Copyright 0 1996 by the Genetics Society of A m e r i c a
Mitochondrial Intronic Open Reading Frames
in
Podospora:
Mobility and Consecutive Exonic Sequence Variations
Carole H.
Sellem, Yves d'Aubenton-Carafa, Michele Rossignol and Leon Belcour
Centre de Ginitique Moliculaire, Centre National de la Recherche Scientijque, 91 198 Gif-sur-Yvette, France Manuscript received July 17, 1995
Accepted for publication February 22, 1996
ABSTRACT
The mitochondrial genome of 23 wild-type strains belonging to three different species of the filamen- tous fungus Podospora was examined. Among the 15 optional sequences identified are two intronic reading frames, nadl-i4-orfl and coxl-i7~$. We show that the presence of these sequences was strictly correlated with tightly clustered nucleotide substitutions in the adjacent exon. This correlation applies to the presence or absence of closely related open reading frames (ORFs), found at the same genetic locations, in all the Pyrenomycete genera examined. The recent gain of these optional ORFs in the evolution of the genus Podospora probably account for such sequence differences. In the homoplasmic progeny from heteroplasmons constructed between Podospora strains differing by the presence of these optional ORFs, nadl-iCorfl and coxl-i7-orf2appeared highly invasive. Sequence comparisons in the nadl- i4 intron of various strains of the Pyrenomycete family led us to propose a scenario of its evolution that includes several events of loss and gain of intronic ORFs. These results strongly reinforce the idea that group I intronic ORFs are mobile elements and that their transfer, and comcomitant modification of the adjacent exon, could participate in the modular evolution of mitochondrial genomes.
G
ROUP I introns are nonessential genetic elements. Most confer no apparent selective advantage at the individual level (SERAPHIN et al. 198'7) but have spread in the course of evolution because endowed with invasive properties imparted by the product of the open reading frames (ORFs) they contain (LAMBOWITZ and BELFORT 1993). Their homing from intron-containing gene to cognate intronless allele is dependent upon this product, a site-specific endonuclease that creates a double-strand break at the exon-exon junction of the intronless allele ( JACQUIER and DUJON 1985; MACREA- DIE et al. 1985; COLLEAUX et al. 1986).The two distinct domains of mobile group I introns (catalytic core and endonuclease-encoding ORF) are supposed to have evolved independently ( LAMBOWITZ
and BELFORT 1993). The presence of related intronic ORFs in divergent organisms was already proposed to be the result of genetic exchanges involving the ORF
sequence independently of the intron (MICHEL and DU- .JON 1986). Likewise, the presence in highly conserved
introns of ORFs potentially encoding endonucleases that vary in both sequence and position (i.e,, CUMMINGS and DOMENICO 1988; MOTA and COLLINS 1988; SHUB
et al. 1988) also strongly suggests that the ORFs rather than the introns were the original mobile elements
( BELL-PEDERSEN et al. 1990). Moreover, OW-encoding, or potentially encoding, endonucleases similar to those found in group I introns have been identified in various
Corresponding author: C. H. Sellem, Centre de GCnCtique MolCcu- laire, C.N.R.S., 1, avenue de la Terrasse, 91198 Gifsur-Yvette, France. E-mail: [email protected]
Genetics 1 4 3 777-788 (June, 1996)
other contexts such as in an archeal intron (DALGAARD et al. 1993) and as free standing sequences, inside (i.e., the inteins; COOPER and STEVENS 1995) or outside
(SHARMA et al. 1992; PEL and GRIVEL 1993; PAQUIN et al. 1994) gene context. In some of these cases, the ORFs
were described as optional and proposed to be recent acquisitions, but their mobility as independent genetic entities has only been demonstrated in the case of in- teins ( GIMBLE and THORNER 1992). Colonization of pre- existing group I introns by independent mobile ORFs is supported by a few examples of optional ORFs (MOTA
and COLLINS 1988; BECHHOFER et al. 1994; BINISZKIE-
WICZ et al. 1994; DE JONCKHEERE and BROWN 1994). Different mechanisms of acquisition have been pro- posed (LOIZOS et al. 1994; VADER et al. 1994), but no experimental evidence of mobility of such ORFs, from OW-containing intron to cognate OW-less allele, has been described.
The mitochondrial chromosome of the filamentous fungus Podospora ansm'na (P. an) contains numerous introns (up to 36 in the reference wild-type strain P. anA) , some of which are optional (L. BELCOUR, unpub- lished results). Among the optional sequences are two ORFs potentially encoding highly specific DNA endo- nucleases, located within two group I introns: the fourth intron of the nadl gene (nadl-i4) and the seventh in- tron of the coxl gene (coxl-i7), in the reference wild- type strain (CUMMINGS et al. 1990). Both introns have the peculiarity of containing either one (mono-orfic introns) or two (biorfic introns) ORFs depending on the wild-type isolate. We have examined the nadl and
778
C.
H. Sellem et al.family: P. comata (P. eo) , P. curvicolla (P. cu) and Sordaria macrospora ( S . ma). Three organizations were evidenced for both genes: intronless, mono- and biorfic. Sequence analysis revealed a systematic correlation between the presence/absence of the optional ORFs and a consen- sus sequence variation in the few adjacent nucleotides of the neighbouring exons (nadl-e4 and coxl-e8). This correlation applies to the various organizations found for the equivalent loci in Neurospora (BURGER et al.
1982; MOTA and COLLINS 1988; FIELD et al. 1989; HAWSE
et al. 1990). We also took advantage of the favorable situation in Podospora to study the behavior of the two optional intronic ORFs in heteroplasmons and demon- strated their efficient invasiveness. The data we present here led us to propose that these ORFs are recent acqui- sitions by the Podospora nadl-i4 and coxl-i7introns and that their arrival was responsible for the modifications in the exonic sequences and thus contributed to mito- chondrial genome evolution.
MATERTALS AND METHODS
Strains: P. ansm’na and P. comata are pseudohomothallic four-spored species that give some viable progenies when crossed one to each other. Most of the P. anserina wild-type strains used in this work (A, B, C, D, E, F, H, M, N, R, S, U, W, X , Z) are French isolates provided by D. MARCOU. Two wild-type strains of P. comata were obtained from the ATCC: P. c02 (no. 26491, Uganda) and P. co3 (no. 36713, Venezuela), a colorless and totally sterile strain incompatible with any others. P. coT was a French isolate. P. curvicolla is a homothallic 128-spored species. P. CUM was given by J. BE- GUERET. P. cuL and P. cuVwere French isolates. S. macrospora strain was obtained from Dr. D. ZICKLER (ATCC no. 60255).
Construction of isogenic strains: P. ansaina wild-type strains A, M and D (P. anA, P. anM, P. anD) were used as the female in crosses with the wild-type strain P. anS. In the prog- eny, the strains that did not present a “barrage” with the P. anS strains were crossed again with P. anS as the male. Two cycles of backcrossing gave rise to strains containing a P. anS like nucleus and either P. anA, P. anM or P. anD mitochon- dria. In the case of interspecific crosses, the P. coT wild-type strain was used as the male with P. anS as the female. Ten cycles of isogenization gave rise to strains containing P. anS mitochondria and rather pure P. coT nuclei. In all the cases, the parental strains used in heterokaryon construction were named according to the origin of their mitochondria.
