least three more females to have a total of six analyzed pregnancies if, among the first three pregnancies, (a) all had one or more dead implants, (b) two of the pregnancies had a total of four or more dead implants, or (c) a t least one pregnancy had three or more dead implants. Otherwise, the test males were classified fully fertile an the basis of the three analyzed females. Where six pregnancies were analyzed, a n unambiguous determination
of partial sterility has always been possible. Males confirmed as partially sterile were progeny- tested and finally killed, the testes were weighed, and cytogenetic analysis of diakinesis and metaphase I spermatocytes was made.
Sterile males were killed soon after confirmation. The two epididymides were examined for presence and quality of sperm and the testes were weighed. The testes of the first four sterile F,
E M S DOSE E F F E C T S O N ABERRATIONS
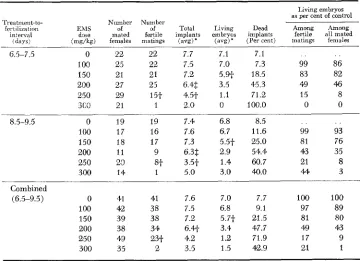
TABLE 1
Reproduction data on normal (C3H
x
C57BL)F, females mated t o normal (IO1 X C3H)F, males'743
Number of embryos Number of Frequency
per litter females (Per cent)
A. Live 6 7 8 9 10 11 12 13
1 44 Total 2 5 18 50 68 51 37 14 1 246 0.8 2.0 7.3 20.3 27.6 25.7 15.0 5.7 0.4 99.8
B. Dead 0 1 2 3 Total 145 73 25 3 245 58.9 29.7 10.2 1.2 1 w.0
* Females were killed for uterine analysis 12 to 15 days after observation of vaginal plug.
metaphase I cells or stoppage of spermatogenic development at earlier meiotic stages. Cytogenetic analysis of sterile males was, therefore, dropped in most cases. In a few instances where testes weight was normal and there was a n abundance of sperm in the epididymis, the testes of sterile
F, males were prepared for cytogenetic analysis. Male and female parents of sterile and par- tially sterile F, males in the controls and in the group exposed to an EMS dose of 50 m g h g were progeny-tested to ensure against the possibility of preexisting translocations. At higher EMS
doses, progeny-testing of parents was not deemed necessary because the incidence of induction was high.
R E S U L T S
E M S dose effect on the induction of dominant lethals: The induction of dom- inant lethal mutations at various doses of
EMS
was determined at spermatogenic stages that were found in previous studies to be most sensitive to EMS(EHLING,
CUMMING and
MALLING
1968;CATTANACH,
POLLARD andISAACSON
1968; GENE- ROSO andRUSSELL
1969), i.e., 6.5-9.5 days after treatment. The 6.5- to 7.5-day period corresponds to treated young spermatozoa while 8.5-9.5 days correspond mainly to treated late spermatids (OAKBERG 1960; OAKBERG andDIMINNO
744 w. M. GENEROSO et al.
pregnant females only gives a closer estimate of the true dominant-lethal effects. On the other hand, for the two high doses (250 and 300 mg/kg) where high dom- inant-lethal effects led to preimplantation embryonic death and consequent classification of the majority of mated females as sterile, the average numbers of living embryos based on all mated females obviously give a closer estimate.
The induction of dominant-lethal mutations was generally similar at the two mating intervals throughout the
EMS
dose range used (Table 2). I n both stages, the lowest dose that induced a detectable increase in dominant-lethal mutations was 150 mg/kg and the frequencies of dominant-lethal mutations were com- parable at each dose, as indicated by significant reductions in the number of living embryos accompanied by increases in the frequency of dead implantations. The dose of 250 mg/kg appears to be the saturation point above which the dominant- lethal-eff ect determination is no longer accurate.For the determination of the dose-effect curve, results for the two mating intervals were combined and the value calculated for the dose of 300 mg/kg was not included. The value used for the 250 mg/kg dose was obtained by taking all mated females (fertile and sterile) into consideration, while for all lower doses the values obtained on the basis of fertile matings only were used. As in the separate mating intervals, no appreciable increase in dominant-lethal mutations
TABLE 2
Effect of E M S dose on the induction of dominant lethal mutations
Living embryos as per cent of control
f n t i l i r a t i i m EM s of of Total Living Dead Among Among interval dose mated fer!ile implants embry;s implants fertile all mated
(days) (mg/kg) females matings (avg) * (avg) (Per cent) matlngs females Treatment-to- Number Number
6.5-7.5 0 92 e2 7.7 7.1 7.1 . . . .
