Copyright 0 1995 by the Genetics Society of America
On
the
Clustered Exchanges
of the
RecBCD Pathway Operating
on
Phage
h
Franklin
W.
Stahl,Claire E. Shurvinton,’
Lynn
C. Thomason,
Stuart H i l l 2 andMary
M.
StahlInstitute of Molecular Biology, University of Oregon, Eugene, Oregon 97403-1 229
Manuscript received July 26, 1994 Accepted for publication October 31, 1994
ABSTRACT
Lytic cycle crosses of Red- Gam- phage A were conducted in rec+ Escherichia coli carrying one or another plasmid with homology to A. A X A recombinants and A X plasmid recombinants were formed by RecBCD-mediated recombination. We showed previously that the act of recombining with a plasmid alters the disposition of selected A X A exchanges. This work reports that the relationships between the A X plasmid and the A X A exchanges is unaltered by the removal from one A parent of the homology shared with the plasmid. This result supports our view that a reciprocal exchange, allowing for cointegrate formation, is associated with but mechanistically separable from a (presumably) nonreciprocal A X A exchange. The nature of this relationship is independent of A’s Rap function, which is shown to alter the ratio of cointegrate formation (splices) to marker pick-up (patches) in A X plasmid recombination mediated by the RecBCD pathway.
I
N eukaryotes, the formation of complementary re- combinant types in individual events is demonstra- ble by analysis of meiotic tetrads. Such a demonstration allows us to say that meiotic recombination, as it usually occurs between markers that are not too close together, is “reciprocal.” Such reciprocal recombination is often referred to as “crossing over” or “reciprocal exchange” to distinguish it from “conversion,” a (set of) nonre- ciprocal mechanism( s ) responsible for most of the re- combination observed between very close markers.A demonstration of reciprocality, as defined above, is not easy with prokaryotes, which lack the orderliness of meiosis. Various approaches have been tried. Among phages, the “classical” approach involves analysis of the viral yields from individual infected cells. The recombi- nant yields from such “single bursts” vary from cell to cell. With appropriate controls, a positive correlation between the yields of complementary recombinants from single cells argues for reciprocality. Single-burst analyses are time consuming and rarely subjected to independent verification. The discovery of DNA se- quences that stimulate recombination in their vicinity (recombinators) allow for tests of reciprocality based on the analysis of the pooled yields of many infected cells (mass lysate analysis)
.
Reciprocality may be analyzable in mass lysates when the two participating genotypes of infecting phage play
Corresponding author: Franklin W. Stahl, Institute of Molecular Biol- ogy, University of Oregon, Eugene, OR 97403-1229.
E-mail: [email protected]
‘
Present address: 1832 Chadwick Ct., Santa Rosa, CA 95401.‘
Present address: Laboratory of Microbial Structure and Function, National Institutes of Health, Rocky Mountain Laboratories, Hamil- ton, MT 59840.Genetics 1 3 9 1107-1121 (March, 1995)
asymmetric roles in the recombination process. In par- ticular, if one parent has a recombinator sequence and the other does not, nonreciprocality in the recombina- tion process might show up in the mass lysate as an inequality in the numbers of complementary recombi- nant phage particles (e.g., STAHL and STAHL 1985) .
For the RecBCD pathway of E. coli, reciprocality can be examined in mass lysates when one parent carries
x,
an asymmetric octameric DNA sequence that activates RecBCD protein to catalyze recombination (see SMITHand STAHL 1985; MYERS and STAHL 1994 for reviews). The analysis of reciprocality when RecBCD operates on A is complicated by features of the phage’s life cycle.
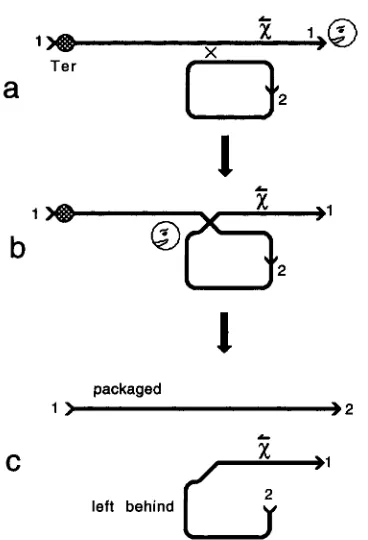
A chromosome packaging is initiated when terminase binds to and cuts a cos sequence, remaining bound to the left end of the linearized chromosome. Each such opening of cos allows the entry of RecBCD into the right end of the chromosome. The enzyme travels from the right end to
x,
where it is activated to catalyze recombi- nation. Because packaging and recombination are both dependent on cos cutting, it is no surprise that the re- combinant that is packaged is usually the one that in- herits the left end of A from the Xcontaining parent(Figure 1, KOBAYMHI et al. 1983). KOBAYASHI et al.