Construction of heterokaryons: All the heterokaryons were constructed between pairs of strains of opposite mating type. Implants of growing mycelia of the OW-less strains (recipi- ents: P. nnM, P. anD or P. COT) were placed on Petri dishes containing rich medium -1 mm in front of the growing thal- lus of the OW-containing strains (donors: P. anA or P. anS). When the mycelia had invaded the Petri dishes, they were exposed to light and fruiting bodies were allowed to develop. To study independent recombination events, a single ascus was collected from each heteroplasmic cultures. Indeed, in Podospora heteroplasmic mycelia were shown to produce only homoplasmic ascospores (BELCOUR and BEGEL 1977). Mycelia issued from 18 to 48 asci were further analyzed in each experiment.
Analysis of mitochondrial DNA: Mitochondrial DNAs were extracted from various Podospora wild-type, isogenic- or h e moplasmic-derived strains by a minipreparation method (LEc
ELLIER and SILAR 1994). Southern blot experiments were per- formed after Hue111 or HinJ digestions, on Hybond nylon membranes, using 32P-labeled synthetic oligonucleotides. As-
cobolus immersus total DNA was a gift from J. L. ROSSIGNOL. Sequencing: PCR amplifications were performed using pairs of synthetic oligonucleotides as primers. The fragments obtained were cloned in a pMOSBZue T-vector (Amersham) and the junctions sequenced using the PCR primers. The whole P. co3 nadl-i4 intron was sequenced on three indepen- dent clones to resolve the mistakes introduced by the Taq polymerase (Appligene); the P. COT, P. anMand S.ma6 nadl-i4 introns were cloned from restriction endonuclease fragments and sequenced either using specific oligonucleotide primers or using the nested deletion kit (Pharmacia) and the universal sequencing primers. The sequences have been entered into the EMBL bank under the number 250030 for P. COT and 250031 for P. co3 nadl-i4 introns.
Sequence data: In all the strains of P. ansm’na and in strains of other species and genera, nadl-i4 and coxl-i7 equivalent introns, inserted at the same genomic location, are not neces- sarily the fourth or the seventh intron but will nevertheless be refered as n u d l 4 and coxl-i7 for simplicity. Neurospora and Aspergillus nadl and cox1 mitochondrial sequences, con- taining or not the nadl-i4 and coxl-i7equivalent introns, were obtained from published data (see legend of Figure 1). Pairs of sequences were aligned using the BESTFIT program from the UWGCG package with a gap penalty of 5 . Phylogenetic tree was built using DNADIST, FITCH and DRAWTREE pro- grams from the PHYLIP package. The Lipman-Pearson algo- rithm (FASTA) was used for comparisons with EMBL (release 58) data banks.
RESULTS
The nadi4 and coxli7 introns in P. anserim- Mito- chondrial DNA extracted from
17
wild-type strains of P. anserina was investigated by Southern blotting. Thenadl-i4 and coxl-i7 introns, reported to be biorfic in the P. anserina wild-type strains A and s (CUMMINCS et al. 1988, 1989) may present a different organization: in place of two ORFs (Figure 1, A and A‘) only one was found in several strains (Figure 1, B and B’). The n a d l - i4-orfI and the coxl-i7-orf2 are optional in P. ansm’na.
We have determined the nucleotide sequence of short DNA fragments (-200 bp) containing the n a d l - e4/i4 and the coxl-i7/e8 junctions, after PCR amplifica- tions of the mitochondrial DNA extracted from 10 wild- type strains of P. anserina (Figure 2, lines 4, 5 and 10,
11). For each region, all the strains displaying the bior- fic intron presented a junction sequence identical to those already published (CUMMINGS et al. 1990). The homologous regions in strains with a mono-orfic organi- zation were found identical to each other, but in the exon adjacent to the optional ORF (the upstream one of nadl-i4 and the downstream one of coxl-i7), a high level of divergence was observed with the correspond- ing sequence of the biorfic intron-containing gene. In- deed, nine and six substitutions are clustered in the last 28 nucleotides of the nadl-i4 upstream exon and in the first 18 nucleotides of the coxl-i7 downstream exon, respectively. This result was further extended to the 17
Mobile Intronic ORFs in Podospora
-
n a d l - i 4*
1 1 1 1
-
64 A
779
A
B
C
D
4 A
I n t r o n S p e c i e s
O r g a n i z a t i o n P . a n s r r i n a P . c n n r a r a P . c u r v i c u l l o O r h c r s
A A , R , E , N , H , 3 s , ,,1 R 6
A'
I
ORFl A ORF24 A
C'
FIGURE 1.-Organization of the n n d l and cox1 loci in various Pyrenomycete strains. The introns present at the same genetic location as P. nnA nndl-i4 and coxl-i7 are either biorfic (A and A'), mono- or pseudomono-orfic ( B and R' or D) introns. The structure in which the intron is absent from this position is also represented ( C and C'). The 5'-, 3"constitutive
(A)
and 3'- alternative ( A ) splicc sites of the introns are indicated; their catalytic core is represented hy a black dot ( 0 ) . Exons are large hoxes divided into several cross-hatched hlocks representing the regions o f polymorphic variations. Thc number of nucleotides in each block is indicated. Open bars indicate the upstream (ORFI) and the downstream (ORF2) intronic reading frames. The first ORFs are in frame with the upstream exon, show the two WGI.1-DADG dodecapeptides motifs, and loop out of the PI and P8 seconday structures o f the intron ( n n d l - i C o f 1 and coxl-i7-ofl, respectively). The second ORFs show either the GIY-MG or LAGLI-DADG motifs ( n n d l - i 4 - o r - and coxl-i7-orf2 respectively) laying in or downstream from the P9-1 secondary structure of the intron and require alternative splicing to he expressed. Bold arrows surround directly repeated exonic or intronic sequences hordering optional ORFs. The 17-hp (hlocks 11+6) sequence, found clownstream from the optional nc~dl-iCoyfl (A), is an exact replica of the 3' end of the upstream exon hording the nndl-i4 mono-orfic intron ( B ) . It contains a CFhp invariant sequence corresponding to the 5' side of the P1 stem of the secondary structure of the intron. The 8-hp sequence found in the 5' end of the optional c o x l - i 7 - m - is an exact replica of the downstream intronexon junction; this replicated sequence contains the 3' side of the P9-0 and P10 stems in the seconday structure of the intron and thus provides an alternative 3"splice site. The inset tahle indicates the organization of the introns for 17 P. nnsm'nn ( A...a,
three P. comntn (7; 2, 3) and three 1'. curuicolla (114, I-,19 strains. The intron organization for strains in other genera was either deduced by sequencing from S. mnrrosporn strain 6025.5 (S. mn6; C. H. SELLEM, unpuhlished result5) and A . immwsus ( A . im;
(3.
