Accepted for publication August 19, 2001
ABSTRACT
The maizerlocus encodes a transcription factor that regulates the developmental expression of the plant pigment anthocyanin. In an unusual example of gene regulatory diversity, theR-sc (Sc,strong seed color) and theR-p(P,plant color) alleles ofrhave nonoverlapping tissue specificity and nonhomologous 5⬘flanking sequences. Heterozygotes between wild-typePandScmutants withDs6transposable element inserts (r-sc:m::Ds6orsc:m) produce colored seed derivatives (Sc⫹) during meiotic recombination. The sc:malleles withDs6 insertion in 3⬘regions ofrproduce crossoverSc⫹derivatives. sc:malleles withDs6 elements inserted in 5⬘regions produce rareSc⫹derivatives borne on nonrecombinant chromosomes. Among 52 such noncrossoverSc⫹derivatives, 18 are indistinguishable from theScprogenitor in phenotype and DNA sequence [Scp(⫹)alleles]. The remaining 34 derivatives have strongSc⫹expression, including darkly pigmented aleurone, scutellum, coleoptile, and scutellar node [Scp(e)alleles]. The coleoptile and scutellar node phenotypes are unique from either progenitor but are similar to those of some naturally occurringralleles. Both classes ofSc⫹derivatives are explained by gene conversion between the promoter region of Sc:124 and a homologous region located proximal to P. The recombinational intermediate formed betweensc:malleles andPresults in deletion of theDs6element alone or bothDs6and a nearby unrelated transposable element-like sequence.
O
NE of several loci in maize that regulates the tran- r haplotype. Individual genes in a haplotype can be arranged in either direct or inverted orientation and scription of the genes that encode thebiosyn-thetic enzymes for anthocyanin, the purple pigment either closely spaced or spread over several hundred kilobase pairs (Eggleston et al. 1995; Walker et al.
found in plants (reviewed inDooner et al. 1991), is r
(red-1). r, and the unlinked genes b (booster-1) and in 1995). This complexity is partially responsible for gener-ating the diverse expression patterns ofralleles during (intensifier-1), encode helix-loop-helix proteins of the
mycfamily of transcription factors (Ludwiget al.1989; maize evolution; individualrgenes within a given haplo-type are often associated with different tissue-specific
Radicellaet al.1991;Goffet al.1992;Burret al.1996).
patterns of expression. Genes in complexrhaplotypes These genes determine the developmental timing and
also participate in the epigenetic phenomena of para-tissue specificity of anthocyanin deposition in maize
(re-mutation (Brink 1958; reviewed in Chandler et al.
viewed in Ludwig and Wessler 1990). Naturally
oc-2000) and imprinting (Kermicle 1970a; reviewed in curringralleles are unique among plant genes because
Kermicle1996;Alleman andDoctor2000). Both of of the diverse array of tissue-specific expression patterns
these phenomena are related to structural complexity that pigment nearly all parts of the plant and kernel.
at r (Kermicle et al. 1995; reviewed in Alleman and For example,R-Nj:Cudu(Navajo) controls pigmentation
Doctor 2000). Simplex (single transcriptional unit) r
in the crown of the kernel, the root, coleoptile, scutellar
alleles can be isolated by out-of-register pairing and node, silk, and anthers, whereasLc(leaf color) pigments
recombination between genes in complex haplotypes the leaf blade, ligule, auricle, glume, lemma, palea,
peri-(McWhirterandBrink1962;DoonerandKermicle
carp, and midrib of the adult plant (Styleset al.1973;
1971;Robbinset al.1991;Egglestonet al.1995). Such
Ludwiget al.1990).
r alleles are well suited for studies of gene regulation Therchromosomal region in many maize varieties is
and recombination. Simplex r alleles have been used structurally complex, containing up to five genes per
to define many aspects of r gene function, including identification of nuclear localization signals (Liu et al.
1998), gene regulation by upstream open reading
Corresponding author:Mary Alleman, Department of Biological
Sci-frames (WangandWessler1998), andcis-acting con-ences, Duquesne University, Pittsburgh, PA 15282.
E-mail: [email protected] trol sequences (Liuet al.1996).
rand other kernel-specific maize genes are also used the property of AcandDsto reinsert at positions near the donor site during transposition (Doonerand
Bela-in the study of homologous recombBela-ination Bela-in higher
plants. Recombinant alleles can be recovered from het- chew1989;Morenoet al.1992). We have determined, in this study and previously, that each of thesc:mmutants eroallelic combinations by selecting phenotypic
deriva-tives from large populations of progeny. Several novel contains the sameDs6transposable element. Insertion sites for the mutants are distributed from 2.5 kbp 5⬘of observations have resulted from studies of
recombina-tion in plants compared to the yeastSaccharomyces cerevis- the start of transcription to the end of the protein-coding region (Alleman and Kermicle 1993). Each
iae. First, studies with the genesa(anthocyaninless-1),r,
wx, andbz(bronze-1) indicate that, although conversion allele is germinally and somatically stable in the absence of Ac but is unstable (spotted) in the presence ofAc, tracts are the same approximate length in plants vs.
yeast, noncrossover products are recovered much less typical of Ds-containing alleles (McClintock 1951;
Kermicleet al.1989). frequently than crossover products (Doonerand
Ker-micle1986;DoonerandRalston1990;Xuet al.1995; We have used heteroallelic combinations ofPand 39 of the 41 sc:mto recover colored seed (Sc⫹) progeny.
DoonerandMartinez-Ferez1997;OkagakiandWeil
1997). This result is in contrast with similar studies in This article reports a striking difference in the recombi-national properties of sc:m that are located in 3⬘ as yeast in which equivalent numbers of crossover and
non-crossover products are usually recovered for alleles at compared to 5⬘regions ofSc:124. Frequently, 3⬘-located mutants recombine with Pto produceSc⫹ derivatives, homologous locations (reviewed in Peteset al. 1991).
Another interesting observation regarding recombina- usually crossover in origin. In contrast, 5⬘-located mu-tants produce Sc derivatives that are reduced in total tion in maize genes is an absence of recombinational
polarity for crossover resolution (DoonerandMarti- frequency and noncrossover in origin. In addition, some of theSc⫹derivatives from 5⬘-locatedsc:mhave a novel
nez-Ferez1997;OkagakiandWeil1997). The strong
polarity of conversion tracts among the many yeast genes phenotype in which aleurone, scutellum, coleoptile, and scutellar node are strongly pigmented, distinct from studied is interpreted as resulting from a limited
num-ber of sites at which double-stranded breaks (DSBs) can sc:m,P, orSc:124. These derivatives are associated with loss of the Ds6element and another nearby sequence initiate recombination (Peteset al.1991). Thus, a lack
of polarity might indicate more frequent or nonspecific fromSc:124. Structural attributes of thergene in which the two groups of sc:m alleles reside are not sufficient sites for DSBs (Schnableet al.1998).
This study explores recombination between tworal- to explain this recombinational dichotomy. leles chosen for their phenotypic dissimilarity.R-sc(Sc)
andR-p (P) alleles are derived as simplex alleles from
MATERIALS AND METHODS
the R-st (stippled) andR-r:std (standard) complexes, re-spectively (McWhirterandBrink1962;Doonerand
Maize lines and genetics:All maize lines used had an inbred Kermicle1971;Robbins et al.1991;Egglestonet al. W22 genetic background, established by five or more
genera-1995). Scalleles specify strong anthocyanin pigmenta- tions of backcrossing. TheralleleR-sc:124(Sc:124) is a simplex allele that is the progenitor of allsc:mmutants described in
tion in the aleurone and scutellum of the maize kernel
this report. The phenotype of Sc:124 and other similar Sc
and weak pigmentation of the coleoptile of the seedling.
alleles is strong nonimprinted aleurone pigmentation and
P alleles pigment the root and coleoptile of the
seed-weak coleoptile pigmentation. Thesc:m alleles used in this
ling and the anthers of the mature plant, but produce research are colorless or pale seed mutations described pre-colorless aleurone. The phenotypes of ScandPalleles viously (Kermicle et al. 1989). The simplex R-p (P) alleles are shown in Figure 1, A and B. Recombinant products used in this research (P:n19,P:n46, andP:n142) are equivalent in structure (Robbinset al.1991).Palleles pigment coleoptile,
are readily recovered from heteroallelic combinations
root, and anthers strongly but do not produce seed
pigmenta-of wild-typePand mutantScalleles. These alleles retain
tion. They are collectively designated as “P.” The phenotypes
the tissue specificities of the 5⬘flanking sequences from ofScandPalleles are shown in Figure 1, A and B.
the original progenitor. These results indicate the equiv- Severalralleles were used in these experiments because alence of 3⬘ coding regions of P and Scand the loca- of contrasting phenotypes or structures readily distinguished from the test alleles. Ther-⌬902allele is a deletion including
tion of regulatory sequences in the 5⬘nontranscribed
11.3 kbp ofrcoding region and 5⬘ untranslated sequences
regions (Kermicle1985, 1988).
