(hpvright 0 1997 by the Genetics Society of America
The Maize Transposable Element Ac Induces Recombination Between the Donor
Site and
an
Homologous Ectopic Sequence
Gil Shalev
and
Avrzlham
A.
Levy
Department of Plant Genetics, The Weizmann Institute of Science, Rehovot 76100, Israel
Manuscript received December 9, 1996 Accepted for publication March 28, 1997
ABSTRACT
The prominent repair mechanism of DNA double-strand breaks formed upon excision of the maize
Ac transposable element is via nonhomologous end joining. In this work we have studied the role of homologous recombination as an additional repair pathway. To this end, we developed an assay whereby &Glucuronidase (GUS) activity is restored upon recombination between two homologous ectopic (nonal- lelic) sequences in transgenic tobacco plants. One of the recombination partners carried a deletion at the 5’ end of GUS and an Ac or a Ds element inserted at the deletion site. The other partner carried an intact 5’ end of the GUS open reading frame and had a deletion at the 3‘ end of the gene. Based on GUS reactivation data, we found that the excision of Ac induced recombination between ectopic sequences by at least two orders of magnitude. Recombination events, visualized by blue staining, were detected in seedlings, in pollen and in protoplasts. DNA fragments corresponding to recombination events were recovered exclusively in crosses with Ac-carrying plants, providing physical evidence for Ac-
induced ectopic recombination. The occurrence of ectopic recombination following double-strand breaks is a potentially important factor in plant genome evolution.
T
HE transposable element Activator ( A c ) of maize and the related nonautonomous Dissociation(Ds)
elements belong to the same class of elements as the En/Spm of maize, P of Drosophila, Tcl of nematodes, TnlO and Tn7 of Escherichia coli, all sharing a conserva- tive “cut-and-paste” mechanism of transposition (see reviews in SAEDLER and GIERL 1996). According to this model, a double-strand break (DSB) is created at the donor site upon the element excision. Healing of the broken donor can occur through a number of different pathways. In E. coli, where DSB repair via nonhomolo- gous end-joining is not efficient, broken DNA at Tn7 or TnlO donor sites is repaired via homologous recom- bination (HR) (HAGEMANN and CRAIG 1993; EICHEN- BAUM and LIVNEH 1995). Similarly, HR is involved in the donor site repair following P (ENGELS et al. 1990) and Tcl (PLASTERK 1991) excision. Based on genetic and molecular characterization of Pand Tcl donor site repair, a model was proposed that includes the follow- ing successive events: DSB formation upon element ex- cision, gap enlargement via exonucleolytic degradation exposing 3’ single-strand regions, and invasion of a se- quence homologous to the donor site to be used as a template for gap repair (ENGELS et al. 1990; PLASTERK 1991; NASSIF et al. 1994). The template for donor site repair can be supplied by either the sister chromatid, the allelic sequence on the homologue, a nonallelic (ectopic) sequence (ENGELS et al. 1990) or an oligonu-Corrrsponding author: Avrahanl A. Levy, Department of Plant Genet- ics, The Weizmann Institute of Science, Rehovot 76100, Israel. E-mail: [email protected]
Genetics 146 1143-1151 (July, 1997)
cleotide (BANGA and BOYD 1992). Such a gap repair mechanism can account for the apparent lack of corre- lation between excision and reinsertion and for the effect of an allelic sequence on the reversion frequency at the donor site.
Although in plants HR is not efficient, there are indi- cations that it is induced upon DSB formation: (1) Irra- diation was found to enhance the rate of HR (TOVAR and LICHTENSTEIN 1992; PUCHTA et al. 1995). (2) Geno- mic DSB, catalyzed by the yeast I-SceI endonuclease, induces HR between a T-DNA template and a tobacco genomic sequence (PUCHTA et al. 1996). (3) An Ac/
Ds
insertion was shown to increase crossover between markers flanking the insertion site (MCCLINTOCK 1953; GREENBLATT 1981). (4) A c excision from the maize P locus was found to induce recombination between large direct repeats flanking A c donor site (ATHMA and PE-TERSON 1991). Similarly, the maize Ivfutatorelement was shown to increase recombination between tandem re- peats at the K n l - 0 locus (LOUT et al. 1992).
joining (or illegitimate recombination), following exo- nuclease and DNA-polymerase activities (COEN et al. 1989; SAEDLER and GIERL 1996; S C O ~ et al. 1996).
To better assess the role of HR in Ac donor site repair, we have developed a nonselective assay in transgenic tobacco that allows us to monitor HR in the region flanking an Ac or a nonmobile
Ds
element through blue sectors, which reflect the repair of a defectivep-
Glucuronidase (GUS) gene. We found that Ac excision induces HR between genomic sequences located at ec- topic (nonallelic) sites by at least two orders of magni- tude. In agreement with these findings, recombinant DNA molecules were detected only in Ac- but not inDS
carrying plants. These data show that in addition to the nonhomologous end-joining repair pathway, HR is used for donor site repair following Ac excision. To our knowledge, this is the first indication of ectopic HR in plants.
MATERIALS AND METHODS
Plasmid construction: Digestions with restriction enzymes, ligations and plasmid preparations were performed according to standard protocols and manufacturer's instructions (SAM-
BROOK et al. 1989). Plasmid pJD330, kindly provided by V.