H. SI:.I.I.IM, unpuhlished results) or ohtained from the literature: N. mmsn strains 74A (A'. 0 - 7 ; BURGER Pt nl. 1982; BURGER and WERNER 1985), AOOott 12 ( N . MAh; HAWSE ~t nl. 1990), Adiopodoume ( N . nAd; F1EI.D ~t nl. 1989), N. i n l m d i n strain Vnrkzcd ( N . inL: MOTA and COI.I.INS 1988), A . nidtclons ( A . ni; BROWN Pt 01. 1983; WARING PI nl. 1984). Mnrchnntiu po!ynorf)tw (IM. Po; OTHA d nl. 1993).tory, by Southern blot experiments, using exon-specific probes designed from the hvo types of exonic sequences
(Figure 3). A strict correlation has thus been clearly demonstrated in P. nnsm'nn between the presence of
two group I intron optional ORFs and consensus nucle- otide substitutions in their neighboring exon.
For the purpose of comparing further mono- and biorfic introns, we have determined the sequence of the whole nndl-i4 and part of the coxl-i7 mono-orfic introns, as well as large parts of the adjacent exons, in
the wild-type strains P. anM and I-'. ann, respectively. It appears that the sequences absent from nudl44 and coxl-i7 mono-orfic introns contain the optional ORFs (391 and 325 encoded amino acids) and, respectivelv,
292 and 49 nucleotides downstream from their stop codon. In the case of nndl, over the 250 nucleotides of
n a d l - ~ 4 , the first 70 nucleotides of nod195 and 1129
m m l d 0 4
c m u u
y.0.
o m 0u u m e m e
;.A nu um um mm m m m
; z 2 2
a
E"
c a 4
.
.
.
.
.
.
.
;I
z-4
E5 5 5 5
.
.
.
.
.
.
.
u - E E E E h-E
? ? 5 5
u u u JJ rd d m $ t 3 m u
u u
u
.
.
.
.
.
.
.
. . . .
.
.
.
.
.
.
.
.
$ uE
4 ? ? ?
$ - E
? E ? ?
2 JL
E
3
8
E O4
2
44
.
.
.
.
.
.
.
c70
.
.
.
w . . .
.
.
.
.
.
.
.
.
. . .
0. . . .
0 u.
u.
u.
v.
J U ? ?
"u E c) 4 w5 5 5 5
3
x $2
.
. . . .
.
.
.
0
u u u u - B E E E E
E 4 c7 c7
i
5
, ? ? ? g - e E ? ? ? E :
!s
H 4 E2
4 -"u ? ? E : ?
u V ? ? ? ?
<
3
.
.
.
.
.
.
.
. . . .
.
.
.
.
. . . .
5 0 : : :
d B::
E - 4
4 4
2
E V U5 5 ? ?
.
.
.
.
3
8
L* ? ? ? "u-"t,I
4 g
c73
d e ? ?
O 8-9
E : ? ? ?c7
. . . .
;
.+ m4 4 4
3-8
u u u u4
- 3
.
.
.
.
.
.
.
.
E E 0 n
.
.
.
.
.
.
.
.
. . .
. . .
VI LL.
.
.
.
u V
. . .
u u u m u c 7 u c 7 t n E m E +
c) u c 7 b l g
u u u v
u u u c 7
. c ? 5 L ?
-
"0? ? e ?
-
-d uZ U ? ?
? ? ? e
ux m
E;
m u u u u u uu
. . .
. . . .
I
+ + + + I
d d d
0 0 0 d
I l l !
0 rl 0 d0 H r0 l H
Y
2
g
G
I 4-
s!-
rr,-
s
3
Lq
2
3 0G u s ' d
q'
6
6 3 6
5
-
: s
4 \ g
5 5 5 2 <L 5
-Iu
z m c
u u 2 26 4
o ; $ &4
a ; d $ s :."$
$ 0 0 1
I 1 1 X - 1
N F l e ul w1 1~ 1 m1 m
t I
N cv N1 H1 N1 H1
0 0 0 0
-
rr,- h\
3
Y '",b o
4
G '
t;
p j - 2
; ; s
< < : \ a $L
4 2 5
~ G G S ti g d
& L i d <
1
a c
a n u s c u0 u3 uk ok
I I
0 rl 1 1 1 1
d rl dN md edudl
e, 0 0 " " C " e, - e , q . a ; q
* & w Z $ z c 4
d i d
&2
c.=.;zq-
hCb 2 0 - g o
"5s
p a e Q p - . *
8
E2
-E e
id%
Ejkcu - s q
. % E & " -5
2
e,*- 0 v u u pi x ..9 idE Z S
M 2 % 5
id 3 id m - 2=Z;a;.Eg
- 2 2 3
c u h e,+2
y)8
m c bD0Sz;- id2
8
$sz.gp
3
$ + , = U 2 Eg
g
b
%e,
1 r= 1 . q$ 8 c z & 2 8 0 = = v
a & 2 Q +d 3 0 . - e, Q
g
E ;'1g ? ' z o &
6 - c
- 0 2 m c i j L a ; o j 0 -a idu s $ e,
E
g
u 2 Y - S
g L
$
ge,
0 00 s
L 2 "g . Q
o ~ 2 ~ & g a Y ? : E w
v1 b b" 5
u . 2 3 + 0g
Ew i d % o a g
2
-
0 e, U - . - ' c & 4 ; % % a x 42 o j
0Ea
agd-,,
i *
u 2c 0 %
g ;
o Q6
g a g 2
*,-. m 1 I .- Y 2
'2
&-<
g
5
hDYG k q
E m i 0 u c 3 . G C "- 0 s > + c : * ' d a -
a
c.l e, 4 O L E 0 * u O ' v l p.&S,,~'a-. L E
51
8 . 5 3
g z & F c . g
c y 08
% p Q
1 v1 o a & Y , y &1 ag
5 5
g%.gyp$tL
,E
s
5
po;o3
p ; g
$
&-s2
cp z : 2 c ,
y
m Tpgg.$;g
s g 5
EE"$%
o.$ u 1 0 Z T & O @ V
u,
$3
- & G v . - , s 0
&?%g!
8,s
g;
so3,
y E 0e,G
* aJ e,2
c -
2
& G < % V . & $ e &55g
:4jw,o
* Q w cZ O
5 . g
w 0-
L ;;iE
= z HE.2
F: id G E Q G 0 E u .Q O l n L . 2
es
5 s g Z . s ;
*
O B 0 G * s - a . z e, e - idF L * o - 3 q 2
W E& o s G
5
i!E 2
y,c
G27i u.-e, c 6 0 ~ '
g
~E* <
e,
S."
I id o s ffi u -4 5 L 0 c M, C = * Q S . ~ - 3 w w c . u
E
g 13 o s G 0 u Q . -9
z
E * g ~ . 8 ~i e - O W ? " a2;;
g
E < u boa u 4$2."
p & - 0 . 2
m ' z
4
e,
e,- - C W g c ; E ' & + b i
3
2
u
3 2 % m u E.9::: 0 a-e, cz
u - " y3
2
y
T " u $ $ $ N U .-e 0
h g
u Q : 9 - ; 0 2 G Z 62i:$'";&p
- 2 E v a J2
. . s a 2
-
0 ? . $ 8 p E g ! i S E a e , L2-G W E ffi c Q 0 - c
2.5
s . 2 . m Q) g 0 0 m % E c Q.0 A . . S $?