100 23 22 7.5 7.0 7.3 99 86
150 21 21 7.2 5.w 18.5 83 82
200 27 25 6.48$ 3.5 4'5.3 49 46
250 29 15t 4 3 1.1 71.2 15 8
3 C'3 21 1 2.0 0 100.0 0 0
8.5-9.5 0 19 19 7.4 6.8 8.5 . . . .
100 17 16 7.6 6.7 11.6 99 93
150 18 17 7.3 5.5t 25.0 81 76
200 11 9 6.3$ 2.9 54.4 43 35
25 0 23 8.t 3.5t 1.4 60.7 21 8
300 14 1 5.0 3.0 40.0 4 4 3
(6.5-9.5) 0 41 41 7.6 7.0 7.7 1" 100
100 42 38 7.5 6.8 9.1 97 89
150 39 38 7.2 5.7f 21.5 81 80 200 38 34 6.4t 3.4 47.7 49 43
250 43 23f 4.2 1.2 71.9 17 9
300 35 2 3.5 1.5 42.9 21 1
Combined
* Per fertile matings.
t P
<
0.01.EMS DOSE EFFECTS O N ABERRATIONS 745
100
EO
J 0 K
+
z 0U
0 lL 6 0
8
-
tn
0
>
K
*
405
W
z_
2
J
20
0 I I I I I
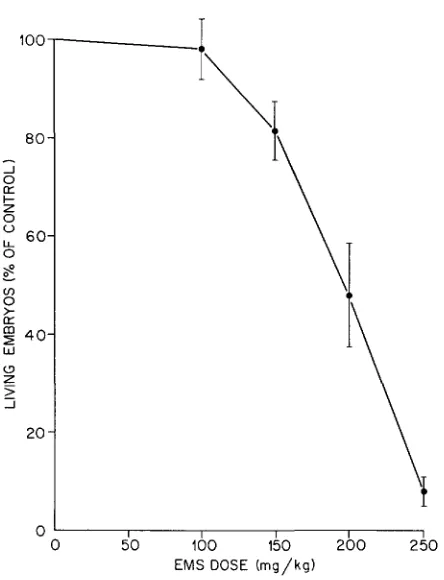
0 5 0 100 150 200 250 EMS DOSE (mg/kg)
FIGURE 1 .-Dose-response curve for EMS-induced dominant-lethal mutations. 95 % confi-
dence intervals are indicated by vertical bars.
can be detected at the dose of 100 mg/kg. Increase in induced dominant-lethal mutations was first detected at 150 mg/kg and thereafter the effect increased very sharply with dose. The dominant-lethal dose-response curve is clearly not linear-it is markedly concave downward (Figure 1 )
.
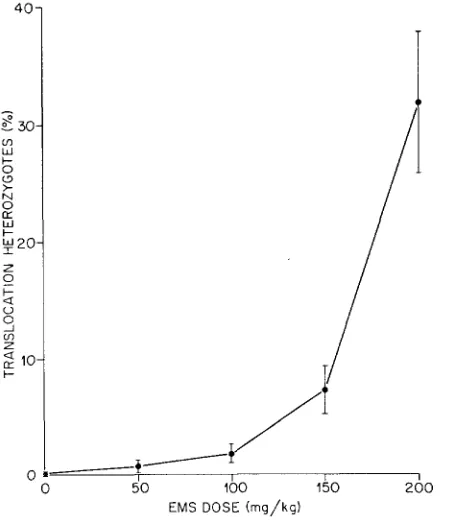
EMS dose efJect on the induction of translocations: In contrast to dominant- lethal mutations, a significant increase in the frequency of translocations was already detectable at the 50 mg/kg dose (Table 3 ) , but like dominant lethals the
TABLE 3
Effect of EMS dose on the induction of reciprocal translccations
Number of translocations
EMS dose Number of Partially Frequency
(mg/kg) Fl males tested sterile Sterile Total (Per cent)
0 1218 0 1' 1 0.08
50 853 2 4 6 0 . 7 w
100 1013 8 10 18 1.78
150 621 36 9 4'5 7.25
200 24.6 52 27 79 32.11
* T h e testes weight of this animal is 0.M grams.