(Alleman and Kermicle 1993). The r-g:8p allele is a pale
The target alleles for this study include
indistinguish-seed colorless plant derivative of R-r:std. R-Navajo:Cudu(Nj)
able P alleles and 41 transposable element-containing is a simplex r allele that pigments the crown of the maize mutants of the simplex Sc:124 allele. This series of kernel, the root, coleoptile, scutellar node, silk, and anthers (Styleset al.1973;Dellaportaet al.1988). TheR-st(stippled)
r-sc:m::Ds6mutants (or, for simplicity,sc:m) were
gener-haplotype is composed of four rgenes [(Sc), (Nc1), (Nc2),
ated using a cyclic mutagenesis scheme in which the
and (Nc3)] (Egglestonet al. 1995). TheR-r:std (standard)
transposable elementDs6was mobilized by the
autono-haplotype is composed of threergenes [(P), (S1), and (S2)]
mous element, Ac, to excise and reinsert back into Sc and one r-promoter pseudogene (q) (Robbins et al. 1991;
at a high frequency (Kermicle et al. 1989; Alleman Walkeret al.1995).
Ther-linked locig1(golden-1 plant color, 20 cM proximal)
Figure1.—The phenotypes of progeni-tors andSc⫹ derivatives from sc:m/P het-eroallelic combinations. (A) TheScallele, Sc:124; (B) the P allele, P:n46; (C) the Scp(⫹)type allele,R-sc:n5993; and (D) the Scp(e)type allele,R-sc:n6008. B includes ab-breviations referring to parts of a maize seedling: AL, aleurone; C, coleoptile; SC, scutellum; SN, scutellar node; R, root. The aleurone is part of the triploid maize endo-sperm whereas the coleoptile, scutellum, scutellar node, and root are derived from the developing embryo. Maize seeds were germinated under fluorescent lights at room temperature and then incubated at 18⬚–20⬚(12 hr light/12 hr dark) for 5 days. All seedlings shown are homozygous for the specifiedralleles.
andmst1 (modifier of stippling due toR-st, 6 cM distal) were sc:mmutant alleles used in these experiments include 42 insertions of the transposable elementDs6(sc:m3,m301-m328, used as flanking markers for recombination studies. Theg1
alleles that were used areG1-w22(wild type) andg1-1(mu- m330-m341, andm9) and one with insertion of transposable elementDs1(sc:m1). These alleles produce colorless or pale tant). Theg1phenotype is scored during a germination test
in which g1-1 produces a light golden-colored plant tissue aleurone in the absence of transposable elementAcand spot-ted aleurone in the presence ofAc(Kermicle1980; Kermicle relative toG1-w22, which produces green tissue. TheMst1-1
mutant allele is a dominant modifier of stippling that is assayed et al. 1989). The Scp(⫹) type alleles produce a phenotype that is indistinguishable fromSc:124, with darkly pigmented in the presence of the r-g:nc3.5 allele (Kermicle 1970b).
r-g:nc3.5produces aleurone that is stippled with sectors of aleurone and scutellum and weakly pigmented coleoptile. Al-leles that were used for molecular analysis include:Sc:n5993 anthocyanin pigmentation in the presence of Mst1-1. The
r-g:nc3.5allele produces nearly colorless aleurone in the pres- and Sc:n5991 (derived from m305), Sc:n7097 and Sc:n7098 (derived from m316), and Sc:n6732 and Sc:n7052 (derived ence of mst1-w22. The w22 designation of alleles refers to
those present in the W22 inbred background. fromm330). TheScp(e)type alleles produce darkly pigmented aleurone, scutellum, coleoptile, and scutellar node. Alleles TheP-vv::Acallele (variegated pericarp color) of thePlocus
on chromosome1was used as the source of the autonomous that were used for molecular analysis include Sc:n5992 (de-rived fromm305),Sc:n6008andSc:n6009(derived fromm311), transposable element Ac (Activator) for these experiments.
The unlinked recessive allelewx1-1(waxy-1seed) was used as Sc:n7095(derived from m316), and Sc:n7053 (derived from m330).
Isolation ofSc⫹derivatives:The genetic starting material and MAR103 (TTCGTTTTTTACCTCGGGTTC). MAR102 and MAR103 are specific to transposable elementAcandDs6 for these experiments includes 39 sc:m mutations ofSc:124
with insertion of theDs6element. Positions of the insertions sequences. Other primers were from theSc:124sequence. The locations ofScandPprimers are shown in Figure 2. are distributed throughout thercoding and 5⬘flanking
re-gions (AllemanandKermicle1993). The positions of theDs6insertions at the 5⬘end of theSc gene were determined usingDs6primer MAR102 or MAR103 Recombination tests were performed as follows. Seeds of
the genotype g1-1, P, Mst1-1/G1-w22, sc:m::Ds6, mst1-w22; and a flankingr-specific primer. PCR was performed using genomic DNA as a template and Amplitaq DNA polymerase wx1-1/Wx1-w22 and involving each of the sc:m alleles were
planted as detasseled female rows in an isolation plot with (Applied Biosystems, Foster City, CA). PCR products were purified and cloned into the pCRII vector (Invitrogen, San alternating rows of homozygousg1-1,r-g:8p mst1-w22;wx1-1as
the male parent. The resulting ears were screened for Sc⫹ Diego) for sequencing.
Probes were gel-isolated restriction fragments or PCR prod-kernels. The selected kernels were progeny tested to
deter-mineSc⫹heritability and the flanking marker combination. ucts that were radiolabeled by random-primed incorporation of [32P]dCTP or digoxygenin-labeled dCTP (Roche Molecular
Seedling phenotypes ofPandg1were assayed in germination
trays under fluorescent lighting at 18⬚–20⬚. Themst1pheno- Biochemicals, Indianapolis). PCR using primer combination F4/B971 andP DNA as a template was used to isolate the type was tested by crossing Sc⫹ individuals with the nearly
colorlessR-stderivative alleler-g:nc3.5and pollinating the het- YL1 probe. Probes BH1415 and PX1029 were gel-purified restriction fragments from a subclone of MF2. Nj:1 was a erozygous plants with a colorless seedrallele. The
recombi-nantMst1-1,r-g:nc3.5phenotype is spotted kernels while the probe derived from clone pR-Nj:1, a BglII/HincII subclone from theR-Nj:Cudumutantr-nj:m1(Dellaportaet al.1988). mst1-w22,r-g:nc3.5phenotype is unspotted kernels.
In most experiments, the genotype g1-1 sc:m mst1-w22/ Probe YL202 was a PCR product generated using primers F3 and B66 and Sc:n5992 genomic DNA as a template. Probe G1-w22 P (P:n19, P:n46, or P:n142) Mst1-1 was the female
parent. In a few experiments involving 5⬘-locatedsc:m, seeds YL210.1 was a PCR product generated using primers F3 and B530 andSc:n6008genomic DNA as a template. Locations of of the genotypeg1-1 sc:m mst1-w22/G1-w22 P(p:m3orp:m1)
Mst1-1in which aDsinsertion was also present at thePallele Scregions used as probes are shown in Figure 2.