WALBOT, was used as a positive control for assaying GUS activ- ity. This plasmid is a Bluescript derivative carrying the GUS reporter gene driven by the cauliflower mosaic virus 35s pro- moter fused to the tobacco mosaic virus translational en- hancer R (GALLIE et al. 1987). The transcription termination fragment of pJD330 is derived from the Nopaline synthase (nos) gene of Agrobactm&m tumefaciens. To test whether Ac
induces HR, we built a series of constructs, derived from pJD330, schematically described in Figure 1. Recombination partners were constructed, each with a different deletion in the GUS gene. One partner (3'AGUS), carried a 700 base pair (bp) (SspI-EcoRI) deletion in the 3' region of GUS, span- ning -500 bp within the GUS gene, and the nos transcription termination fragment. The 3'AGUS (laboratory number pGS001) plasmid was prepared by digestion with EcoRI, blunt- ing with Klenow, followed by partial Ssp1 digestion and self- ligation of the 4.5-kilobase (kb) fragment. The second partner consisted of a deletion in the 5' end of GUS with either Ac or Ds inserted at the deletion site. For this purpose pJD330 was linearized at the SalI site located between R and the GUS gene. The deletion was created by treating with BaB1, the ends were blunted with Klenow and a 15-bp KpnI-SpeI adapter was synthesized and ligated to the treated vector fragment. This yielded the 5'AGUS (pGSOO3) plasmid. The BaZ31 dele- tion in pGSOO3 consisted of a 12-bp deletion, which included the ATG initiation codon of GUS. This plasmid as well as pGSOOl were tested for GUS activity in transient assays and found to be GUS inactive (data not shown). The KpnI-SpeI adaptor was also ligated into the blunted SalI site of pJD330. The resultant plasmid pGSOO2 was found to be GUS active in transient assays (data not shown). Two plasmids, derivatives of pGS002, were constructed to test Ac ability to excise and
to confirm Ds lack of mobility. The Ac element used in this study is derived from the wx-m7 allele (CHOMET et al. 1987). It was isolated together with some Waxy flanking nucleotides from plasmid pJJ4411, generously given by H. DOONER (KEL LER et al. 1993), and subcloned into pBluescript I1 Ks (Blues- cript) giving rise to plasmid pNAOO2. An Ac-containing KpnI- SpeI fragment was isolated from pNAOO2 and subcloned into
the same sites in the adaptor of pGSOO2 plasmid giving rise to GUS:Ac (pGS004). A Ds-containing plasmid GUS:Ds (pGSOO5) was derived from GUS:Ac by filling in the unique A d 1 site, located in the coding region of Ac. In these plasmids GUS activity can be restored upon element excision. Ac- or Ds-containing KpnI-SpeI fragments were isolated from pNAOO2, or from pCSOO5, and subcloned into the same sites in the adaptor of pGSOO3 giving rise to the two recombination partners, 5'A GUS:Ac (pGSOO8) and 5'AGUS:Ds (pGSOO9) , re- spectively. In these plasmids GUS activity cannot be restored upon element excision.
All plasmids described above were subcloned into binary vectors to produce transgenic plants. The HindIII-BglrI frag- ment of pJD330 was subcloned into the same sites of the pGA492 binary vector ( A N 1987) giving rise to pGSOO6. The HindIII-PVuII fragment containing the 3'AGUS of pGSOOl was subcloned into the HindIII-filled-in CluI sites of the same binary vector giving rise to pCSOO7. The 7-kb PstI-BglII GUS:Ac containing fragment of pGS004 was subcloned into the HpaI- BamHI sites of the same pGA492 binary vector giving rise to pGS010. The 7-kb PstI-BgZII fragment of 5'AGUS:Ac (pGSOO8) was subcloned into the PstI-BamHI sites of the binary vector pCGN1548 (MCBRIDE and SUMMERFELT 1990) giving rise to pGSOl2. Similarly, the blunted 7-kb EcoRI-XbuI fragment of 5'AGUS:Ac (pGS008) was subcloned into the blunted PstI- BglrI sites of the pCPTV-HPT binary vector (BECKER et al.
1992) giving rise to pGS015. The 7-kb PstI-BglII fragments of GUS:Ds (pGS005) and 5'AGUS:Ds (pGSOO9) were ligated with the 14.2-kb PstI-BglII vector-containing fragment of pGS012 giving rise to pGSOll and pGSO13, respectively.
Tobacco transformation: The binary vectors mentioned above were introduced by electroporation into the Agrobacter- ium tumefuciens LB4404 strain (HOEKEMA et al. 1983). The transformed Agrobacterium strains were used to infect leaf discs of Nicotiana tabacum and regenerate transgenic plants as described by Horsch et al. (1985). All transgenic plants carried transformation markers that confer resistance to kanamycin (100 mg/liter) or hygromycin (18 units/ml). Analyses were carried out in progenies of crosses whose parents were hemi- zygous for one copy of the T-DNA. The number of T-DNA copies was determined on the basis of kanamycin or hygro- mycin resistance frequency in T2-selfed seedlings and by Southern blot analysis (data not shown).
GUS assays: GUS activity in transgenic plants was deter- mined according to the fluorimetric or histochemical proce- dures
JEFFERSON
et al. 1987). Tissues of transgenic plants were stained with the X-Gluc (5-bromo-chloro-3-indolyl-~-D- glucoronide from Molecular Probes) substrate and subse- quently cleared as described (BEECKMAN and ENCLER 1994).Pollen staining was performed with a solution consisting of 250 mM Mannitol, 100 mM (pH 7) phosphate buffer and 1% DMSO. Pollen cells were stained at the day of anthesis for 1
hr at 37" under sterile conditions.
Microcalli staining was done using mesophyll protoplasts isolated from leaves of double hemizygote F1 plants of the 3'AGUS x 5'AGUS:Ac and 3'AGUS X 5'AGUS:Ds combina- tions as described (FRIDLENDER et al. 1996). The protoplasts were placed in tissue-culture plates containing 10 ml of the VKM X 2 proliferating medium for 6 days in a tissue-culture chamber, under low light conditions ( A W a n d GALUN 1985). On the sixth day, when microcalli consisted of four to 16 cells, GUS activity was assayed by X-Gluc staining and clearing was done as described for seedlings. A total of 500,000 micro- calli were screened, and recombinant blue cells were counted under an inverted Axiovert 35 Zeiss microscope.