2 . 5
c " . a ;
E -
2
E 7 2
8,s
sp"
g
"_
g . -
a 2 0 - s . E
C ' 36
E T 3g
E a ; rmg
2 s o r d - . 2 3 - & 3& E
EFLk
g.?
39
5z
g+g
~
.*Le2
d
. A c
2 s
g5
e , z
2
@ & %2
% j < O e, -2
Y"~e,b.5c-.-$@;~
u e,m.%&2$
k3bo
E g
g i g
g
; g . $ 2 s
q
a
b n L > 0 0J Q - % - $ q @ -1 , 3 g : s p i d 3
u c Lc 0 . 5 - x S ; E g 3 8 $ %
0.I
i d y a a a s " a . g
';f2a;9
3",-y 0 G
;3 E G - o v .p
5
:.kg2
:, ER 9 S a
2
o Q Z E $ ; . g C $ $
6.2 o s5 %
5 3
3b s
3 S he
uz
q s c
"'21,,
in* o +..Eg
& *
5,-. E , < G i d U
0 ,
v1 0 " a M.E
z
b
g
2 @w $ z y $ g
v e,k G , ~ $ T L Ed
"-
O v i d m m C , E ' T - , & * C "
8
2
g
~
id E E g g.
~
0~
g.5
ECg,&uC
- 4 L Q g
id.-
Mobile Intronic ORFs in Podospora 78 1
R N M H E C B A S
"-
.-
___"-__i-z . ~4 -2265 bp
A
'
+3OO2 bpB
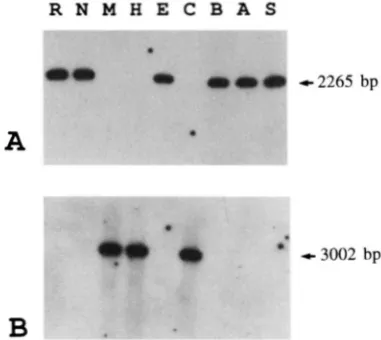
FIGURE 3.-Southern blot analysis of HueIIIdigested mito- chondrial DNAs extracted from nine wild-type strains of P.
anrprinn. "P-labeled oligonucleotide probes complementary
to the 3' end of the P. anA (A) or P. anM (B) nadl-e4 were used. (A) GCGAGATCGAACGGGGCTCTG. (B) GCTAAAT- CAAAGGGTGCCCGA.
the absence of the optional ORF. In the case of coxl, over 140 exonic nucleotides flanking the mono-orfic intron and 650 nucleotides in the intron, no sequence variation was detected when compared with the biorfic counterpart, except for the six substitutions already mentioned and an additional triplet ( T I T ) three nucle- otides upstream from the end of the intron (see Figure 1B'). These results are in agreement with previous data showing that mitochondrial sequence divergence among the P. ansaina strains is very low (CUMMINGS et al. 1990). They furthermore show that the differences between mono-orfic and biorfic introns containing genes are tightly clustered in close proximity to the optional sequences. The last six nucleotides of the nadl-
i4 upstream exon and the first five nucleotides of the coxl-i7 downstream exon, which are known to be in- volved in base pairings and required for intron splicing (5' side of the P1 and 3' side of the P10 stems, respec- tively), remain identical in the mono- and biorfic orga- nizations.
All the nucleotide differences observed between ex- ons flanking biorfic and mono-orfic introns are silent even though, in several cases, positions other than the third in the codons differ. The nine nucleotide substitu- tions in nadl-e4 led to the formation of seven modified codons. According to the codon usage in the mitochon- drial genes of P. ansm'na (CUMMINGS et al. 1990), even though unusual codons are found in both the exonic sequences (Figure 2, lines 4 and 5), the exonic sequence linked to the biorfic intron would be at least 1000 times less frequently used than the sequence linked to the mono-orfic (or to the equivalent sequence in the intron- less locus). In the case of coxl-i7, four codons are modi- fied when the optional ORF is absent, but there is no global significant difference in the probabilities of the exonic sequences (Figure 2, lines 10 and 11).
The d l - i 4 and coxZ-i7 equivalent introns in other Podospora species: P. comata and P. curuicolln: The or- ganization of nadl-i4 and coxl-i7 equivalent introns was examined in six other wild-type strains of Podospora belonging to two different species. nndl-i4 equivalent intron in P. comata was found again either biorfic (P.
co3) or mono-orfic (P. COT: P. co2), the upstream ORF (ORFl) being optional. In P. curuicolla, the nadl-i4 equivalent intron was either mono-orfic (P. cuV, P. c u m
or absent (P. CUL). coxl-i7 equivalent intron was found exclusively in the mono-orfic form in the three P. comnfn strains and no such sequence was present in the three
P. curuicolln strains. The absence of equivalent nndl-i4 (in P. cuL) and coxl-i7 (in P. curuicolln) mitochondrial introns, corresponds to a new (third) configuration for both genes (Figure 1, C and C ' ) . As the nndl-i4-orfl and the coxl-i7-@ were found optional in the species
P. anserina, the whole nadl-i4 and coxl-i7 introns are optional in the genus Podospora.
We have determined the nucleotide sequences of the exons flanking the nndl-i4 and coxl-i7 introns (or the corresponding regions in strains devoid of the intron) in the six additional Podospora strains. For nadl, the sequences are again of two types, with a high degree of similarity with the P. nnsaina equivalent exons flanking biorfic introns in the case of the P. eo3 (biorfic) and with the P. unsa'na equivalent exons flanking mono- orfic introns in the case of the P. C O T , P. c02, P. CUM,
P. CUV (mono-orfic) and P. C U L (intronless) strains. In
the case of the coxl-i7downstream exon, all the P. com- uta mono-orfic and P. curuicolla intronless strains pres- ent high degree of sequence similarity with the P. nnw-
n'na equivalent exon bordering the monwrfic introns. Alignment of the sequences with that of P. nnsm'nn strongly reinforces the previous correlation where six exonic consensus substitutions remain associated with the presence of the optional ORF (Figure 2, lines 3, 6,
7
and 12, 13).Whereas the mitochondrial genome of Podospora strains, within the same species, is >99% identical, ex- cepted the polymorphism for optional sequences, the exonic sequences adjacent to introns devoid of the o p
tional ORFs (or of the whole intron) present much more similarity with their counterparts bording mono- orfic intron in other species than with their counter- parts bording biorfic intron in the same species. Such comparisons were reported in Table 1 for the nndl-i4 locus (3'EX). The sequence comparison of the P. coT and P. co3 nadl-i4 mono- and biorfic introns revealed that they mainly differ by the presence of the nadl-i4-
orfl and associated exonic sequence: their catalytic core domain and O W 2 are very similar as is the case for
C. H. Sellem et
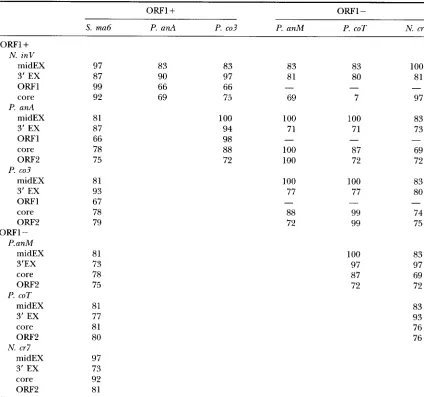
TABLE 1
Painvise comparisons in the d L i 4 locus
ow1
+
owl-
S. ma6 P. a d P. eo3 P. anM P. COT N. cr7
o w 1
+
N. inVmidEX 97 83 83 83 83 100
3' EX 87 90 97 81 80 81
o w 1 99 66 66
core 92 69 75 69 7 97
midEX 81 100 100 100 83
3' EX 87 94 71 71 73
ow1
66 98 -core 78 88 100 87 69
ow2
75 72 100 72 72- - -
P. anA
-
-
P. e03
midEX 81 100 100 83
3' EX 93 77 77 80
o w 1 67
core 74
ow2
79 7 2 99 75-
78 88
-
99
-
o w 1 -
P. anM
midEX 81 100 83
3'EX 73 97 97
core 78 87 69
ow2
75 72 72P. COT
midEX 81 83
3' EX 77 93
core 81 76
o w 2 80 76
midEX 97
3' EX 73
core 92
ow2
81N. cr7
The comparisons (percentage of identity, in nucleotides) concern the 28 last nucleotides of nadEe4 (3'EX), the nadEi4 intronic coding sequences ( O w l , OlZF2), and the catalytic core (from the 5' side of the P1 to the 3' side of the alternative P10 stems) of the intron between various Podospora strains belonging to the same or different species or between strains of different genera. The alignment of P. COT and P. anA nadCi4-orp
nucleotide sequences (675 nucleotides) revealed 72% identity with never more than 13 matched adjacent nucleotides. One hundred fifty nucleotides in the middle part of the upstream exon (midEX) were used as a mean of evaluation of the degree of divergence between strains of different genera. P. cuVand P. COT nadl- 24 intron presents the same organization and shows more than 99% similarity in sequence; P. cuVwas thus not included in the table.