The location ofP:Awas determined with respect to theP located in the protein-coding region were used (Doonerand
Kermicle 1986; Alleman and Kermicle 1993). Data from gene using Southern blot analysis of restriction fragment length polymorphism (RFLP) data. Two types of recombinant this population are included in Table 2.
alleles were used: (1) threeSc⫹alleles fromsc:m/P heterozy-Molecular analyses:Maize DNA was isolated according to
gotes in which flanking marker combinations indicated a re-Chen et al. (1992) except that DNA extractions were
per-combinant origin (Sc:n3081,Sc:n3082, andSc:n3083derived formed at room temperature. DNA was digested, size
fraction-fromsc:m308/Pheterozygotes) and (2) threep:m::Dsalleles ated, and blotted as described in Alleman and Kermicle
(p:m1,p:m3, orp:m9) that had the reciprocal combination of (1993). For cloning, DNA fromR-stwas digested withSau3AI
flanking markers relative to theSc⫹alleles (Kermicle1988). (1.74 units/100g/ml) for 15, 30, 45, and 60 min; pooled;
and partitioned on a 10–40% glycerol gradient. DNA frag-ments (8–15 kbp) were ligated with EMBL3 arms using T4 DNA ligase. A total of 1.2⫻106recombinant phage were
RESULTS screened using radiolabeled r probe (Nj:1). Three clones
(MB1,MD1, andMF2) contained the 5⬘region of theSc Recombination with P divides theSc gene into two gene, confirmed by restriction endonuclease mapping. Clone distinct regions:The general approach of these experi-R4.7 containing thePgene ofR-r:stdwas received from Dr.
ments was to use meiotic recombination to explore the
T. Robbins (Robbinset al.1991).
structure and behavior of two distinct rprogenitor
al-Regions of clonesMF2 and R4.7 were subcloned into
pBluescript vectors (Stratagene, La Jolla, CA) for DNA se- leles,Sc(seed color)andP(plant color). These tworalleles
quence analysis. Dideoxy chain termination sequencing using were derived from very different progenitorrhaplotypes Sequenase II kits (United States Biochemical, Cleveland) was and have nonoverlapping tissue specificity. By con-used with single-stranded templates. Sequencing strategies
in-structing heterozygotes of 39 colorless seedsc:mmutants
cluded subcloning, synthetic primer design, and the
genera-and wild-typePalleles, colored seed derivatives can be
tion of nested deletions (Exo/Mung deletion kit, Stratagene).
Subsequent sequencing of mutants and derivatives used dou- selected. The diagram in Figure 3 shows the
composi-ble-stranded sequencing of plasmids or PCR products and tion of the heterozygotes that were used in these experi-the ABI automated DNA sequencing system (Perkin-Elmer, ments and the strategy for derivative selection.
Norwalk, CT). All DNA sequence analysis used MacVector
From populations of 13,700–46,500 progeny kernels,
software (Oxford Molecular Group, Palo Alto, CA).
Sc⫹derivatives were recovered in frequencies ranging
Synthetic oligonucleotides (Oligos or Operon, Alameda,
CA) specific to regions of theDs6element or thergene were from 0 to 0.233% Sc⫹ kernels per sc:m chromosome
used as PCR primers to generate probes, for inverse PCR, and tested (Table 1). The 39sc:malleles fall into two distinct for the mapping of the position ofDs6elements inSc. They groups defined by the relative frequency ofSc⫹ deriva-include the following: B32 (CTCGATGTCTGTCAAGCGT
tives and by the association of these derivatives with
TGG), B66 (TTCACCAAACAAAAGCTGGCGG), B971 (GCT
recombination of alleles of the flanking marker loci
AGCCTTGCTAATAC), B104 (TTGGCTGTACGCATTACCA
TGC), B129 (TTGTGTCATGTGGCTCC), B530 (GTACGT golden-1 (g1) and modifier of stippled-1 (mst1). Group I
AACACGCATGG), B2261 (TAGGAGTTTGGGACTCTTCCG), sc:mmutants produceSc⫹derivatives at relatively high F3 (TGCGACGGACAAGAAGATTTG), F4 (TTCTCGGACGT frequencies, averaging 6.1⫻10⫺4Sc⫹per chromosome
TTTCCCAGCAG), F17 (AAGCAGCAGGAGACGAAGGTAG),
tested. Nearly all of these derivatives (416 of 434Sc⫹)
F59 (GACGGACGAATTGAAGAACACCTG), F66 (CCGCCA
are carried on recombinant chromosomes carrying the
GCTTTTGTTTGGTGAA), F1178 (ATCCTCAGACGCTACA
Figure2.—The positions ofDs6insertions in the 5⬘region ofSc. Differentially shaded regions represent sequence landmarks. The positions of the origin of transcription and translation (atg) are based on those determined for theLcgene (Ludwiget al. 1989). “Rcommon sequences” refer to regions present in bothScandP, including 698 bp of noncoding DNA (shaded section) and allr-coding sequence (labeled in the figure). Vertical lines indicate positions ofDs6insertions in ther-sc:mmutations. The Sc:Aregion is defined as Sc-specific sequences contained withinclone MF2 and located 5⬘of allrcommon sequences. The arrows represent the positions of PCR primers discussed in the text. Probes Nj:1, YL202, YL210.1, BH1415, and PX1029 are designated by shaded bars. Probe YL202 was synthesized by PCR using an allele (R-sc:n5992) with a deletion of theBnotsegment.
marker of theP parent. In contrast, heterozygotes in- kernels were absent in populations ofsc:mhomozygotes (Kermicle et al. 1989). It is unlikely, therefore, that volving group IIsc:mmutants produceSc⫹infrequently,
averaging 6.4⫻10⫺5Sc⫹per chromosome tested, most the noncrossoverSc⫹ class marked as parentalsc:m is attributable toAc-mediated transposition.
of which are associated with the noncrossover flanking
marker combination of thesc:mparent (21 of 28Sc⫹). Group II sc:mmutants are located in 5ⴕflanking re-gions ofSc:Preliminary studies indicated that group I Spotted kernels were not present among the testcross
populations, indicating absence of an activeAcelement. alleles contain Ds6 insertions located in relatively 3⬘ regions ofrwhereas group II alleles are located in rela-Furthermore, germinalSc⫹revertants as well as spotted
tively 5⬘ regions, mostly upstream of the transcription start site (AllemanandKermicle1993). To determine the precise location of the insertions, the 5⬘ flanking region of the Scgene and the insertion site of the mu-tants were sequenced, and 5⬘ portions of the Scgene from R-st were cloned into EMBL3 as recombinant phage MF2, part of which was subcloned and se-quenced (GenBank accession number forSc5⬘flanking DNA: AF380388). TheScsequence was compared with the published sequence of thelcgene, a displaced mem-Figure3.—Structure ofsc:m/Pheterozygotes used as pro- ber of thergene family, to determine sequence
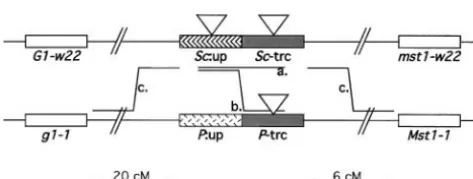
land-genitors forSc⫹derivative selection. Shown are therchromo- marks and the orientation of the cloned DNA (Ludwig somal regions containing ansc:mallele (top) and aPallele
et al.1989).