Ac-Induced Homologous Recombination 1145
92“ for 1 min, annealing at 58” for 55 sec and elongation for 50 sec at 72” for 30 cycles. To isolate recombinant and excision products, two rounds of PCR amplification with nested primer pairs were done using primers a, b, c and d shown in Figure 1. In the first round, primers a (5”ATGAGCGGAGAATTA- AGGGAG3‘) and b (5’-GTCGGGTCGAGTTTACGCG3’) were used. As illustrated in Figure 1, this step could not am- plify a product from pGS007, as primer b is in the deleted region of GUS in pGS007 and primer a is in the right border region unique to the pCPTV binary vector. This step could not amplify a product from pGSOl3 or pGS015 if excision had not occurred. A 5-p1 sample of the first round was used
as a template in the second round of amplification carried out with primers c (5’-GCCGTTTTACGTTTGGAACTG3’) and d (5’-CGCGATCCAGACTGAATG3’). Southern blots were per- formed with MSI membranes according to the manufacturer’s instructions. Two different probes, prl and pr2, were used (see Figure 1). One probe was prepared by random priming (FEINBERG and VOGELSTEIN 1983) of an internal DNA frag- ment from the 35s promoter (see probe prl in Figure 1 ) . For a second probe, a synthetic oligonucleotide (pr2, 5’-GAG GCTGTACGTGGGGCGCGTTGG3‘), corresponding to the
Waxy sequence flanking Ac insertion in the original maize allele, was end labeled (see probe pr2 in Figure 1) as de- scribed by SAMBROOK et al. (1989). Excision products were amplified using primer pr2 (described above) and primer w (5‘-AGTGGATCCCCCGCGCGGCG3‘) flanking Ac 3’ end.
Cloning PCR products: The PCR products corresponding to putative recombination or excision eventS were cloned within the pGEM-T vector system (Promega) and sequenced using the T7 or SP6 primers.
RESULTS
An assay €or ectopic recombination: To assay Ac-in- duced HR, transgenic plants were made with the con- structs described in MATERIALS AND METHODS. A recom- bination assay was developed, based on reactivation of the GUS gene using constructs described in Figure 1. Crosses were made between T1 plants transformed with the 3‘AGUSand TI plants transformed with 5’AGUS:Ac or 5’AGUS:Ds. Ac is expected to generate a DSB near the 5’ deletion site upon excision, while Ds is not ex- pected to excise. This Ac-induction of DSB is expected to trigger HR. GUS reactivation could occur if the 12- bp deletion in the 5‘AGUSis repaired by a gene conver- sion-like mechanism using the intact 5’ region of the 3’AGUS partner as a template. Alternatively, one cross-
over product containing the NPTII region, the 35s pro- moter, the intact ATG of the 3’AGUS construct and the intact 3’ region of GUS from the 5‘AGUS could also produce an active GUS enzyme. The reciprocal cross- over product containing the right border from the T- DNA in 5’AGUS, the 35s promoter and the CAT gene in 3‘AGUS would not produce an active enzyme. Eight different genetic combinations, involving parents de- rived from independent transformation events, were analyzed for the 3‘AGUS X 5‘AGUS:Ac cross (pGSOO7 X pGSO12 or pGS007 X pGS015), and nine combina- tions were analyzed for the 3’A GUS X 5’AGUS:Ds cross (pGSOO7 X pGSO13). The position of the 3‘AGUS, 5’AGUS:Ac or 5’AGUS:Ds constructs in the various
B
i u
u
I
end-joining product (GUS)
recombination product (GUS+)
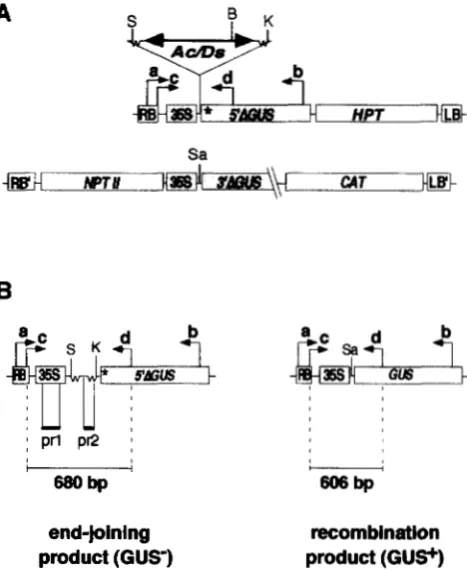
FIGURE 1 .-Assay for Ac-induced homologous recombina- tion. A recombination assay, based on activation of the GUS reporter gene, was developed. (A) The constructs used in this assay drawn not to scale, carrying an inactive GUS gene. The 3’AGUS construct (pGSOO7) contains a GUS gene deleted in the 3‘-end of the gene and cloned in the pGA492 binary vector whose borders are indicated (RB’ and LB’). The 5’AGUSconstn~ts contain a GUSgene with a deletion in the 5‘-end of the gene (*) with an Ac (pGS015=5’AGUS:Ac) or a Ds element (pGS013=5’AGUS:Ds) between the 35s promoter and the deleted GUS gene. The Ac/Ds elements are indicated by a two-sided bold arrow and are flanked by the Waxy se- quences (wavy line) of the maize w x m 7 allele from which Ac originated. The 5‘ deleted GUS gene with the Ac or Ds ele- ments were cloned into binary vector pGPTV-HPT, whose borders are indicated ( R B and LB). The regions of homology between the 3’ and 5‘ deletion constructs are shown as gray boxes and aligned. Primers used for PCR amplification (a-
d ) are indicated by arrows. Major restriction enzyme sites are indicated: S, SpeI; B, BamHI; K, KpnI; Sa, Sun. (B) Amplifica- tion products obtained by two nested rounds of PCR are shown. Ac/Ds excision products expected from end-joining repair are shown on the left: a 680-bp amplification product is expected in the second round of amplification with primers c and d. Recombination is expected to generate a 606bp PCR fragment upon amplification with primers c and d. Two probes, prl and pr2, were used to characterize the excision/ recombination products.