other regions of the two mitochondrial genomes
(C. H. SELLEM and F. KOLL, unpublished results). (2)
The sequencing of a large part of the P. cuV nadl-i4
intron revealed >99% similarity with that of P. COT.
This high level of similarity is surprising since both spe- cies (P. curuicolla a n d P. comata) are believed to have diverged before the speciation P. ansm'na/P. comata. A
scenario giving an account for these unexpected levels of similarity will be proposed below (see Figure 5).
Mobility of nadl-i+olJI and coxl-i7+ in Podospora:
Heteroplasmons, resulting from the vegetative fusion between mycelia of Podospora strains differing by the
Mobile Intronic ORES i n Podospora 23.3
Probe 1
t
Probe 3
t
2.3 kb
-
-
Probe 2
-
1.9 kb-
1 . 1 kb4
Probe 4 FIGURE 4.--Southern blot analysis of of HndII-digested mi- tochondrial DNAs extracted from 10 homoplasmic strains ( I -
IO). Heterokaryons were constructed between parental strains bearing P. comntn nuclei and P. COT (T) and P. nnS (S) mitochondria. Each filter has been hybridized twice. Probes 1 and 2 allow the detection of the two optional ORFs. The exonic probe 1 lights a 2.2- or 1.3-kb band when nndl-i4-ofl is present together or without n d I - i 3 , respectively. A 1.5-kb band reflects the absence of both the nndI-i4-o?fl and nndl- i3 in P. cornntn (T) mitochondria. The transfer of nnd143 without nndl-i4-mfl (2.5-kb band expected) was never oh- served. The intronic probc 2 lights a 2.3-kb band according to the presence of coxl-i7-orj2. Probes 3 and 4 allow the itlenti- fication of polymorphic H n d I restriction sites. Probe 3 lights a 2.7- or 2.4-kb band according to the P. nnsm'nn or P. comrrtn origin of the nod595 locus. Probe 4 lights a 1.9- or 1.1-kb band according to the P. nnsm'nn o r P. comnfn origin of the coxl-il locus. Result? for 30 homoplasmic strains are summa- rized in Table 2 (see *).
strains harboring a given marker ( i . ~ . , polymorphic re- striction site) from the donor strain was used as a mea- sure of the transmission rate (Figure 4). nadl-i4-urfl
and coxl-i7-0@ were found in 100% of the homoplas- mic strains whereas the polymorphic genetic markers from the donor were transmitted with an efficiency of
<50% differing from one experiment to another (Ta- ble 2). Intronic ORFs thus appeared highly invasive in agreement with the behavior of mobile elements propa- gating via the homing mechanism described for group
I introns (DUJON 1989). Moreover, the sequences of the new exon-intron junctions indicated that the trans- mission of the optional ORFs was, in all cases, accompa- nied by coconversion in the adjacent exon, bringing the exonic sequence characteristic of the optional ORF-
containing strain (Figure 2, lines 4 and 10). Bidirec- tional coconversion was expected if the double strand break repair process, involved in group I intron hom-
ing, also applies to ORF mobility. Monitoring of the conversion inside the intron was not possible when the two parental strains contained identical intronic se- quences (core and constitutive ORF). In the case of the H P ~ ro7'/nnS progeny, where the parental strains contained vel? divergent nndl-i4 sequences (Tablel),
the entire intron sequence (ORFl, core, ORF2) was
found to be that o f the donor strain (not shown). T h e nudl-i4 and coxl-i7introns in other Pyrenomycete genera: ,VPuros/)ortr nnd .Yordm-iri: n d l - i 4 and roxl-i7
equivalent introns, inserted at iclentical genomic sites
(a highly consenred domain of the exonic amino acid sequence), have been described in mitochondrial DNA of Neurospora. Their structure is similar to that found in Podospora (see Figure 1). nndl-i4, nndl-i4-orfl and
nndl-i4-orJ2 are optional in Neurospora as deduced from the three different organizations found in this genus (MOTA and COI.I.INS 1988; H x ~ s l ; . PI rrl. 1990). T h e nndI-i4 equivalent intron is m o n w r f i c in some strains, containing either an equivalent ORFl ( N . intw-
medin, strain Vnrhud) or an equivalent ORF2 ( N . rrnssn, strain 74A), or intronless in others ( N . rmssn, strain
Abbott 12). Interestingly, the nndl-i4 equivalent in N. intme dia was probably derived from a biorfic intron; it contains a full length and most likely functional OW1
equivalent but a truncated OW2 in which 47 codons of the N. rrnssn equivalent ORF were retained as well
as the hypothetical alternate .?'-splice site that would ensure the translation of the free standing ORF before it was lost (Figure 1D). We have also undertaken the sequencing of the nndI-i4 equivalent intron in a related fungus, S. mnrros/mn, we found to present a biorfic orga- nization with an exceptionally long and fragmentary
OW2 (C. H. SE:I.I.EM, unpublished results). T h e coxl-i7
intron is optional in Neurospora. In the only strain where it has been described ( N . c7'n~ssn Adiopodoump), it presents the same mono-orfic organization as in Po- dospora strains (FIELD et d . 1989). Alignment of the exonic sequences, adjacent to optional ORFs o r in the homologous location in genes devoid of the intron, revealed that the correlation previously observed in
Po-
dospora is maintained throughout all the Pyrenomycete strains examined (Figure 2). Over a total of 29 strains examined, belonging to six species in three different genera, five consensus substituted positions in the nndl- i4 equivalent upstream exon and four in the coxl-i7
equivalent downstream exon are perfectly correlated with the presence of the corresponding optional ORF.