(bottom) that were used in these experiments. Outside
flank-The precise locations of the 5⬘-located group II alleles
ing markers includeg1(golden plant parts) andmst1(modifier
and alleles near the boundary of the two regions were
of stippling by R-st).ScandPare divided into transcribed (trc)
and 5⬘-flanking (up) sequences. The differential shading of determined by PCR analysis and DNA sequencing. PCR
Sc:upand P:updenotes the nonoverlapping tissue specificity products overlapping the Ds6 insertion sites for each produced by these alleles.Ds6 insertions insc:mare located
of the mutants were generated using oligonucleotide
in eitherSc:uporSc:trc(triangles). In some experiments,Ds6
primers, one specific to Ds6 and the other using Sc
insertions were located inP:trc. Recombination distances are
sequence near the mapped positions of thesc:malleles.
designated in centimorgans. Thesc:mphenotype is colorless
or pale-colored seeds and colorless plant parts. ThePallele Sequencing determined the insertion site for each
mu-phenotype is colorless seed and pigmented root, coleoptile, tant as the 8 bp of target site duplication adjacent to and anthers. Two types ofSc⫹derivatives are produced by
theDs6element. The insertion sites for the 19 5⬘-most
recombination betweensc:mand P, designated as “a”
(non-Ds6insertional mutants are shown on the map ofScin
crossover) and “b” (crossover). Double crossover (DCO)
TABLE 1
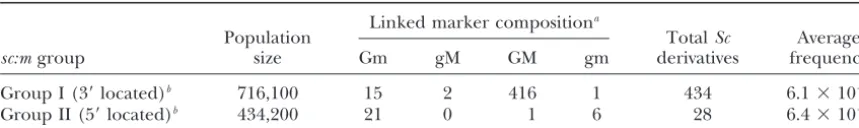
Sc⫹derivatives fromG1-w22 r-sc:m mst1-w22/g1-1 R-p Mst1-1heterozygotes
Linked marker compositiona
Population TotalSc Average
sc:mgroup size Gm gM GM gm derivatives frequency
Group I (3⬘located)b 716,100 15 2 416 1 434 6.1⫻10⫺4
Group II (5⬘located)b 434,200 21 0 1 6 28 6.4⫻10⫺5
Sc derivatives are isolated from G1-w22 sc:m::Ds6 mst1-w22/g1-1 P Mst1-1 heterozygotes.g1 is thegolden-1 gene, located 20 cM proximal toron chromosome10;mst1is themodifier of stippled-1gene located 6 cM distal toron chromosome10L. Chiasma interference in theg1-r-mst1region was estimated at 0.87 (Kermicle1970b). Group I alleles arem3,m301,m302,m303,m304,m306,m307,m308,m310,m312,m313,m314,m317,m318, m319,m320,m321,m325,m326,m327,m334,m335,m336,m339, andm340. Group II alleles arem305,m311, m315,m316,m322,m323,m328,m330,m331,m332,m333,m337,m338, andm341.
aFor the proximal marker, G is theG1-w22allele, g is theg1-1allele. For the distal marker, m is themst1-w22 allele, M is theMst1-1allele.
bGroup I mutants (3⬘locations)vs.group II mutants (5⬘locations) are defined by the change from predomi-nantly crossover to noncrossoverScderivatives.
II alleles are demarcated at an approximate position genome (not shown). This region (called P:A) was cloned using an inverse PCR (I-PCR) strategy (Ochman
370–470 bp 3⬘ of the Sc transcription start site or
be-tween the insertion sites ofsc:m337andm312. Although et al.1988). Genomic DNA ofR-r:std(P) and ther dele-tion mutantr-⌬902was digested separately with the re-derivative frequencies from sc:m/P for sc:m located
close to this position are infrequent,Ds6insertion sites striction enzymesBamHI,BglII, andXbaI. Digested DNA was diluted to 2.5 ng/l, favoring the formation of mo-close to the boundary includem338(four convertants/
zero crossovers), m337 (two convertants/zero cross- nomeric circles, and ligated with T4 DNA ligase to circu-larize the fragments. PCR using primers F4 and B530 overs), m312 (zero convertant/one crossover), m320
(zero convertants/zero crossovers), and m308 (zero amplified the flanking sequences. Southern blots of the PCR products were hybridized with YL210.1 to confirm convertants/three crossovers). Crossover data for
indi-vidual alleles are not delineated in Table 1. homology withSc:A. No PCR products or hybridization resulted when r-⌬902 DNA was used as the template The sequence of thePregion homologous to thesc:m
insertion sites indicates a structurally segmented region (not shown). Three PCR products fromPwere cloned, including pYLB, pYLG, and pYLX, denoting the restric-proximal to thePallele:Because upstream regions of
maize genes can be very different for alleles with distinct tion enzyme source of the digested DNA asBamHI (B),
BglII (G), or XbaI (X). Clone pYLB was sequenced as tissue specificities (Radicellaet al.1992), it was
impor-tant to compare the sequence of theScandP5⬘flanking described above.
The sequence of I-PCR clone pYLB indicates a highly regions to determine the location of theDs6insertion
relative to homologous regions of P. The P allele 5⬘ homologous, yet fragmented,P:Aregion relative toSc:A
(diagrammed in Figure 4). Four significant regions of flanking region was received as part of cloneR4.7 from
Dr. T. Robbins (Robbinset al.1991). Maize DNA from discontinuity exist between these two sequences: (1)
P:YL1is a 1.6-kbp region located inP:Abut not inSc:A;
R4.7 was subcloned and sequenced (published as
Gen-Bank accession no. AF380390). Sequence comparison (2)Bnotis a 471-bp region located inSc:Abut not inP:A
(published independently ofSc:Aas GenBank accession of a 5646-bp region of MF2 and a 4702-bp region of
R4.7 indicates thatScandPare 99.7% identical in the no. AF380387); (3)P:YL2is a large region of unknown length located between P:AandP, partially contained 698 bp 5⬘ of the start of transcription. Further 5⬘ of
⫺698, the two alleles are completely nonhomologous in clone R4.7 (P); (4) A 416-bp segment of Sc:A is located adjacent to the r common region but is not within the sequenced regions, including 3099 bp of 5⬘
flanking DNA fromSc(MF2) and 3560 bp of 5⬘flank- present in P or P:A (Figure 4). These four insertion-deletion differences betweenP:AandSc:A are defined ing DNA fromP(R.47). The segment fromMF2 that
is unique to theScgene contains the insertion site for as sequences that are present on one homolog but absent from the other. Excluding these insertion-deletion dif-13sc:mmutants and is designatedSc:A(Figure 2).
A recombination-based mechanism for the produc- ferences, the homologous segments shared bySc:Aand
P:Aare identical in sequence, a total of 2209 bp of DNA. tion ofSc⫹alleles from sc:m/Pheterozygotes requires
that sequences homologous to the insertion sites ofDs6 To characterize the genomic andr-linked copy num-bers of insertion-deletion differences betweenP:Aand mutations must be present inP. Southern blot analysis
Figure4.—rgenesScandPshare regions of discontinuous homology in the 5⬘region. Alignment of the 5⬘regions ofScand Prelative toDs6insertion sites insc:mmutants is based on sequence data for the 5⬘flanking DNA ofSc,P, and a homologous, proximally linked segment found in thePgenotype (P:A). The sequences of clonesMF2 (Sc), R4.7 (Pgene,Robbinset al. 1991), and YLB (P:A) were used in the generation of these maps. Sequence landmarks and regions of sequence nonhomology are diagrammed as shaded boxes and are consistent with shading in Figure 2. Homologous stretches of DNA in thePand Sc alleles are separated by blocks of nonhomologous DNA. Four insertion/deletions comparingSc:Aand P:Aare shown on the map: (1) theBnotinsertion in Sc:A; (2) theYL1region in P:A; (3)P:YL2, theP-specific 5⬘ flanking region adjacent to theP allele in cloneR4.7; and (4) a 416-bp segment located inSc:Abut missing fromPorP:A. All inserts are to scale except the P-specific regionP:YL2, of unknown length.
moter of allrgenes; BH1415, a 1.4-kbp segment ofSc:A multiplergenes or pseudogenes. Previous studies indi-cate thatR-r:stdandR-stcontain four Nj:1-homologous that is homologous to bothP:AandSc:A;YL1, a segment
unique to P:A; PX1029, a segment ofSc:A containing segments (Robbinset al.1991;Egglestonet al.1995;
Walkeret al.1995). These Southern blot data can be part of theBnotelement; and YL202, a region ofSc:A
that overlaps PX1029 but excludesBnot. Figures 2 and summarized as follows: Whereas therpromoter region probe Nj:1 hybridizes to allrgenes in complex alleles, 4 show the location of these probes. Results are
summa-rized below. the Sc:A probe BH1415 hybridizes to a single copy
se-quence inR-st,R-r:std, and all derivatives. A representa-First, the relative copy number of the
insertion-dele-tion differences was determined by Southern blot analy- tive Southern blot is shown in Figure 5C. These results indicate that the Sc:A/P:A region is a single-copy se-sis. The YL1 region, present in P:A but not Sc:A, was
used as a probe on Southern blots containing genomic quence, which is not duplicated in complex r alleles such as R-standR-r:std.