1146 G. Shalev and A. Levy
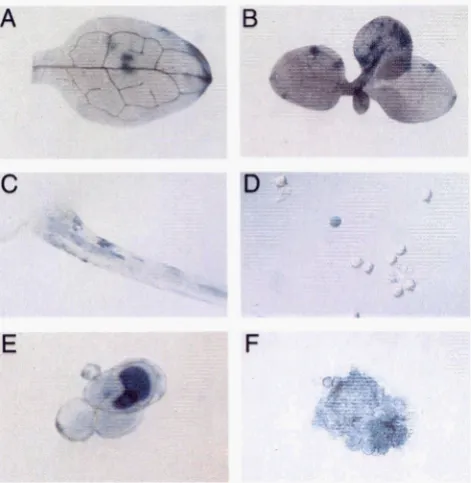
FIGURE 2.-In situ staining for GUS activity in double hemi- zygous F, plants for 5'AGUS:Ac and 3'AGUS. Tissues were stained at various developmental stages according to the histo-
logical assay described by JEFFERSON (JEFFERSON et al. 1987).
GUS' sectors were detected in the cotyledons (A), true leaves
(B), and in roots ( C ) of lway-old seedlings. Similar staining was detected in the pollen (D) and in microcalli derived from protoplasts isolated from Fl plants (E and F).
gamete, cannot be in cis on the same molecule. There- fore, in this assay, recombination partners are necessar- ily located at ectopic locations.
Genetic evidence for Ac-induced ectopic recombina- tion: F1 seedlings were stained histochemically with
X-
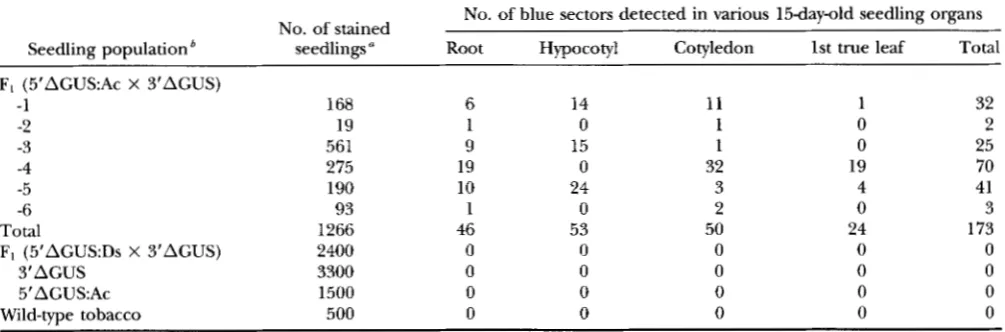
Gluc. Blue sectors were detected exclusively in the eight 3'AGUS X 5'AGUS:Ac combinations and in variousseedling organs, as shown in Figure 2A-C. Each sector is assumed to represent a single genetic event, most probably recombination, which occurred during plant development. In some seedlings, blue spots occurred in clusters (Figure 2B and C ) . Since there were some doubts as to whether these clusters were clonally re- lated, they were counted as a single sector. Otherwise, multiple sectors that seemed clonally unrelated were counted as independent events. For example, in Figure 2A, only two events were counted the first, correspond- ing to the central spots near the midvein, and the sec- ond, at the tip of the cotyledon. The frequency and the distribution of blue sectors in F1 seedlings obtained from six different combinations of the Y A G U S X 5'AGUS:Accross and nine combinations of the 3'AGUS X 5'AGUS:Ds cross are presented in Table 1. The sec- tors observed ranged from a single cell to -100 cells.
, Significant differences ( P
<
0.001; chi squared) in fre-quency of blue sectors were found among the six 3'AGUS X 5'AGUS:Ac combinations (Table 1). The genetic data shown in Table 1 indicate that Ac increases
ectopic HR by at least two orders of magnitude. Fluori- metric analysis of GUS activity in seedlings extracts con- firmed the results from X-Gluc staining, i e . , back- ground GUS levels in parents or
3'A
GUS X5'A
GUS:Ds crosses and increased activity in 3'AGUS X 5'AGUS:Ac crosses (data not shown). The physical characterization of HR events is described below.Physical evidence for ectopic recombination: To physically characterize the observed ectopic HR events, we searched for germinal recombination in F2 seedlings derived from the 3'AGUS X 5'AGUS:Ac crosses. Of 73,000 double hemizygote F2 seedlings, we failed to find germinal recombinant plants using a viable staining screen for GUS (MARTIN et al. 1992). Therefore, we used PCR to amplify recombinant molecules in somatic tissues. Two primer pairs were used for nested PCR amplification: primer pair a-b (see Figure 1) was used in the first round of amplification, and primer pair c d was used in the second one. Ac excision and subsequent repair by end joining of the donor is expected to give rise to the amplification of a fragment of -680 bp (Fig- ure 1). In this end-joining fragment, the Waxy sequence flanking Ac is expected to be a mixture of fragments carrying excision footprints (SAEDLER and NEVER^ 1985; SCOTT et al. 1996). On the other hand, if
H R
repair of Ac-induced DSB is done using the 3'AGUS as a tem- plate, a recombinant 606-bp fragment, corresponding to a conversion-like event should be amplified (Figure 1 ) . To determine the identity of the amplified frag- ments, the EtBr-stained gel containing PCR products from the second round of amplification was blotted and hybridized with two different probes (Figure 3). A W u y oligoprobe (pr2 in Figure 1) hybridized to the 680-bp end-joining fragment only in lanes where genomic DNA from5'A
GUS:Ac or3'A
GUS X5'A
GUS:Ac F1 plants was used as template. No visible bands were observed inNN
Ac-Induced Homologous Recombination 1147
TABLE 1
Frequency and localization of blue sectors in plants carrying the GUS ectopic recombination assay constructs
No. of blue sectors detected in various 15day-old seedling organs No. of stained
Seedling population seedlings ‘’ Root Hypocotyl Cotyledon 1st true leaf Total
F, (5’AGUS:Ac X 3‘AGUS)
-1 168 6 14 11 1 32
-2 19 1 0 1 0 2
-3 561 9 15 1 0 25
-4 275 19 0 32 19 70
-5 190 10 24 3 4 41
-6 93 1 0 2 0 3
Total 1266 46 53 50 24 173
F, (5‘AGUS:Ds X 3’AGUS) 2400 0 0 0 0 0
3‘AGUS 3300 0 0 0 0 0
5’AGUSAc 1500 0 0 0 0 0
Wild-type tobacco 500 0 0 0 0 0
‘’ Kanamycin-resistant seedlings were histochemically stained: no kanamycin selection was done in the wild-type seedlings. One third of the kanamycin-resistant seedlings are double hemizygous for the 5’AGUS:Ac and 3’AGUS constructs or for 5’AGUS:Ds and 3’AGUS.