The exonic sequences located upstream of the nndI-i4-
of1 in various species and genera ( N . inl< P. nnA, P. co3, S. mar;) present much more similarity one to an-
other than with the corresponding exonic sequences in strains of the same species devoid of ORFl ( N . 0-7, P. nnM,
I?
ro7) that in turn appear to be more closely related to one another (Table 1).TABLE 2
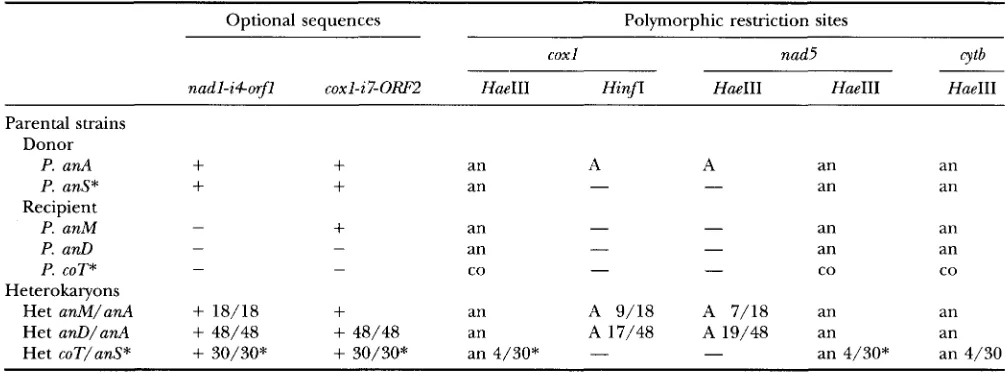
Mobility of intronic ORFs in heterokaryons
Optional sequences Polymorphic restriction sites ~~ ~~~
cox1 nad5 CYtb
nadl-i4orfl coxl-i7-ORFZ HueIII Hinfz HueIII HaeIII HaeIII
Parental strains Donor
P. anA
+
P. anS*
+
Recipient
P. anM -
+
+
+
an an
an
A A
- -
an an
an an
an P. a d
P. COT* Heterokaryons
- - an
-
- an an- - co - - co co
Het anM/anA
+
18/18+
an A 9/18 A 7/18 an anHet anD/anA
+
48/48+
48/48 an A 17/48 A 19/48 an anHet coT/anS*
+
30/30*+
30/30* an 4/30* - - an 4/30* an 4/30Homoplasmic strains were issued from heterokaryons raised between (recipient/donor) isogenic strains bearing P. anA and P. anM (Het anM/anA), P. anA and P. anD (Het anD/anA) or P. COT and P. anS (Het coT/anS) mitochondria. The donor and recipient parental strains exhibit the A (or A') and the B (or B') organization, respectively (see Figure 1). The presence (+)
of nadl-i+orfl and coxl-i7-orf2 was visualized on Southern blot of mitochondrial DNA digested with HaeIII as described in legend
of Figure 4. The five independent mitochondrial genetic markers monitored in the progenies are polymorphic restriction sites: Hue111 digestions allowed the detection of the P. ansm'na (an) or P. cornata (co) origin of the 60x1-il, nad5e5 (see Figure 4) and cytkil loci. In the same way, Hue111 and Hinfz restriction sites specific of the strain P. anA (A) in the nad5il and coxl-il introns, respectively, were also monitored in the progenies. The number of homoplasmic strains (over the total examined) harboring the various markers from the donor strain. is indicated. The asterisks point out the characteristics of the parental strains and heterokaryon progeny (Het coT/anS) presented in Figure 4.
mycete species other than those already mentioned har- bor a nadl-i4 equivalent intron. Among filamentous fungi, Aspergdlus nidulans (BROWN et al. 1983) and Ascc- bolus immersus (C. H. SELLEM, unpublished results) are intronless (Figure 1C). A pairwise comparison of the sequences equivalent to the P. ansm'na nadl-e4 30 last nucleotides was undertaken for distantly related spe- cies. Even based on a very short sequence comparison, the resulting unrooted phylogenetic tree (inset in Fig- ure
2 )
reflects rather well the evolution of the various kingdoms (bacteria, fungi, plants, animals) with only few cases of incorrect branching. The division of the fungal kingdom in two distant clusters, one of which included all the nadl-i4-ofl-containing strains and branching earlier in the tree, will be discussed belowas the result of horizontal transfers of nadl-i4-orfl and neighboring exonic sequences.
The correlation between the presence of the optional sequence and a consensus nucleotide sequence in the adjacent exon led us to evaluate the degree of similarity in optional against nonoptional ORFs in various wild- type strains of different genera. This has been done only for nadl-i4 since no biorfic coxl-i7 equivalent was found elsewhere than in P. ansm'na. The coxl-i7equiva- lent introns found in sequence banks and not already mentioned belong to A. nidulans (WARING et al. 1984)
and, surprisingly, to the liverwort Murchantia polymorphs (OHTA et al. 1993); in both cases they show a mono- orfic organization (Figure 1C'). Three of the four sub-
stitutions associated with the absence of the down- stream ORF remain conserved. nadl-i4 sequences are available for one strain of P. ansm'na and two species of Neurospora (MOTA and COLLINS 1988; HAWSE et al.
1990). They were compared with mono-orfic or biorfic
nadl-i4 intron sequences we have obtained for various Podospora strains from different species and for
S.
mac- rospura. Table 1 shows that, among the various strains examined, core and OW2 evolved in a very similar way. The level of similarity observed within a species is high while it is much lower between species and genera. This is suggestive of the evolution of the mono-orfic intron from a common ancestor to the three genera. On the contrary, the n a d l - i 4 - ~ r f l sequences of P. ansm'na andP. comata are very similar in otherwise divergent introns (core and ORF2) as are those of N. intermedia and
S.
macrospora suggesting, in those cases, horizontal transfer events had occurred.
The 17-bp sequence, located downstream from n a d l - i h r f l and upstream from the core structure in the P. ansm'na biorfic intron, was identified as an exact replica of the 3' end of the N.crassa nadl-i4 equivalent up- stream exon (CUMMINGS et al. 1988). We found the same sequence in the 3' end of P. ansm'na nadl-e4
Mobile Intronic Oms in Podospora 785
5' side of the P1 stem of the RNA structure, then consti- tute a direct repeated sequence flanking the optional
OW. A direct repeat organization was also found in the coxl-i7 biorfic intron: a &bp sequence overlapping the intronexon junction is also present at the core-
ORF2 junction. These repeats constitute the proximal alternative (upstream repeat) and the distal (down- stream repeat) 3"splice sites determined by the 3' side of the alternative and constitutive P9-0 and P10 stems flanking the last G of the intron (SELLEM and BELCOUR
1994). A comparable situation has been described for the Neurospora nadl-i4 (MOTA and COLLINS 1988), but direct repeats flanking both optional ORFs are not pre- cise elements of pairing in the RNA secondary struc- ture, as in Podospora. The significance of such short repeats (5-8 bp) flanking the optional ORF and of sequences
(17
bp) found both in the exon bording the mono-orfic and in the biorfic intron is not clearly understood, but they have been proposed to be in- volved in the acquisition or loss of the optional se- quences (MOTA and COLLINS 1988; VADER et al. 1994).DISCUSSION
The mitochondrial chromosome in wild-type strains of the filamentous Ascomycete P. a n s m k presents a previously unreported feature: the presence, within a species, of optional intronic ORFs. Two introns refered as nadl-i4 and coxl-i7 may present several organizations depending on the natural isolates examined: either bi- orfic (containing two ORFs) or mono-orfic (containing only one). The organization of these two introns in
P. ansm'na offered the opportunity to investigate the mobility of the optional ORFs as independent genetic elements. Furthermore, the systematic clustered nucleotidic differences we found between exons flank- ing mono- and biorfic introns led us to question the evolutionary origin of these ORFs and the mechanisms involved in their transfer.