DNA fromSc:124andP:n46. A single band appears for
P DNA but not for Sc:124 DNA (Figure 5A). YL1 is, TheP:Aregion is located proximally toPon chromo-some 10 and is not duplicated in complex r alleles: therefore, unique to the genome of P and does not
exist in theScgene or genome. Similarly, probes were Locating P:A relative to theP transcriptional unit was also accomplished by Southern blot analysis using RFLP generated from Bnotand the region of Sc:A that
sur-roundsBnot. The probe PX1029 contains most of the analysis of recombinantralleles. Two types of recombi-nant alleles were used, including (1) Sc⫹fromsc:m/P
471-bpBnotsegment. Hybridization of PX1029 to
South-ern blots containing Sc:124 and P:n46 DNA indicates heterozygotes that were recombinant with respect to flanking markers and (2) the reciprocal product of this that PX1029 hybridizes toⵑ20 bands for DNA of either
genotype (Figure 5B). A larger probe of this region event,p:m::Ds (refer to Figure 3). Southern blots were produced using digested DNA from recombinant alleles excluding the Bnot segment (YL202) hybridizes to a
single band on the same Southern blots (not shown). and theSc:Aregion probe BH1415 to map the position of P:Arelative to the P structural gene. Recombinant Thus,Sc:Ais a single-copy segment and theBnotelement
is the cause of the repetitive hybridization of probe Sc⫹alleles (for example,Sc:n3081) contain the BH1415-hybridizing restriction fragment that is associated with PX1029.
To determine the copy number of Sc:A/P:A within theScallele. Recombinantp:m::Dsalleles (for example,
p:m1andp:m9) contain the BH1415-hybridizing restric-complexrhaplotypes, Southern blots containing
geno-mic digests ofSc:124,P:n46, complexr haplotypesR-st tion fragment that is associated with thePallele. Repre-sentative Southern blot data are shown in Figure 5D. and R-r:std Lc, were hybridized with probes Nj:1 and
In addition, individual restriction fragments from P, moter, do not hybridize to both BH1415 and Nj:1 (not shown), indicative of distance between thePpromoter even fragments ⬎15 kbp in length in which the
andP:A. Thus,P:Ais located at a quasi-ectopic proximal 3⬘-located restriction site is located within the P
pro-site compared to the location ofSc:A, which is immedi-ately 5⬘to the Scpromoter.
Bnotis a non-LTR-transposable element:Several fea-tures of theBnotsegment fromSc:Aare consistent with this element being a maize transposable element. First,
Bnot is homologous to a multicopy sequence in the maize genome (Figure 5B). In addition, the Bnot se-quence in Sc:A is flanked by a 22-bp imperfect direct repeat sequence [TCTTGCTTTTTTCTACTT (T/G) TTT] that is present once in theP:Asequence. This 22-bp sequence is assumed to represent the target site that was duplicated during insertion ofBnotinto theScgene ofR-stduring maize evolution.
The 449-bpBnotsequence, excluding the 22-bp flank-ing repeats, was compared to sequence databases usflank-ing the BLASTN program (Altschulet al.1997). A strongly homologous sequence of 447 bp (83% sequence iden-tity, BLAST expect value⫽3⫻10⫺75) was identified in the maize genome several kilobase pairs downstream of theAdh1-Fallele (Tikhonovet al.1999). Nine of 10 bp near the termini of this element are duplicated and may represent part or all of the direct repeat produced duringBnotinsertion nearadh1-F(not shown). In addi-tion, each of the six reading frames forBnotwas trans-lated (MacVector software; Oxford Molecular Group, Genetics Computer Group) and compared with protein database sequences using BLASTP. One open reading frame of 107 amino acids identified homology to four putative plant reverse transcriptase sequences, includ-ing three rice sequences and one Arabidopsis sequence
Figure5.—RFLP and Southern blot analysis of theSc:A/P:A regions ofr. Maize genomic DNA was digested with restriction enzymes and analyzed by Southern blot hybridization, using
32P- or digoxygenin-labeled probes shown in Figures 2 and 3.
Southern blots were probed with BH1415 (A and B), Nj:1 (B), YL1 (C), and PX1029 (D). (A) DNA from allelesP:n46(lanes 1 and 2) andSc:124(lanes 3 and 4) were digested withBamHI (lanes 1 and 3) orEcoRI (lanes 2 and 4) and hybridized with digoxygenin-labeled YL1 probe. (B) DNA was digested with BamHI andHindIII and hybridized with32P-labeled PX1029.
The alleles included are Sc:124, Sc:n6008, and P:n46. The repetitive region from probe PX1029 is designatedBnotand is presumed to be a novel maize transposable element. (C) DNA was digested withHindIII and hybridized with32P-labeled
Nj:1 (lanes 1–5) or BH1415 (lanes 6–10). Alleles included in lanes 1–5, and repeated in lanes 6–10, are as follows:Sc:124, R-st,R-r Lc,R-g:1, andP:n46. Thergene composition for these alleles is as follows:Sc:124, [Sc];R-st, [Sc, Nc1,Nc2,Nc3];R-r Lc, [P,q,S1,S2,Lc];R-g:1, [q,S1,S2]; andP:n46, [P]. Band assignment shown is based on published results forR-r:stdand R-st(Robbinset al. 1991;Egglestonet al. 1995). (D) DNA was digested withHindIII and hybridized with32P-labeled BH1415.
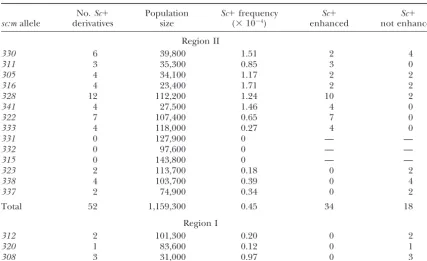
322 7 107,400 0.65 7 0
333 4 118,000 0.27 4 0
331 0 127,900 0 — —
332 0 97,600 0 — —
315 0 143,800 0 — —
323 2 113,700 0.18 0 2
338 4 103,700 0.39 0 4
337 2 74,900 0.34 0 2
Total 52 1,159,300 0.45 34 18
Region I
312 2 101,300 0.20 0 2
320 1 83,600 0.12 0 1
308 3 31,000 0.97 0 3
Sc⫹derivatives are isolated as colored seed fromG1-w22 sc:m mst1-w22/g1-1 P Mst1-1heterozygotes orG1-w22 sc:m mst1-w22/g1-1(P,p:m1orp:m3)Mst1-1.sc:mare ordered by their position inSc(from 5⬘to 3⬘).