’
Data are given separately for six combinations of the 5’AGUS:Ac v X 3’AGUS crosses. For 5’AGUS:Ds X 3’AGUS crosses, pooled data from nine genetic combinations are presented.the fact that the 5‘AGUS:Ac and 5’AGUS:Ds constructs are almost identical in sequence. This supports the no- tion that the formation of the 606 bp is excision depen- dent rather than sequence dependent.
To further confirm the nature of the 680-bp religa- tion and 606-bp recombination bands, the PCR prod- ucts corresponding to these bands were cloned and sequenced. The sequences of three independent clones of the 606-bp fragment, obtained from two different F1 combinations, were found identical, corresponding to a recombinant molecule between the T-DNA right bor- der of 5’AGUS:Ac (pGSO15) and the region around SalI in ?‘AGUS, which served as a template for gap repair (see Figure 4A). Note that the T-DNA right border of 5‘AGUS:Ac (pGS015) is different in sequence com- pared to the right border of 3’AGUS (Figure 1) and that no such chimeric molecule existed previously in our laboratory. Sequencing of three clones of the -680- bp fragment revealed typical Ac excision footprints
(SCOTT et al. 1996) as shown in Figure 4B. This confirms that the 680-bp band, which hybridizes to the Waxy oligoprobe, is truly an end-joining product. The physi- cal evidence shown here, for Ac-induced formation of recombinant molecules-conversion-like products-is in agreement with the genetic data obtained by GUS staining (Table l ) , suggesting that ectopic HR does not occur, or is very rare, in the absence of Ac-induced DSB. Although additional recombination products (via crossing over) might also be Ac induced, our prelimi- nary attempts at long range PCR to amplify such prod- ucts failed (data not shown).
The frequency of ectopic recombination: The fre- quency of ectopic recombination was estimated by vari- ous means and in different tissues.
Pollen grains from double hemizygote plants from the 3’AGUSx 5’AGUS:Acor the YAGUSX 5’AGUS:Ds crosses were stained with X-Gluc for GUS activity. Twelve pollen grains with strong blue color (Figure 2D) were found out of a population of 5 X lo5 pollen grains in the 3‘AGUS X 5‘AGUS:Ac cross while no blue pollen grains were found in a population of 3 X l o 6 grains in the 3’AGUS X 5’AGUS:Ds cross. This represents a recombination frequency of 2.4 X in pollen cells. Microcalli derived from mesophyll protoplasts were used to estimate the frequency of somatic ectopic re- combination. GUS activity was assayed by X-Gluc stain- ing in four to 16 cells microcalli of F, 3’AGUS X
5‘AGUS:Ac plants. A total of -500,000 microcalli were stained and screened for the presence of blue cells as shown in Figure 2E and F. The frequency of blue cells was 2.4 X
PCR experiments also provide rough estimates on the frequency of Ac-induced ectopic recombination (Figure 3). These experiments were carried out using 500 ng genomic template in the first round, corresponding to -33,000 tobacco genomes (C value = 15.5 pg) (BENNETT and SMITH 1976). All the PCR reactions, where genomic DNA prepared from the 3’AGUS X
5’AGUS:Ac combinations was used as template, gave rise to the 606-bp recombinant band; hence at least one recombinant molecule was present in the template. This further supports the estimates that Ac-induced ec- topic recombination occurred in 10-3-10-4 of the cells.
DISCUSSION
1148 G. Shalev and A. A. Levy
A.
1 2 3 4 5 6 7 8 9--e - . - ~ . ~ . ~
(I) < P.T -680
B a . .
-
680-
606FIGURE 3.-Identification of recombination and excision products. PCR products were obtained by two nested rounds of PCR using primers a-d (see Figure 1) and genomic DNA
template from wild-type NN tobacco ( l ) , 3'AGUS (2),
5'ACUS:Ds (3). a double hemizygous plant for 5'AGUS:Ds and 3'AGUS (4-6), 5'AGUS:Ac (7) and a double hemizygous plant for S'AGUS:Ac and 3'AGUS (8 and 9). Products were run on an agarose gel, blotted and hybridized with two probes.
(A) An oligoprobe (pr2 of Figure l ) , whose sequence is de- rived from the Ac flanking sequence of the Waxy gene, was
used to identify the expected -680-bp excision products. (B)
A second probe derived from the 35s promoter (prl of Figure 1) was used to detect all amplification products (excision and recombination).
tween two nonallelic recombination partners that shared 2 kb of homology (0.5 kb upstream of Ac exci- sion site and 1.5 kb downstream of it, as shown in Figure 1). This assay has provided both genetic (GUS reactiva- tion) and physical evidence that Ac induces recombina- tion between homologous sequences located at ectopic sites. The findings that GUS reactivation occurred only with Ac in 3'AGUS X 5'AGUS:Ac crosses but not with an almost identical Ds element provided the genetic evidence for recombination. The physical evidence is that a 606-bp PCR fragment was obtained exclusively in all the 3'AGUS X 5'ACUS:Ac combinations tested, and its sequence confirmed that it is a recombinant mole- cule. The 606-bp fragment has the size and sequence expected for such a recombination product and it was not obtained in any of the crosses carrying Ds (3'A GUS X s'AGUSrDs), whose sequence is almost identical to Ac. This fragment was not amplified either in S'AGUS:Ac, i.e., in the absence of the 3'AGUStemplate, or in the other controls (Figure 3B). Therefore, its for- mation is both excision (DSB)- and templatedepen-
A
~ G C A A C G T T G A A G G A C C X T C A G C C C A W ~ W T
--(AGAAGG
ATCCCCCC
f=tk*&E*-
L_ " " " "
--
CAP.gATECMTTETATTT&CAP.'.TA~ ~
-
i C ' A ~ c ~ c ? .i
-
-
- - -- ~~-
-... ~ ."" _""....""...