746 w. M. GENEROSO et al.
frequency increased sharply and nonlinearly with dose. The translocation dose- response curve (Figure 2) showed that there is a more rapid increase in the number of translocations with dose than would be expected on the basis of dose- squarekinetics (P forlackof fit,
<
0.01, ( ~ ~ ( 3 d . f . ) = 16.2).A total of 148 EMS-induced translocations were recovered. Of these, about one- third (50) were completely sterile and two-thirds (98) were partially sterile. The distribution of sterile and partially sterile translocations at each dose is given in Table 3 . There are slightly more sterile than partially sterile
F,
males in the50 and 100 mg/kg doses although the numbers are low and the differences are not significant. On the other hand, there were significantly more partially sterile than sterile males at the 150 and 200 mg/kg doses (P for difference
<
0.01 in both doses).The great majority of male sterile translocations had distinctly small testes. Forty-eight out of 50 sterile males were so classified, and the 44 whose testes weights were obtained had weights of the two testes ranging from 0.03 to 0.12 gram (Table
4).
Pourteerr of the steriles had very few sperm in the epididymis while in 34 no sperm were observed. Among those with few sperm, most of the sperm were nonmotile and in many cases a high frequency of sperm with bent tail or other abnormality was observed. The other two sterile F, males had normal testes weights (0.22 and 0.24 gram) and many normal-looking sperm were found in the epididymis. Cytogenetic analysis confirmed that these two males were translocation heterozygotes./
FIGURE 2.-Dose-response curve for EMS-induced reciprocal translocations. 95% confidence intervals are indicated by vertical bars. Broken line shows the curve, Pi = 0.03 f 3.22 X Ik4Di2,
E M S DOSE E F F E C T S O N A B E R R A T I O N S
TABLE 4
Testes weight of EMS-induced sterile and partially sterile translocations
74 7
Class
Number of Average testes Standard
Fl males weight* (grams) deviation
Sterile 44.f .07 .020
Partially sterile 943
.w
.MNormal 24 .22 .@AI
* Weights are for the two testes per male.
9 There were a total of 50 sterile F, males. The two sterile males with normal testes weights
$ There were a total of 98 partially sterile. Testes weights for four of the animals were not were not included. Four of the animals clearly had small testes but weights were not taken.
taken.
Testes weights of the partially sterile animals were, on the average, similar to those of normal males. However, it appears that there is more variation in testes weights among those partially sterile than in normals, as indicated by a signifi- cantly higher standard deviation (P
<
. O l ) . Although there was no overlap in testes weights between the sterile males with small testes and the partially sterile ones, the weight of the testes of one of the latter was only 0.14 gram which is considerably smaller than normal.For 96 out of 98 partially sterile males, translocation heterozygosity was con- firmed by cytogenetic examination of diakinesis and metaphase I spermatocytes. Furthermore, partial sterility of all 96
F,
males was found to be transmissable to the following generation. The other two confirmed partially sterile mice, one each from 200 and 150 mg/kg doses, died before they could be progeny-tested.TABLE 5
Auerage fertilify of partially sterile mules bused on fGur litters
Number of partially Litter size Percent Litter no. sterile males (a%)* of normal
1 98 4c.2 6 . 3
2 98 4.9 43.6
3 98 4 7 40.9
4s C6 4.8 42.0
Average 43.2
* For comparison with litter size of normal males refer to Table 7.
TABLE 6
Auerage fertility of partially sterile males bmed on six females killed
Number Implants Living embryos Dead implants Class of males ( a v d ( a w l (Percent)
Partially sterile 98 9.3* 4.5 57.8
Normal 39 10.7 10.2 4.8
748 w. M. GENEROSO et al.
Two sets of fertility data (Tables 5 and 6) are available for the partially sterile
F, progeny. The first set was obtained by caging each male with a young female
of the (SEC X C57BL)F, strain to produce at least four litters. The second set was obtained by mating each one to at least six virgin (C3H X C57BL)Fl females that were killed during pregnancy for uterine analysis. On the basis of live births, the average fertility of partially sterile translocation heterozygotes was 43.2% that of normal males. This is comparable to the results obtained on sacrificed females. The average n u z b e r of living embryos for the partially sterile group is 44.1
%
that of normal controls. Data on sacrificed females (Table 6) show that fertilization involving sperm carrying unbalanced chromosome constitutions leads to death of embryos shortly be€ore or shortly after implantation i n most cases. This is indicated by the observation that reduction in the number of living embryos is attributable mainly to a n accompanying increase in dead implan- tation. However, a small proportion of embryonic loss occurred in early cleavage stages, as indicated by a significantly lower number of implantations in the partially sterile group.DISCUSSION
The dominant-lethal and translocation dose-effect curves clearly indicate that the process involved from the time the chemical is administered until the genetic damage is transmitted through the germ cells is a complex one. The dominant- lethal dose-effect curve departs markedly from linearity while the translocation curve significantly differs from the one expected on the basis of a two-hit kinetic.