(Lin et al. 1999). Overlap among the four sequences A novel result emerges.Sc⫹ derivatives from eight of the group IIsc:mare of two distinct phenotypic classes. defines a total length of 82 amino acids. These
se-quences were identified as non-LTR-transposable ele- The first class, Scp(⫹), is indistinguishable fromSc:124
crossover-derivedSc⫹ alleles from group I alleles and ments with a range in homology to theBnotopen
read-ing frame. Expect values of 0.26–4.8 with 76–50% Sc⫹revertants isolated in the presence ofAc. The sec-ond class,Scp(e) or enhanced pigment alleles, has the protein sequence similarity were produced for the four
database sequences. darkly pigmented aleurone and scutellum ofSc:124but
also has strongly pigmented coleoptile and scutellar Recombination insc:m/Pheterozygotes involving the
group IIsc:malleles produces a unique class ofScderiva- node, unique fromSc:124, from the immediate progeni-torssc:mandP, and from otherSc⫹derivatives. Figure tives:Initial populations of progeny fromsc:m/P
hetero-zygotes produced a dichotomy between the derivatives 1, C and D, shows the phenotypes of representative members of both classes of Scpalleles.
of group Ivs. group II sc:m mutants. In these
experi-ments, few Sc⫹ were obtained from group II (NCO- TheScp(⫹)andScp(e)phenotypes are due to differ-ences in gene conversion events in sc:m/P heterozy-generating) alleles. To determine a possible unique
re-lationship between derivatives from group Ivs. group gotes: The structures of theScpderivatives were deter-mined using PCR amplification and DNA sequence IIsc:m and the basis for the difference between these
groups, larger populations of progeny kernels were gen- analysis. DNA from several derivatives of both Scp(⫹)
andScp(e)classes were amplified via PCR using primers erated from group II mutants. Two types of parental
populations were used. These included sc:m/P (n19, F17 and B2261 and sequenced directly. A total of six
Scp(⫹) and five Scp(e) alleles were sequenced. The
se-n46, orn142) as described in Table 1 andsc:m/p:m::Ds
(p:m3 or p:m1). The latter populations, in which a Ds quences of representative derivatives and progenitor alleles are shown in Figure 6. In all Scp(⫹) that were insertion was located in the 3⬘region ofP(see diagram
in Figure 3), cannot generateSc⫹products by recipro- sequenced, theDs6element is deleted, consistent with the revertantSc⫹phenotype. In each case, theDs6 ele-cal recombination and were excluded from the data
presented in Table 1. A total of 52Sc⫹were generated ment as well as the 8-bp target site duplication is excised precisely, restoring the progenitor Sc:124sequence in fromsc:m/Porsc:m/p:mheterozygotes. Table 2
summa-rizes these data for individual alleles in group II mutants the 517 bp flanking the original site of Ds6 insertion. In contrast, in addition to the loss of Ds6, the Scp(e)
Figure6.—The structure ofScpderivative alleles. Shown are the sequences surrounding theDs6 insertion site (A) and the position of theBnotelement (B) for two of thesc:malleles (sc:m305andm330), the progenitor allele (Sc:124), oneScp(⫹)type derivative for eachsc:m[Sc:n5993 (m305)andSc:n6732 (m330)], oneScp(e)type derivative for eachsc:m[Sc:n5992(m305) and Sc:n7053(m330)], andP. The underlined sequences are the 8-bp target site duplication caused by the insertion ofDs6(A) and the 22-bp direct repeat flanking theBnotelement (B). Boxes denote the entire 2.1-kbpDs6element or the 471-bpBnotelement.
derivatives contain precise deletions of Bnot and one (m305, m311, m316, m322, m328, m330, m333, and
m341). Three insertions located 3⬘of theBnotelement copy of the 22-bp direct repeat flanking it. Thus, there
is a one-for-one correlation between enhanced expres- in a 416-bp segment that is not present inP:A(segment 4, Figure 4) produced noSc⫹derivatives (m315,m331, sion of theScp(e)alleles and deletion ofBnot. Production
of Scp(⫹)and production ofScp(e) derivatives are pre- and m332). Three alleles that were located 3⬘ of the
Sc:A region produced only Scp(⫹) derivatives (m323, sumed to occur by the same mechanism, gene
conver-sion events in which the recombinational intermediate m337, andm338). can be repaired in two ways, one deleting only Ds6
[Scp(⫹)] and the other deletingDs6andBnot[Scp(e)].
DISCUSSION
A conversion tract of at least 200 bp is necessary to
remove bothBnotandDs6from thesc:m. To determine The maizer locus is unique among plant genes be-if theYL1region was included in the conversion tract, cause of the diverse array of expression patterns of vari-primers F4 and B2261 were used to amplify DNA from ant r alleles affecting nearly all parts of the seed and the same 11 Scp derivatives. The presence of the YL1 plant. r allele phenotypic diversity is the result of the
sequence in any of the Scp derivatives would indicate complex structure ofralleles, including variation among coconversions of both this region and theBnotregion the genes in an
rhaplotype and regulatory complexity during meiotic recombination. In each case, amplifica- of single transcriptional unitr genes (Kermicle1985, tion of a 1.5-kbp product [Scp(⫹)] or a 1-kbp product 1988;Robbinset al.1991;Egglestonet al.1995). Two [Scp(e)] indicated that YL1 was not present in the Scp such divergentralleles with nonoverlapping tissue
speci-alleles (not shown). ficity areSc(seed color) andP(plant color). The approach
Thus, a region of the Sc gene containing Ds inser- of this research is to use mutations of these alleles to tional mutants is able to undergo gene conversion with elucidate aspects of plant intragenic recombination and a proximal segment of homology located upstream of gene regulation.
the promoter of thePgene fromR-r:std. The positions We have used heteroallelic combinations ofPand 39 of theDs6insertions and the types and numbers ofSc⫹ Ds6insertion mutations (sc:m) in the maizeSc:124allele
derivatives that are produced from sc:m alleles show a to recover colored seed (Sc⫹) progeny. Three principal strong correlation (Figure 2 and Table 2). Of the 14 results emerge from these studies.
mapped group IIsc:m alleles, the 8sc:m for which the
insertion is located within or 5⬘ of the Bnot element 1. A recombinational dichotomy occurs at a position ⵑ370–470 bp from the start ofrtranscription. sc:m
rectly with heteroduplex formation for nonidentical se-deleting either one or two polymorphic sites from
quences (reviewed inModrichandLahue1996;Chen
the sc:m (Ds6alone or Ds6 and the Bnotelement).
andJinks-Robertson1999). Deletion of Bnotfrom the sc:m alleles produces
al-Neither of these models for the recombinational di-tered tissue specificity of anthocyanin deposition,
chotomy between group I and group II mutants is consis-unique from either progenitor allele.
tent with our data. If the position ofP:Awere the basis Structural explanations for the recombinational be- for the nearly exclusive production of gene convertants havior ofsc:m/Pheterozygotes:Two discrete recombi- from group II mutants, these structural parameters are nationally based regions ofScare demarcated at a point not predicted to affect recombination that involved in-between the m337 and m312 insertion sites, near the serts located within or near the transcribed region of start ofrgene translation.sc:m/Pcombinations involv- Sc. Three mutants (m323,m337, andm338) located near ing group I alleles result in reciprocal crossover events theScpromoter produce primarily convertant progeny. at rates similar to intragenic recombination frequencies It is, therefore, difficult to explain a recombinational for other genes in maize and with double crossover dichotomy using the distribution of insertion/deletion frequencies consistent with expected values (Kermicle differences betweenScandP. Similarly, single-base
het-1970b). The total length of the region delimited by the erologies and insertion-deletion differences are scat-3⬘mutants isⵑ26 kbp/cM (J. Kermicle, M.Alleman tered through Sc and P with no apparent positional
and C. Illingworth, unpublished data). This length dichotomy separating group II and group I alleles. is consistent with the published range of 14–50 kbp/ Transposition of Ds6 or Bnot as an explanation for cM for maize genic regions (Dooner and Kermicle production of Scpderivatives: An alternative
explana-1986; Brown and Sundarasan 1991; Civardi et al. tion for the production of Scp alleles from 5⬘-located
1994;DoonerandMartinez-Ferez1997;Okagakiand sc:mis that eitherDs6orBnotexcision occurs by
transpo-Weil1997). Thus, the unusual question presented by sition. Transposition ofDs6in thesc:m/Pheterozygotes these data is what property ofrallelesScandP causes is highly unlikely in the absence of autonomous element the nearly exclusive production of noncrossover Sc⫹ Ac.sc:malleles are completely stable as homozygotes in
from the group IIsc:malleles. the absence of the autonomous transposable element
One explanation for the recombinational dichotomy Ac, as described forDsinsertion mutants in other maize observed in these data is that DNA structure or sequence genes (McClintock1951;Kermicleet al.1989;Scott influences crossing over insc:m/Pheterozygotes. Possi- et al.1996). TheScpderivatives occur exclusively as prog-ble structural parameters might include the position of eny of somatically stablesc:m/Pheterozygotes. In addi-insertion/deletion differences betweenScandPor the tion, a Ds6 excision footprint, characteristic of typical distribution of single-base-pair polymorphisms. The lo- Ds or Actransposition events, is not present in any of cation ofP:Ais unknown but is at least 15 kbp proximal the Scpalleles that were sequenced (Fedoroff1989). to theP structural gene. Because of this distance, P:A Bnotis a novel maize transposable element of the non-may behave as an ectopic segment. According to this LTR class of retroelements but its excision by transposi-model, recombination events involving P:A preferen- tion does not contribute to our recombination data. tially produce noncrossover progeny whereas recombi- Non-LTR retroelements transpose extremely rarely nation events between “homologous” regions would (Schmidt1999). To useBnottransposition to explain produce primarily crossover progeny. Ectopic recombi- the existence of Scp(e) derivatives, transposition must nation has been studied extensively in yeast and in mam- occur frequently, only insc:m/Pheterozygotes, and only mals and is shown to proceed through normal meiotic for alleles in the 5⬘ end of the Sc allele. Excision of pathways (Lichtenet al.1987;KupiecandPetes1988; Bnotdoes not occur duringAc-induced reversion ofsc:m
unpublished data). In addition,Bnotexcision does not same position as for Sc:124 and containing YL1 but lackingBnot( J.Bernotand M.Alleman, unpublished occur insc:m/Pheterozygotes involving group I alleles
(not shown). We believe that, althoughBnotis a trans- data).