GT-c ~
_ _ _ . _ . . .
B
waxr
CGCGGCCGGdCACGCAACGCGC5 'AWS:Ac C G C G G C ~ ( A c ) ~ C O C G
RLl CGcGGcGGCITcACG- GC -GTCACGCAACGCG
RL2 GCGGCTGGGTCA
RL3 CGCGGCCCYXITC"
"-
FIGURE 4.-Sequencing of Ac-induced recombination and excision products. PCR products corresponding to the puta- tive recombination product (the -606bp fragment in Figures 1 and 3) and the excision products (the -680-bp fragment in Figures 1 and 3) were cloned and sequenced. (A) The sequence of the 606bp fragment. This sequence was identical in three independent clones, confirming the recombinant nature of this fragment. It consists of the right border (gray box) of the 5'AGUS:Ac T-DNA (binary vector pGPTV-HPT), the sequence of the 35s promoter (empty crossed box) and of the TMV translational enhancer R (the dashed box), the Sull site (bolded) as in 3'AGUSand the intact open reading frame GUS sequence of 3'AGUS (dotted box). (B) The se- quence of three independent clones of the -680-bp fragment revealing typical Acexcision footprints. The wild-type Waxy sequence is shown with the Ac insertion site underlined. The sequence flanking Ac in 5'AGUS:Ac is shown with the host duplication underlined. The sequence of three independent clones of the -680-bp fragment (RL1, RL2 and RL3) shows typical excision footprints, with two base substitutions (RLl), a 4 b p deletion represented by dashes (RL2) and a perfect restoration of the Waxy sequence (RL3).
dent, as expected for a recombination product. This argues against the possibility that the 606-bp recombi- nant fragment was formed by a PCR artifact. Moreover, neither this fragment nor a similar molecule had ex- isted in our laboratory, hence excluding a PCR contami- nation.
DSBs induce homologous recombination in plants: DNA DSBs have been shown previously to induce re- combination in plants using assays based on different types of homologous partners: (1) Somatic crossover between homologous chromosomes was induced by DNAdamaging agents such as y-irradiation (CARLSON
Ac-Induced Homologous Recombination 1149
duced in vitro with restriction enzymes (ENGELS and MEYER 1992) or in vivo with site-specific recombinases (PUCHTA et al. 1993). Single-strand annealing was pro- posed as the major pathway for extrachromosomal re- combination in plants (PUCHTA et al. 1994). (3) Homol- ogous integration of a T-DNA sequence into a chromo- somal site was shown to be increased by the formation of a DSB in the chromosomal target (PUCHTA et al. 1996). (4) Intrachromosomal recombination between large direct repeats was induced upon Ac excision at the Plocus in maize (ATHMA and PETERSON 1991). Simi- larly, Mutator was shown to increase recombination be- tween tandem repeats at the K n l - 0 locus (LOWE et al. 1992). Intrachromosomal recombination was also found to be induced by DNA-damaging agents (TOVAR and LICHTENSTEIN 1992; PUCHTA et al. 1995). Recently, the HO endonuclease was shown to increase intrachro- mosomal recombination by 10-fold in Arabidopsis (CHI- URAZZI et al. 1996). (5) Sisterchromatid exchange was shown to be induced by DNA-damaging agents (SCHARTZMAN 1987), suggesting that recombination be- tween sister chromatids can play a role in DNA repair. It was also shown that Ac excision can induce gap repair using a sister chromatid as a template (E. RUBIN and A. A. LEVY, unpublished data).
In this article we show for the first time the induction of ectopic HR in plants by DSB formation. We did not find evidence for ectopic recombination in 3'AGUS X 5'AGUS:Ds crosses using the GUS reactivation assay or by PCR. Moreover, ectopic recombination in the a b sence of DSBs has not been reported previously in plants, suggesting that ectopic recombination in the plant genome does not occur or occurs very rarely. This is in contrast to yeast, in which HR, including ectopic recombination, is efficient (ERNST et al. 1981; MIKUS and PETES 1982) and to Drosophila in which ectopic HR was reported even in the absence of transposon excision ( LIM and SIMMONS 1994).
Despite the extensive abovequoted literature on DSBs-induced HR in plants, the question remains how efficient is HR as a DSB repair mechanism in plants, compared to the alternative nonhomologous end-join- ing or illegitimate-recombination pathways. In the case of Ac the frequent occurrence of revertant sectors and the extensive analysis of excision footprints (SAEDLER and NEVERS 1985; SCOTT et al. 1996) suggest that nonho- mologous end joining is the major DSB repair pathway. If this is the case, it is surprising that the recombinant 606bp and the religation -680-bp fragments showed equal stochiometry in Southern blots of the PCR reac- tions (Figure 3B). This discrepancy might be caused by the fact that PCR is not quantitative, particularly after two rounds of amplification, or that there is PCR tern- plate preference.
As discussed above, DSBs are repaired by both illegiti- mate and homologous recombination in plants. From the data presented here, one may extrapolate that any
DSB occurring in a repetitive DNA region can induce ectopic recombination in plants. The implications of such a possibility are described below.