It is obvious for both endpoints that the effectiveness of
EMS
is proportionately much lower at low doses and that there is a point in EMS dosage beyond which additional chemical results in a very sharp increase in mutation rates. That dose appears to be between 100 and 150 mg/kg. This observation suggests that there might be a mechanism operating at low doses of EMS that reduces either the production of chromosonial damage or transmission of the damage. Such a mechanism may manifest itself before the chemical reaches the target cell, or within the target cell before damage to the chromosomes, or after damage to the chromosome has been done. At the moment there is no substantial information available to help explain the shapes of the dose-effect curves. It is clear that genetic damage can be induced at doses as low as 50 mg/kg, which was the lowest dose used in this experiment. The threshold for genetic effects ofEMS,
if there is any, must be at a lower dose.One important aspect of this dose-response study from the practical standpoint is the comparison in efficiency between dominant-lethal mutations and heritable translocations as meascres of induced chromosome aberrations in male mice. T h e dominant-lethal test carried out in male mice is generally regarded as a very impel-tant mammalian test in the evaluation of mutagenic hazards of chemicals
EMS DOSE EFFECTS O N ABERRATIONS
749
and
RAY
et al. (1972) who were unable to detect dominant-lethal effects until the150 mg/kg dose was used. It is likely that dominant-lethal mutations had also been induced at the 100 nig/kg o r lower doses but were not detected owing to the relative insensitivity of the dominant-lethal procedure. It should be pointed out that the germ-cell stages sampled in this study are most sensitive to dominant- lethal induction but may not necessarily be so to the translocation induction. In addition to the higher sensitivity of the translocation procedure, translocations are a much more reliable endpoint in terms of human hazards than dominant- lethal mutations because, unlike the latter, they represent transmissible genetic damage. Thus, for the detection of low levels of chromosome breakage, transloca- tions are a more reliable endpoint than dominant-lethal mutations.
There is no doubt that EMS-induced partial sterility among F, male progeny, as determined by the procedure described here, is associated with induced recipro- cal translocations. Cytogenetic analysis of partially sterile animals, the trans- mission o€ partial sterility, and the study of CATTANACH, POLLARD and ISAACSON
(1968) o n EMS-induced partial sterility support this. Similarly, partial sterility in F, male progeny produced by radiation and other chemicals to male parents has almost always been associated with induced reciprocal translocations (see review by RUSSELL 1962). These observations clearly establish that partial sterility alone can be taken as an unequivocal indicator of a reciprocal transloca- tion. But what about induced complete sterility in F, male progeny?
It may be recalled that of the combined completely and partially sterile
F,
male progeny in the EMS experiment about one-third were of the completely sterile kind. I n a much lower number of F, males tested, CATTANACH, POLLARD
and ISAACSON (1968) obtained about equal numbers of sterile and partially sterile animals. Thus there can be little doubt that complete sterility among F, male progeny comprises a significant proportion of EMS-induced effects and it is important to know the basis for the induced sterility. There are now strong reasons to believe that at least the great majority of induced sterility among F,
males, like partial sterility, is attributable to induced chromosomal rearrange- ment. Some of the evidence had already been discussed by CATTANACH, POLLARD
and ISAACSON in 1968. In addition, through cytogenetic analysis of gonia1 and meiotic cells, they established the presence of translocations in seven out of 10
sterile F, males. The three that did not have observable abnormal karyotypes did not have meiotic cells. Since mitotic cells were analyzed, it is possible that rearrangements may have been present in these three cases but may not have resulted in obvious changes in chromosome size. Additional data on the cytologi- cal causes of F, male sterility after mutagenic treatment have recently been obtained by CAcHhmo, RUSSELL and SWARTOUT (1974) who found that the great majority of sterile sons produced by fathers post-meiotically treated with EMS
or X-rays were carriers of a translocation and that a majority of the transloca- tions were of the autosome-autosome type.