In addition, the flanking region ofPcontains several posable element, its transposition is rare or nonexistent
and does not contribute to these recombinationally known repetitive elements, including ones with homol-ogy to the maize retroviral element Ji-6 (SanMiguel
based data.
Production ofScpderivatives by heteroduplex repair et al. 1996). LTR and non-LTR retrotransposons are ubiquitous elements in plants, contributing strongly to during gene conversion at r:A second aspect of these
results bears on the types of products obtained via re- the repetitive fraction of the genome (reviewed in
Schmidt1999). Retroviral-related transposons have ex-combination involving differentsc:m alleles.
Heteroal-lelic combinations of sc:m andP presumably produce panded the intergenic regions and changed the se-quence context of the maize gene as compared to the
Sc⫹derivatives via the formation and repair of
recombi-national intermediates (heteroduplex) involving the two more compact genomes of rice and sorghum (White
et al.1994;Bennetzenet al.1998). Through the study alleles. The proposed heteroduplex tract would include
sequences fromsc:mand eitherPorP:Adepending on ofrand other related genes in plants, we are developing a view of gene regulatory evolution to include the oppor-the location of oppor-the sc:m insert. sc:m mutants located
withinSc:Aand producing bothScp(⫹)andScp(e)deriva- tunistic use of transposable element insertions and ge-nomic recombination events.
tives are informative. These sc:m (m305, m311, m316,
m328, andm330) are able to generateScp(⫹)orScp(e) We thank Dr. Timothy Robbins for theR4.7 clone. We also thank
alleles. The two sites are located from 108 to 159 bp Beverly Oashgar, David Heller, and Darla West for technical assistance. United States Department of Agriculture grant 93-37301-8878 (M.A.), apart dependent on thesc:mallele used (Figure 2). Our
National Science Foundation grant 9603747 (M.A.), and Department data do not require discontinuous conversion tracts,
of Energy grant DE-FG02-86ER-13539 ( J.L.K.) supported this work. only the formation of two types of recombinational
in-termediates or differential repair of polymorphic sites. It is possible that there is either incomplete repair of the
LITERATURE CITED
heteroduplex or initiation of DSB in different positions.
The reciprocal event, transfer ofDs6andBnottoP:A, or Alleman, M., and J.Doctor, 2000 Genomic imprinting in plants: observations and evolutionary implications. Plant Mol. Biol.43:
loss ofBnotalone, could not be identified in our screen.
147–161.
Generation of gene regulatory differences by trans- Alleman, M., and J. L. Kermicle, 1993 Somatic variegation and germinal mutability reflect the position of the transposable ele-posable element insertion, genetic recombination, and
mentDissociationwithin the maizeRgene. Genetics135:189–203. chromosomal rearrangement: These data support the
Altschul, S. F., T. L.Madden, A. A.Scha¨ffer, J.Zhang, Z.Zhang evolution of maize genes by transposable element inser- et al., 1997 Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res.25:3389– tion, rearrangement, deletion, and genetic
recombina-3402. tion. Most of the first 3.5 kbp 5⬘of the start of
transcrip-Bennetzen, J. L., P.SanMiguel, M.Chen, A.Tikhonov, M.Francki tion of Scand P are nonhomologous, consistent with et al., 1998 Grass genomes. Proc. Natl. Acad. Sci. USA95:1975–
1978. the structure of alleles of the maize b (booster-1) gene.
Borts, R. H., and J. E.Haber, 1987 Meiotic recombination in yeast: Two well-characterized alleles,B-Peru (strong seed
pig-alteration by multiple heterozygosities. Science237:1459–1465. mentation) and B-I (strong leaf blade pigmentation), Borts, R. H., and J. E.Haber, 1989 Length and distribution of meiotic gene conversion tracts and crossovers inSaccharomyces
although homologous in coding region sequence, are
cerevisiae. Genetics123:69–80. highly divergent in the 5⬘ flanking sequences (
Chan-Brink, R. A., 1958 Paramutation at theRlocus in maize. Cold Spring
dleret al.1989;Radicellaet al.1992). Harbor Symp. Quant. Biol.23:379–391.
Brown, J., and V.Sundarasan, 1991 A recombinational hotspot in A putative P-like progenitor would have undergone
the maizea1intragenic region. Theor. Appl. Genet.81:185–188. rearrangement to bring theP:Aregion into contact with
Burr, F. A., B.Burr, B. E.Scheffler, M.Blewitt, U.Wienandet al., thergene, deletion of theYL1region, and insertion of 1996 The maize repressor-like geneintensifier1shares homology with ther1/b1multigene family of transcription factors and exhib-theBnottransposable element during maize evolution.
its missplicing. Plant Cell8:1249–1259. Because a P phenotype appears in most teosinte lines
Chandler, V. L., J. P.Radicella, T. P.Robbins, J.Chenand D. tested thus far (Hansonet al.1996), it is presumed that Turks, 1989 Two regulatory genes of the maize anthocyanin pathway are homologous: isolation ofButilizingRgenomic se-a P gene represents ther gene progenitor. The
inser-quences. Plant Cell1:1175–1183. tion of Bnot into Sc is assumed to have changed the
Chandler, V. L., W. E.Egglestonand J. E.Dorweiler, 2000 Para-phenotype from strongly expressed in the coleoptile mutation in maize. Plant Mol. Biol.43:121–145.
Chen, J., I. M.Greenblattand S. L.Dellaporta, 1992 Molecular and scutellar node to theSc:124phenotype, weakly
pig-analysis ofActransposition and DNA replication. Genetics130:
mented coleoptile and colorless scutellar node. This
665–676.
hypothesis is consistent with the structure and pheno- Chen, W., and S.Jinks-Robertson, 1999 The role of the mismatch repair machinery in regulating mitotic and meiotic recombina-type of another naturally occurring variant allele, the
tion between diverged sequences in yeast. Genetics151:1299–
Scm gene from R-marbled (R-mb;Weyers 1961). Scm is
1313.
McClintock, B., 1951 Chromosome organization and genic
expres-Dsaffects the pattern of intragenic recombination at thebzand
sion. Cold Spring Harbor Symp. Quant. Biol.16:13–47.
Rloci in maize. Genetics113:135–143.
McWhirter, K. S., and R. A. Brink, 1962 Continuous variation
Dooner, H. K., and I. M.Martinez-Ferez, 1997 Recombination
in level of paramutation at the Rlocus in maize. Genetics47:
occurs uniformly within the bronze gene, a meiotic
recombina-1053–1074. tion hotspot in the maize genome. Plant Cell9:1633–1646.