Genetic and evolutionary implications of ectopic H R : Ectopic recombination could lead to chromo- somal translocations in cases of crossover or to gene- conversion. In organisms such as plants or mammals, in which a high percent of the genome is repetitive, crossover between ectopic sequence would create a ge- nomic chaos. Therefore, ectopic recombination should occur at a very low rate, or a mechanism favoring con- version over crossover should be used. In mice, ectopic HR was found at a frequency of 0.1
-0.7%
(MURTI et al. 1994). These ectopic recombination events corre- sponded to gene conversion, thus providing a mecha- nism to maintain genome stability. Nevertheless, evi- dence was found for ectopic crossover, leading to chro- mosomal translocations (KVALOY et al. 1994; WANG et al. 1995) in humans. Such events are probably rare or are efficiently selected against. In plants, translocation lines have been isolated in several species, though, no sequence analysis of translocation breakpoints is avail- able. Therefore, it is not known whether translocation occurred as a result of crossover between homologous ectopic sequences or as a result of nonhomologous end joining of broken chromosomes. The ectopic recombi- nation events characterized in this work occurred at a rate of 10-3-10-5. As Ac excision is usually in the 10" range (JONES et al. 1989; LEVY and WAIBOT 1990) ; this suggests that, once a DSB is formed, it can be repaired using an ectopic homologous template for 0.1-10% of the events. Such frequencies make ectopic recombina- tion in plants a potentially important factor in genome evolution. Ac/Ds insertion, or any other type of DSB within repeated DNA, could lead to shuffling of the genome if recombination is followed by crossover. This work could not determine whether conversion or cross- over occurred since recovery of germinal recombina- tion events and rescue of the GUSf calli were not suc- cessful. A system similar to the one described here, but with a selectable marker, e.g., antibiotic resistance, could allow regeneration of somatic events or screening of large plant populations for germinal events.1150 G. Shalev and A. A. Levy
For practical applications, the DSB-induced HR re- ported here can facilitate targeted integration at any locus where an active transposable element insertion is present. Targeting could be done via T-DNA integra- tion as described by PUCHTA et al. (1996). Alternatively, a new strategy of gene targeting, based on ectopic re- combination, can be conceived. Under this scheme, one line where the DSB is induced at a specific locus serves as the targeted site. This line is then crossed to another line carrying an engineered construct (the template), which can recombine with the targeted lo- cus. Resolution of such recombination via gene conver- sion will result in gene targeting.
We thank EITAN RUBIN and Dr. SHAHAL ABBO for help and discus- sion; NAOMI A m , 1 ~ 4 ~ LEW and MICHAI. RAZ for excellent technical help and YIGAI. AVM for editing the manuscript. This work was sup- ported by a grant from AFIRST to A.A.L.
LITERATURE CITED
A N , G., 1987 Binary Ti vectors for plant transformation and pro- moter analysis. Methods Enzymol. 1 5 3 292-305.
ATHMA, P., and T. PETERSON, 1991 Acinduces homologous recombi- nation at the Maize Plocus. Genetics 128: 163-173.
AWV, D., and E. GAI.UN, 1985 An in vitro procedure to assign pig- ment mutations in Nicotiana to either the chloroplast or the nucleus. Heredity 76: 135-136.
BANGA, S. S., and J. B. BOYD, 1992 Oligonucleotidedirected site-spe- cific mutagenesis in Drosophila melanoguster. Proc. Natl. Acad. Sci.
BECKER, D., E. KEMPER, J. SCHEI.1. and R. IMASTERSON, 1992 New plant binary vectors with selectable markers located proximal to the left T-DNA border. Plant Mol. Biol. 2 0 1195-1197. BEECKMAN, T., and ENGLER, 1994 An easy technique for the clearing
of histochemically stained plant tissue. Plant Mol. Biol. Rep. 12:
BENNETT, M. D., and J. B. SMITH, 1976 Nuclear DNA amounts in angiosperms. Philos. Trans. R. SOC. Lond. B. Biol. Sci. 274: 227-
274.
CARLSON, P. S., 1974 Mitotic crossing-over in a higher plant. Genet. Res. Camb. 2 4 109-112.
CHEN, J., I. M. GREENBLATT and S. L. DELLAPORTA, 1992 Molecular analysis of Ac transposition and DNA replication. Genetics 130:
665-676.
Enhancement of somatic intrachromosomal homologous recom- bination in Arubidopsis by the HO endonuclease. Plant Cell 8:
2057-2066.
CHOMET, P. S., S. WESSLER and S. L. DELLAPORTA, 1987 Inactivation
with i t s DNA modification. EMBO J. 6: 295-302.
of the maize transposable element Activator (Ac) is associated
COEN, E. S., T. P. ROBBINS, J. ALMEIDA, A. HUDSON and R. CARPENTER, 1989 Consequences and mechanism of transposition in Antir- rhinum mujus, pp. 413-426 in Mobile DNA, D. E. BERG, M. M. HOW. American Society for Microbiology, Washington, DC. DELLAPORTA, S. L., J. WOOD and J. B. HICKS, 1983 A plant DNA
minipreparation: version 11. Plant Mol. Biol. Rep. 1: 19-21. EICHENBAUM, Z., and Z. LNNEH, 1995 Intermolecular transposition
of IS10 causes coupled homologous recombination at the trans- position site. Genetics 140: 861-874.
ENGELS, P., and P. MEXER, 1992 Comparison of homologous recom- bination frequencies in somatic cells of petunia and tobacco suggest two distinct recombination pathways. Plant J. 2: 59-67. ENGELS, W. R., D. M. JOHNSON-SCHI.ITZ, W. B. EGGL.ESTON and J. SVED,
1990 High-frequency P element loss in Drosophila is homolog dependent. Cell 62: 515-525.
ERNST, J., J. STEWART and F. SHERMAN, 1981 The cycl-11 mutation in yeast reverts by recombination with a nonallelic gene: compos- ite gene determining the isocytochrome-c. Proc. Natl. Acad. Sci. USA 78: 6334-6338.
USA 89: 1735-1739.
37-42.
CHIURAZZI, M., A. RAY, J-F. VIRET, R. PERERA, X-H. WANG et U L , 1996
FEINBERG, A. P., and B. VOGELSTEIN, 1983 A technique for radiola- beling DNA restriction endonuclease fragments to high specific activity. Anal. Biochem. 132: 6-13.
FORMOW, T., and B. M. ALBERTS, 1986 DNA synthesis dependent on genetic recombination: characterization of a reaction cata- lyzed hy purified bacteriophage T4 proteins. Cell 47: 793-806. FRADKIN, C.-M. W., and R. L. BRINK, 1956 Crossing over in maize in
the presence of the transposable factors activator and mudolator.