Arrival of intronic Oms in the nadl44 and coxl-i7 introns of Podospora: The large number of clustered substitutions we found in P. anserina mitochondrial ex- ons adjacent to the optional ORFs in biorfic nadl-i4 and coxl-i7introns, relative to their mono-orfic counter- parts, is clearly unusual since much more than 99% conservation of sequence has been found in the overall genome of the wild-type strains belonging to this species
( CUMMINGS et al. 1990). The correlation established be- tween the presence of the optional ORFs and the nucleotidic sequence in the adjacent exon is strong enough to be extended to all other Pyrenomycete spe- cies we have examined. What does this correlation mean? The presence of an optional sequence in a spe- cies raises the question of its loss or gain in the course of evolution. When the sequence is an intron, optional- ity is generally explained by a steady state between loss and gain (DUJON 1989). How to loose and gain group
I introns seems now to have been elucidated: loss pro- ceeds via reverse transcription of mature RNAs (GAR-
GOURI et al. 1983; LEVRA-JUILLET et al. 1989; SAINSARD- CHANET et al. 1993) and gain (homing), via double- strand breaks and conversion of the intronless allele using the intron containing gene as a template (DUJON
1989; LAMBOWITZ and BELFORT 1993).
The known mechanism of loss could not be invoked for the nadl-i4 and coxl-i7optional ORFs, since no splic- ing event generating RNA molecules cleanly devoid of the optional sequences is expected or detected.
Any-
how, all the spliced RNA molecules issued from nadl and coxl transcripts containing the optional ORF exhib ited, as expected, the exon characteristic of the biorfic introncontaining strains (SELLEM and BELCOUR 1994). A loop out at the DNA level, between direct repeats flanking the optional ORFs, could exceptionally pro- mote their deletion as it has probably been the case for part of the nadl-i4-orf2 in a close ancestor of N. interme- diu (MOTA and COLLINS 1988). According to the pres- ence of similar direct repeats, complete excision of the two optional ORFs would also be possible in Podospora nadl-i4 and coxl-i7 intron. However, such an ORF loss would not be accompanied by concerted modifications in the region of excision. Loss of the intronic OW,
unless invoking a new mechanism with coconversion, could then not be invoked in a steady-state scenario where optional sequences are periodically lost and gained. Such a mechanism of loss, first invoked by HEN-
SGENS and coworkers (1983) for an intron, implying cut and repair using a foreign OW-less genome as tem- plate, cannot however be completely excluded. In con- trast, gain of intronic ORFs could be explained by a mechanism equivalent to that proposed for group I
intron homing that perfectly accounts for the modifica- tions encountered in the sequence flanking the inser- tion region. Moreover, in the case of nadl-i4 and coxl- i7optional ORFs, homing (gain) is consistent with their observed invasiveness.
Mobility of intronic ORFs inside the genus Podospora:
In the progeny of heteroplasmons constructed between biorfic and mono-ofic intron-containing strains, we have shown that the nadl-i4-orfl and the coxl-i7-orJ2 appeared highly invasive. Indeed, all the homoplas- mons (100%) tested had acquired the optional ORF
ent strains (Table 1 ) could be responsible for an inevita- ble coconversion of the intron initiated by the transfer of the ORF as the mobile element. The alternative and less likely interpretation would be the transfer of the intronic ORF together with the whole intron, proceed- ing via an intron replacement; the biorfic intron taking the place of the mono-orfic one. These experiments nevertheless constitute the first examples of driven O W transfer from an OW-containing intron to the cognate OW-less allele, probably miming the horizontal events that have recently taken place in the nadl-i4 and coxl- i
7
introns of Podospora.The acquisition of nadl-i4-0@
and
coxI-i7-0@ is re- cent in Podospora: The pattern of presence/absence of optional ORFs suggests that the steady state has not been reached and that they are currently invading the population. While for an intron steady state would mean periodical losses and gains, in the case of mobile intronic ORFs, steady state would be all the introns being invaded. The acquisition of nadl-i4-orfl and coxl- i7-orf2 by Podospora may be relatively recent, so that insufficient time has elapsed to permit evolution of the overall genome in two strains of the same species dif- fering by the presence of the optional ORF. The pres- ence of a 17-bp sequence, both in the 3' noncoding region of the optional nadl-i4-orfl in biorfic intron (P. anS and P. eo?) and in the 3' end of the nadl-e4 exon adjacent to mono-orfic intron, is surprising. This could suggest that the optional ORF has integrated into the exonic sequence adjacent to mono-orfic intron, bring- ing at the same time a sequence equivalent (but suffi- ciently modified) to the one displaced by the insertion. An analogous situation was already described (PAQUINet al. 1994). In the case of P. ansm'nu, the identity in the 17-bp sequence, present in mono- and biorfic loci, could be an additional argument for the recent acquisi- tion. Data given in Table 1 agree with the current acqui- sition of nadl-i4-orfl and associated exonic modifica- tions by the Podospora nadl-i4 mono-orfic intron (core and ORF2). P. ansprina ( P . a n A ) and P. comata (P. eo?)
nadl-i4-orfl present much more similarity one with an- other than the overall intron does. This strong similarity at both amino acid and nucleotide levels implies not only similar functional constraints but also a recent common origin.
Origin of the optional ORFs: The acquisition of ad- ditional intronic ORFs by Podospora nadl-i4 and coxl-
i7 introns, together with modifications in the flanking exons, are most probably the result of recent horizontal transfers via homing-like events from other fungal n a d l
and coxl mitochondrial genes. Indeed, searching in se- quence banks has revealed that the exonic sequence adjacent to the studied optional ORFs present no sig- nificant similarity with any other gene than the mito- chondrial n a d l and coxl. In addition, the codon usage of the two ORFs appears to conform to that used in fungal mitochondria, whereas in the case of n a d l , that
of the adjacent exon is especially abnormal. This could mean that the two optional ORFs were present in mito- chondrial genomes for a sufficiently long time to have adapted their codon usage, whereas the associated exo- nic sequences are probably assigned to different con- straints preventing such adaptation. The especially highly improbable codon usage in exons flanking bior- fic nadl-i4 intron led us to search, in sequence banks, for an hypothetical n a d l foreign donor gene. A phylo- genetic tree was constructed from painvise comparisons of equivalent nadl-e4 30 last nucleotides among various unrelated species including those containing mono-or- fic and biorfic nadl-i4 introns. Although based on a very short sequence, this tree evidences the horizontal tranfer of nadl-i4-orfl in various fungal strains (Figure
2, inset) by the independent clustering of all the O w l - containing strains. The branch point could give an idea of the possible foreign donor organism that appears to be more distantly related to Podospora (Pyrenomycete) than Podospora is to Aspergillus (Plectomycete) .