Modrich, P., and R.Lahue, 1996 Mismatch repair in replication
Dooner, H. K., and E. J.Ralston, 1990 Effect of theMu1insertion
fidelity, genetic recombination, and cancer biology. Annu. Rev. on intragenic recombination at the bz locus in maize. Maydica
Biochem.65:101–133.
35:333–337.
Moreno, M. A., J.Chen, I. M.Greenblattand S. L.Dellaporta,
Dooner, H. K., T. P.Robbinsand R. A.Jorgensen, 1991 Genetic 1992 Reconstitutional mutagenesis of the maizePgene by short-and developmental control of anthocyanin biosynthesis. Annu. rangeActranspositions. Genetics131:939–956.
Rev. Genet.25:173–199. Nag, D. K., and T. D.Petes, 1990 Meiotic recombination between
Eggleston, W. B., M.Allemanand J. L.Kermicle, 1995 Molecular dispersed repeated genes is associated with heteroduplex forma-organization and bases for germinal instability inR-stippledmaize. tion. Mol. Cell. Biol.10:4420–4423.
Genetics141:347–360. Ochman, H., A. S.Gerberand D. L.Hartl, 1988 Genetic
applica-Fedoroff, N. V., 1989 Maize transposable elements, pp. 375–411 tion of an inverse polymerase chain reaction. Genetics120:621– inMobile DNA, edited by D. E.Bergand M. M.Howe. American 623.
Society for Microbiology, Washington DC. Okagaki, R. J., and C. F.Weil, 1997 Analysis of recombination sites
Goff, S. A., K. C.Coneand V. L.Chandler, 1992 Functional analysis within the maizewaxy-1locus. Genetics147:815–821. of the transcriptional activator encoded by the maize Bgene: Petes, T. D., R. E.Maloneand L. S.Symington, 1991 Recombina-evidence for a direct functional interaction between two classes tion in yeast, pp. 407–521 inThe Molecular and Cellular Biology of of regulatory proteins. Genes Dev.6:864–875. the Yeast Saccharomyces, Vol. 1. Genome Dynamics, Protein Synthesis,
Hanson, M. A., B. S.Gaut, A. O.Stec, S. I.Fuerstenberg, M. M. and Energetics, edited by J. R. Broach, E. W.Jones and J. R.
Goodmanet al., 1996 Evolution of anthocyanin biosynthesis in Pringle. Cold Spring Harbor Laboratory Press, Cold Spring Har-maize kernels: the role of regulatory and enzymatic loci. Genetics bor, NY.
143:1395–1407. Radicella, J. P., D.Turksand V. L.Chandler, 1991 Cloning and
Jackson, J. A., and G. R.Fink, 1981 Gene conversion between dupli- nucleotide sequence of a cDNA encodingB-Peru, a regulatory cated genetic elements in yeast. Nature292:306–311. protein of the anthocyanin pathway in maize. Plant Mol. Biol.
17:127–130.
Kermicle, J. L., 1970a Dependence of theR-mottled aleurone
phe-Radicella, J. P., D.Brown, L. A.Tolarand V. L.Chandler, 1992 notype in maize on mode of sexual transmission. Genetics66:
Allelic diversity of the maizeBregulatory gene: different leader 69–85.
and promoter sequences of twoBalleles determine distinct
tissue-Kermicle, J. L., 1970b Somatic and meiotic instability ofR-stippled,
specificities of anthocyanin production. Genes Dev.6:2152–2164. an aleurone spotting factor in maize. Genetics64:247–258.
Robbins, T. P., E. L.Walker, J. L.Kermicle, M.Allemanand S. L.
Kermicle, J. L., 1980 Probing the component structure of a maize
Dellaporta, 1991 Meiotic instability of theR-rcomplex arising gene with transposable elements. Science208:1457–1459.
from displaced intragenic exchange and intrachromosomal
re-Kermicle, J. L., 1985 Alternative tests of allelism, pp. 491–507 in
arrangement. Genetics129:271–283.
Plant Genetics, edited by M.Freeling. Alan R. Liss, New York.
SanMiguel, P., A.Tikhonov, Y.-K.Jin, N.Motchoulskaia, D. Zakh-Kermicle, J. L., 1988 Recombinant mutable alleles of the maizeR
arovet al., 1996 Nested retrotransposons in the intergenic re-gene, pp. 81–89 inPlant Transposable Elements, edited by O. E.
gions of the maize genome. Science274:765–768.
Nelson. Plenum, New York.
Schmidt, T., 1999 LINEs, SINEs, and repetitive DNA: non-LTR
ret-Kermicle, J. L., 1996 Epigenetic silencing and activation of a maize
rotransposons in plant genomes. Plant Mol. Biol.40:903–910. r gene, pp. 267–287 inEpigenetic Mechanisms of Gene Regulation,
Schnable, P. S., A.-P.Hsiaand B. J.Nikolau, 1998 Genetic recom-edited by V. E. A.Ruzzo, R. A.Martienssenand A. D.Riggs.
bination in plants. Curr. Opin. Plant Biol.1:123–129. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Scott, L., D.LaFoeand C. F.Weil, 1996 Adjacent sequences
influ-Kermicle, J. L., M.Allemanand S. L.Dellaporta, 1989 Sequential ence DNA repair accompanying transposon excision in maize. mutagenesis of a maize gene using the transposable element Genetics142:237–246.
Dissociation. Genome31:712–716. Steele, D. F., M. E.Morrisand S.Jinks-Robertson, 1991 Allelic
Kermicle, J. L., M.Allemanand W. B.Eggleston, 1995 Paramuta- and ectopic interactions in recombination-defective yeast strains. tion of the maizeRgene depends onRgene copy number and Genetics127:53–60.
arrangement. Genetics141:361–372. Styles, E. D., O.Ceskaand K. T.Seah, 1973 Developmental
differ-Kupiec, M., and T. D.Petes, 1988 Allelic and ectopic recombination ences in action ofRandBalleles in maize. Can. J. Genet. Cytol. betweenTyelements in yeast. Genetics119:549–559. 15:59–72.
Lichten, M., R. H. Bortsand J. E. Haber, 1987 Meiotic gene Symington, L. S., and T. D.Petes, 1988 Expansions and contrac-conversion and crossing over between dispersed homologous tions of the genetic map relative to the physical map of yeast sequences occurs frequently inSaccharomyces cerevisiae. Genetics chromosome III. Mol. Cell. Biol.8:595–604.
115:233–246. Tikhonov, A. P., P. J.SanMiguel, Y.Nakajima, N. M.Gorenstein,
Lin, X., S.Kaul, S.Rounsley, T. P.Shea, M. I.Benitoet al., 1999 J. L.Bennetzenet al., 1999 Colinearity and its exceptions in Sequence and analysis of chromosome 2 of the plantArabidopsis orthologousadhregions of maize and sorghum. Proc. Natl. Acad.
Sci. USA96:7409–7414.
Walker, E. L., T. P.Robbins, T. E.Bureau, J. L.Kermicleand White, S. E., L. F.Haberaand S. R.Wessler, 1994 Retrotranspo-S. L. Dellaporta, 1995 Transposon-mediated chromosomal sons in the flanking regions of normal plant genes: a role for rearrangements and gene duplications in the formation of the Copia-like elements in the evolution of gene structure and expres-maizeR-rcomplex. EMBO J.14:2350–2363. sion. Proc. Natl. Acad. Sci. USA91:11792–11796.
Wang, L., and S. R.Wessler, 1998 Inefficient reinitiation is respon- Xu, X., A. P.Hsia, L.Zhang, B. J.Nikolauand P. S.Schnable, sible for upstream open reading frame-mediated translational 1995 Meiotic recombination break points resolve at high rates at repression of the maizeRgene. Plant Cell10:1733–1746. the 5⬘end of a maize coding sequence. Plant Cell12:2151–2161.
Weyers, W. H., 1961 Expression and stability of the marbled allele