Undoubtedly, the most sensitive method for studying the ability of low doses
750 w. M. GENEROSO et al.
situation; (these results will be published elsewhere). Both lines of evidence indicate that translocations are a much more sensitive endpoint f o r measurement of low levels of chemically induced chromosome breakage than are dominant- lethal mutations.
We have developed a new procedure, a modification of the one used i n this study, for large-scale screening of translocations. The most common procedure for detecting translocation heterozygotes is to mate
F,
male progeny to three or more different females. Each female is opened at midpregnancy and living and dead implantations are counted. From the fertility data on three or more females, detection of complete sterility is straightforward. A partially sterile male is expected to sire, on the average, less than half as many living embryos as a normal male, with accompanying increases in the number of dead implantations. When only low numbers of F, males are to be tested, this procedure is the most suitable one. But when large numbers ofF,
males are involved, this is obviously a n expensive method and requires a good deal of animal handling and record keeping. The procedure described below reduces these problems, hence it is more suitable f o r large-scale screening and experimentation.The key to this screening procedure is the use of a strain 01 females with exceptional fertility such as the (SEC X C57BL)F,. The normal fertility of the (SEC x C57BL) F, females has previously been described ( GENEROSO, STOUT and
HUFF
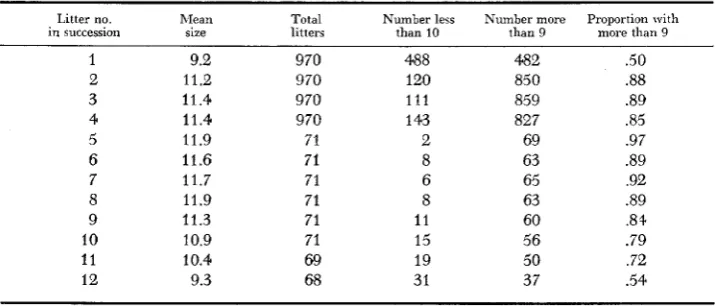
1971). Additional information is given in Table 7. Each F, male to be tested is caged with a female of the (SEC x C57BL)F1 strain. The females must be 10 to 12 weeks old and can continue to be used through the time the 12th litter is produced. If the size of the first litter is large enough, the male is declared fertile and discarded immediately after the litter is scored. If the first litter is not large enough, a second litier is produced. If the second litter size is large enough the male is declared fertile and immediately discarded. (When neither the first nor the second litter is large enough, the male is a suspect and is tested further by mating him to 3 virgin females, of any suitable strain, which are killed during pregnancy.) In any case, another male is placed with the female one week afterTABLE 7
Reproductive performance of (SEC X C57BL)F, females mated with normal mules
Litter no. hlean Total Number less Number more Proportion with in succession size litters than 10 than 9 more than 9
1 9.2 970 488 482 .50
2 11.2 970 120 850 .88
3 11.4 970 111 859 .89
4 11.4 970 1 43 827 .a5
5 11.9 71 2 69 .97
6 11.6 71 8 63 .89
7 11.7 71 6 65 .92
8 11.9 71 8 63 .a9
9 11.3 71 11 60 .8+
10 10.9 71 15 56 .79
11 10.4 69 19 50 .72
EMS DOSE E F F E C T S O N A B E R R A T I O N S 75 1
the litter is born and the same procedure is followed until the last male is added no later than after the 10th litter. The lapse of one week is required so that parentage of litters will not be confused. Since the female is normally mated shortly after parturition, a maximum of three litters are scored per male-two litters if a male is declared fertile after scoring his first litter and three litters if the decision is made after scoring the second litter. In all cases checking for newly born mice is made only when they are expected, i.e., pens are examined daily during weekdays beginning 18 days after pairing or appearance of a litter. Young are discarded immediately after they are scored.
From the EMS translocation experiment, it was found that out of 462 litters sired by 98 mice confirmed as partially sterile, only nine litters sired by inde- pendent males consisted of 10 o r more young, and only four of the nine litters had 11 o r more. The probability that a partially sterile male will sire a litter of 10 or more is then estimated as 0.0195 and of 11 or more is 0.0087.