Horizontal transmission of introns between different organisms, although never observed in a laboratory, has previously been invoked to explain the presence of related introns in otherwise more divergent genes
(LANG 1984; WARINC et al. 1984; TRINCKL and WOLF 1986) or in different locations (MICHEI, and CUMMINCS
1985; COLLEAUX et al. 1990). Such lateral transmission events were also proposed for the ORFs containing the
GIY-YIG motif (found in Podospora and Neurospora mitochondrial group I introns), as a genetic exchange between a close ancestor of the Neurospora-Podospora group and the bacteriophage T 4 genome (MICHEL and
DUJON 1986).
Evolution of the d l - i 4 intron: In Figure 5 we have schematically drawn, from data of different sources tak- ing into account both biological (see MATERIALS AND
METHODS) and sequence data, the lineage of the various strains studied in this work. Clearly, numerous events of gain and loss of optional sequences are needed to explain the current status of the various nadl-i4 equiva- lent introns examined. We have indicated the most probable events that also take into account the se- quence similarity found over the three domains of the intron between the strains, species and genera of the F'yrenomycetes examined (Table 1). When very similar sequences were found in divergent contexts, we have invoked horizontal tranfer events: between P. a n A and
P.co3 and between N. intermedia and S. macrospora for
nadl-i4-orfl and nearby exonic sequences, and between
P.comata and P.curuicolla for the mono-orfic nadl-i4 in- tron. The rather poor degree of sequence similarity found between Podospora and Neurospora nadl-i4-orfl
Mobile Intronic ORFs in Podospora
i : n a d 1 - i 4
core
+
0 2mono-orfic intron
787
Sordaria Neurospora
&
PodosporaI
iN.interrnedia ~ o2
,
N. c ri.
~ ,P.an;erina co,4P. c o ~ t a,
,P. cuy'co//;S. ma N inV N.crA N.cr7 P.anM P.anA P.co3 P.coT P.cuM P.cuL
01 0 2 0 1 0 2 i- 0 2 01 0 2 01 0 2 0 2 0 2 i-
FIGURE 5.-Proposed nadl-i4 evolution in three Pyrenomycetes genera. The dendogramme is a schematic representation reflecting the evolution of the nadl-i4 intron in the various genera where we have indicated the most probable events of gain
(e) or loss ( \ ) of the optional sequences [nadl-ikwfl ( O l ) , nadl-i4-orf2 ( 0 2 ) and the whole n u d l 4 mono-orlic intron (i, ie., core and ORF2)] consistent with the current organization of the locus in the various strains examined (bottom). The evolutionary relationship between the three Pyrenomycetes, Podospora, Sordaria and Neurospora, is based on the sequence comparison of ribosomal RNA (WILMOTTE et al. 1993). The resulting tree is in agreement with the conservation of their chromosomal genetic map (LEBLON et al. 1987), and with nuclear (B. GAGNY and P. SILAR, personnal communication) and mitochondrial sequence comparisons in regions outside of the nadl-i4 locus (C. H. SELLEM, unpublished results). The phylogeny between the various Podospora strains is based on the conservation of mitochondrial chromosome maps (BOCKELMANN and ESSER 1986) and on mitochondrial sequence comparisons (F. KOLL, unpublished results).
-01 i
the overall mitochondrial genome (-80% similarity). The transfer of nadl-i4 mono-orfic intron from P. curui- colla to P. comata implies a prior loss of the intron in P. comata and explains why the degree of similarity of the
nadl-i4 introns (core and ORF2) is low between two closely related species (P. ansmha and P. comata) and is high between two more distant ones (P. comata and
P. curuicolla). All the transfer events proposed on the basis of sequence similarities (Table 1 ) now agree with the presence of an ancestral nadl-i4 mono-orfic intron that predates in the three F'yrenomycete genera. Figure
5 gives a parsimonious scenario that also takes into ac- count the loss of intronic sequences such as nadl-i4-
.If2
inN.
inV (loop out of the O W ) and occasional loss of the mono-orfic intron (P. comata, P. cuL andN.
crA).
It could be predicted that loss of biorfic introns may also occur: according to the known mechanism of loss, this would result in an intronless gene bearing an up- stream exon of the biorfic type. No such strains have yet been found among the 23 strains examined in Po-
dospora, neither in the case of nadl-i4 nor coxl-i7, sug- gesting again that the acquisition of the two optional
ORFs and associated exonic sequences is very recent. Our results greatly support the idea that group I in- tronic ORFs are mobile by themselves and have found a safe location in preexisting introns. The case of biorfic introns is interesting since the acquisition of the op- tional ORFs is recent and provides evidence that the mechanism of their mobility involves modifications of
adjacent exons, thus playing a role in the modular evo- lution of the mitochondrial genomes.
We are grateful to E. DUFOUR for technical help in the preparation of the figures and to S. HADDAD for sequencing assistance. This work was supported by grants from the GREG (no. 71/94) and from the Fondation pour la Recherche MPdicale.
LITERATURE CITED
BECHHOFER, D. H., K. K. HUE and D. A. SHUB, 1994 An intron in the thymidylate synthase gene of Bacillus bacteriophage p22:
evidence for independent evolution of a gene, its group I intron, and the intron open reading frame. Proc. Natl. Acad. Sci. USA
91: 11669-11673.
BELCOUR, L., and 0. BEGEL, 1977 Mitochondrial genes in Po- dospora anserina: recombination and linkage. Mol. Gen. Genet. 153: 11-21.
BELL-PEDERSEN, D., S. QUIRK, J. CLYMAN and M. BELFORT, 1990 In- tron mobility in phage T4 is dependent upon a distinctive class of endonucleases and independent of DNA sequences encoding the intron core: mechanistic and evolutionary implications. Nu- cleic Acids Res. 18: 3763-3770.
BINISZKIEWICZ, D., E. CESNAVICIENE and D. A. SHUB, 1994 Self-splic- ing group I intron in cyanobacterial initiator methionine tRNA evidence for lateral transfer of introns in bacteria. EMBO J. 13:
BOCKELMANN, B., and K. ESSER, 1986 Plasmids of mitochondrial origin in senescent mycelia of Podospmu curvicollu. Curr. Genet. BROWN, T. A,, DAVIES, R. W., J. A. RAY, R. B. WNNG and C. SCAZZOC
CHIO, 1983 The mitochondrial genome of Aspergillus nidulans
contains reading frames homologous to the human URFs I and 4. EMBO J. 2: 427-435.
BURGER, G., and S. WERNER, 1985 The mitochondrial URFI gene in Neurospora crussu has an intron that contains a novel type of
URF. Mol. Biol. 186: 1231-242. 4629-4635.