Genetics 41: 901-906.
FRIDLENDER, M., K H ~ S O N , J. D. G. JONES and A. A. LEW, 1996 Repression of the Ac transposase gene promoter by Ar transpo- sase. Plant J. 9: 91 1-917.
GALLIE, D. R., D. E. SLEAT, J. W. WAITS, P. C. TURNER and T.-M. A. WIISON, 1987 The 5”leader sequence of tobacco mosaic virus RNA enhances the expression of foreign gene transcripts in-vitro and in-vivo. Nucleic Acids Res. 15: 3257-3273.
GREENBIATT, I., 1981 Enhancement of crossing-over by the trans- posable element, Modulator, in maize. Maydica 26: 133-140. HAGEMANN, A. T., and N. L. CRAIG, 1993 Tn7 transposition creates
hotspots for homologous recombination at the transposon donor site. Genetics 133: 9-16.
HOEKEMA, A,, P. R. HIRSCH, P. J. J. HOOWS and R. A. SCHILPERO- ORT, 1983 A binary plant vector strategy based on separation of vir- and T-region of the Agrobuctm’um tumfaciens Ti-plasmid. Nature 303: 179-180.
HORSCH, R. B., J. E. FRY, N. L. HOFFMANN, D. EICHHOI.TZ, S. G. Roc;
ERS et al,, 1985 A general and simple method for transferring genes into plants. Science 227: 1229-1231.
JEFFERSON, R. A,, T. A. KAVANAGH and M. W. BEVAN, 1987 GUS fu- sions: @-Glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 6: 3901-3907.
JONES, J. D. G., F. M. CARLAND, P. MALIGA and H. K. DOONER, 1989 Visual detection of transposition of the maize element Activator
(Ac) in tobacco seedlings. Science 244: 204-207.
KEI.I.ER, J., E. LIM and H. K. DOONER, 1993 Preferential transposi- tion of Ac to linked sites in Arubidopsis. Theor. Appl. Genet. 86:
KVALOY, R, F. GALVAGNI and W. BROWN, 1994 The sequence organi- zation of the long arm pseudoautosomal region of the human sex chromosomes. Hum. Mol. Genet. 3: 771-778.
LEW, A,, and V. WALBOT, 1990 Regulation of the timing oftranspos- able element excision during maize development. Science 248:
1534-1537.
LIM,J., and M. SIMMONS, 1994 Gross chromosome rearrangements mediated by transposable elements in Drosophila mlanogaster. Bi- oEssays 16: 269-275.
LOW, B., J. MATHERN and S. HAKE, 1992 Active Mutator elements suppress the knotted phenotype and increase recombination at the Knl-0 tandem duplication. Genetics 132: 813-822. MARTIN, T., R. SCHMIDT, T. ALTMANN and W. FROMMER, 1992 Non-
destructive assay systems for detection of p-Clucuronidusc in higher plants. Plant Mol. Biol. Rep. 10: 37-46.
MCBRIDE, K., and K. SUMMERFELT, 1990 Improved binary vectors for Agrobacterium-mediated plant transformation. Plant Mol. Biol.
MCCLI~VTOCK, B., 1953 Mutation in maize. Carnegie Inst. Washing- ton Year Book 52: 227-237.
MIKUS, M., and T. PETES, 1982 Recombination between genes l o - cated on nonhomologous chromosomes in Saccharomyces cffpui.y-
iae. Genetics 101: 369-404.
M U R T I , ~ . R., M. BUMBULIS and J. C . SCH~MENTI, 1994 Gene conver- sion between unlinked sequences in germline of mice. Genetics
137: 837-843.
NASSIF, N., J. PENNEY, S. PAL, W. R. ENGELS and G. B. GI.OOR, 1994 Efficient copying of nonhomologous sequences from ectopic sites via P rkmt-induced gap repair. Mol. Cell. Biol. 14: 1613-
1625.
PLASTERK, R. H. A., 1991 The origin of footprints of the ?il transpo- son of Cnenorhabditis ekgans. EMBO J. 10: 1919-1925.
PUCHTA, H., D. BERNARD and H. BARBARA, 1993 Homologous re- combination in plant cells is enhanced by in-vivo induction of double strand breaks into DNA by a site specific endonuclease. Nucleic Acids Res. 21: 5034-5040.
PUCHTA, H., P. SwoBoDAand B. HOHN, 1994 Homologous recombi-
PUCHTA, H., P. SWOBODA and B. HOHN, 1995 Induction of intra- nation in plants. Experientia 5 0 277-284.
585-588.
Ac-Induced Homologous Recombination 1151
chromosomal homologous recombination in whole plants. Plant
PUCHTA, H., B. DUJON and B. HOHN, 1996 Two different but related mechanisms are used in plants for the repair of genomic double- strand breaks by homologous recombination. Proc. Natl. Acad. Sci. USA 93: 5055-5060.
SAEDLER, H., and A. GIERL, 1996 Transposable Elements. Springer- Verlag, Berlin.
SAEDLER, H., and P. NEVERS, 1985 Transposition in plants: a molecu- lar model. EMBO J. 4 585-590.
SAMBROOK, J., E. F. FRITSCH and T. MANUTIS, 1989 Molecular Clon- ing: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N Y ,
J. 7: 203-210.
SCHARTZMAN, J. B., 1987 Sisterchromatid exchange in higher plant cells: past and perspectives. Mutat. Res. 181: 127-145.
S c o r r , L., D. LAFOE and C. WEIL, 1996 Adjacent sequences influ- ence DNA repair accompanying transposon excision in maize. Genetics 142 237-246.
TOVAR, J.. and C. LICHTENSTEIN, 1992 Somatic and meiotic chromo- somal recombination between inverted duplications in transgenic tobacco. Plant Cell 4 319-332.
WANG, I., D. WEIL, J. LEVILLIERS, N. ha, A. DE LA CHAPELLE et aL, 1995 Prevalence and molecular analysis of two hot spots for ectopic recombination leading to XX maleness. Genomics 2 8
52-58.