It
is possible that these probabilities may not hold true for translocations induced by other agents but, if a litter of 10 o r more is used as a n indicator of full fertility, chances are that those that will be missed will be difficult to detect by any other means. Analysis of the fertility data of the (SEC X C57BL)F1 females mated with fully fertile males revealed that, if a litter size of 10 or more sired by a male inhis first or second litter is used as the indicator of full fertility and each female is allowed to produce 12 consecutive litters, a n average of 5.27 males from a sample clear of translocations may be tested per female. This average was calcu- lated by considering all possible configurations of two and three litters required far males for each female. Since each configuration is not equally likely, the average is a weighted average. For each configuration the number of males that can be tested is weighted by the chance that it will occur. There are 21 combi- nations of two and three litter periods that can test 4 , 5 or 6 males. For example, 6 males can be tested i n 6 periods of two litters,
5
males can be tested in one period of three litters followed by4
periods of two litters, and4
males can be tested i n4
periods of three litters. The chances of the three configurations are 0.293, 0.264 and 0.002, respectively. The chances are computed by using the last column in Table 7, which gives the probability that a fertile male will be declared fertile and thus require only two litters. The three configurations above can be combined with the other 18 and grouped according to the number of males that can be tested. The estimates of the proportions for 4, 5 and 6 males that can be tested are 0.0225, 0.6845, and 0.2930, respectively. The weighted average of 4, 5 and 6. by use of the above proportions, is 5.27. Out of these, the number of test males per female that can be declared fully fertile is estimated as 4.20. The remainder are classified as suspects and have to be tested further by mating them to at least three virgin females which will be killed during pregnancy. Obviously, the higher the frequency of translocations the lower the number of males that can be tested per female.Thus a large percentage of the F, males can be declared fertile by the simple procedure of counting the number of live births in at most two litters, with only small risk of declaring a partially sterile male as fertile. Since (SEC x C57BL) F,
752 w. M. GENEROSO et al.
males per female instead of sacrificing three or more females per F, male as in the old procedure. The procedure described above is now routinely used in our laboratory for screening male translocation heterozygotes produced in chemical and radiation experiments. A modification of this procedure can be made depend- ing upon the reproductive nature of the strain of females used in the screening and on the efficiency desired.
script.
The authors are grateful to DRS. RHODA F. GRELL and R. J. PRESTON for reviewing the manu-
LITERATURE CITED
CACHEIRO, N. L. A., L. B. RUSSELL and M. S. SWARTOUT, 1974 Translocations, the predominant
CATTANACH, B. M., C. E. POLLARD and J. H. ISAACSON, 1968 Ethyl methanesulfonate-induced
EHLING, U. H., R. B. CUMMING and H. V. MALLING, 1968 Induction of dominant lethal mu-
GENEROSO, W. M., SANDRA K. STOUT and SANDRA W. HUFF, 1971 Effects of alkylating chemi-
GENEROSO, W. M. and W. L. RUSSELL, 1969 Strain and sex variations in the sensitivity of mice
OAKBERG, E. F., 1960 Irradiation damage to animals and its effect on their reproductive ca-
X-ray sensitivity of primary spermatocytes of the cause of total sterility in sons of mice treated with mutagens. Genetics 76: 73-91.
chromosome breakage in the mouse. Mutation Res. 6: 297-307.
tations by alkylating agents in male mice. Mutation Res. 5 : 41 7-428.
cals on reproductive capacity of adult female mice. Mutation Res. 13: 171-184.
to dominant-lethal induction with ethyl methanesulfonate. Mutation Res. 8 : 589-598.
pacity. J. Dairy Sci. (Suppl.) 43: 54-67.
mouse. Intern. J. Radiation Biol. 2: 196209.
Comparative studies of induced mutations with host-mediated, dominant-lethal and cyto- genetic assays. Program and Abstracts, 3rd Annual Meeting, Environmental Mutagen So-
ciety, March 26-29, 1972, p. 14.
Chromosome aberrations in experimental mammals. pp. 230-294. In:
Progress in Medical Genetics. Vol. 2. Edited by A. G. STEINBERC and A. G. BEARN. Grune and Stratton, New York.
Corresponding editor: E. H. Y. CHU OAKBERG, E. F. and R. L. DIMINNO, 1960
RAY, V. A., H. E. HOLDEN, D. S. SALSBURG, J. H. ELLIS, L. J. JUST and M. H. VOYER, 1972