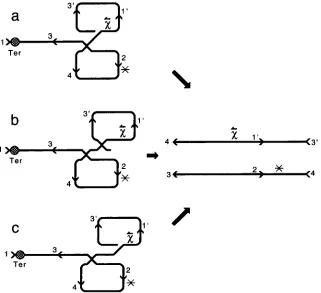
(1984) removed this bias by separating recombination from packaging. The standard cos site in the parent lacking
x ( x o
parent) was deleted for the terminase cut-site, preventing the recombinant from packaging via standard cos. Packaging was achieved via cloned cossites located near the middle of the A chromosome. The cloned cos sites were oriented parallel to
x
so they could not activate it (Figure 2 ) . From such crosses,1108 F. W. Stahl et al.
1- X
‘i
1,@Te r
a
[j;
1- 3
.
.
X
‘i
)1b
Ter
a
4C
1
packaged
1 ) ) 2
L
C
4 <
‘i
.1.
< 3FIGURE 1 .-Chromosome linearization by terminase cut- ting at cos both initates packaging and allows for the entry of RecBCD at A’s right end. ( a ) Terminase (Ter) linearizes a A chromosome by cutting cos ( 1 ) . RecBCD protein enters the DNA at the right end and travels leftward until it encounters
x.
The enzyme is activated by the encounter and catalyzes exchange as it continues traveling leftward. ( b ) The exchange is presumed to be reciprocal. However, whenx
is present in only one of the two parents, the resulting association between recombination and packaging biases the yield of recombi- nants in favor of the one that does not inheritx
(c; KOBAYMHI et al. 1984).present in comparable numbers. They concluded that RecBCD-mediated exchange was reciprocal and that a fraction of the time cos is resealed, allowing packaging of one recombined chromosome from each dimer (but see below)
.
The distinguishing phenotype of E. coli re& mutants is the loss of doublechain exonuclease activity (exoV) with the acquisition of heightened recombination activ- ity (AMUNDSEN et al. 1986).
A
recombination in re&mutants is similar to that occurring in rec+ cells in the neighborhood of
x
(AMUNDSEN et al. 1986; THALER etal. 1989). THALER et al. ( 1988,1989) proposed that the RecD subunit is lost from the RecBCD protein when that traveling protein encounters
x.
S. LIU and A. V.KUZMINOV (see STAHL et al. 1990b) argued on grounds of economy that the proposed loss of the RecD subunit at
x
(or a X-like sequence) implies the loss of exoV activity. They pointed out that the apparent reciprocal- ity observed by KOBAWHI et al. ( 1984) was harmonious with the concept of exoV digestion tox
only if theFIGURE 2.-An early interpretation of the approximately equal recovery of complementary X-stimulated recombinants
( KOBAWHI et al. 1984). ( a ) Terminase (Ter) linearizes a
x-
containing A chromosome by cutting cos at its standard loca- tion ( l ) . RecBCD protein enters the DNA at the right end and travels leftward until it encountersx
in its standard (left- ward) orientation. That encounter leads to an exchange to the left of thex
site. The exchange is presumed to be recipro- cal, because complementary, X-stimulated recombinants are recovered among progeny particles in nearly equal numbers. Ter-governed packaging from the cut cos site ( 1 ) to its homo- logue is impossible because site 2 is uncuttable ( * ).
( b ) Some of the time, the linearized, Xcontaining circle recloses at cos, generating a dimer circle. The two complementary recombi- nants can be packaged equally well from cos site 3 to cos site4 or from cos site 4 to cos site 3, respectively. ( c ) The resulting packaged recombinant chromosomes contain
x
or lackx,
respectively.observed recombination events were typically triparen- tal. The third parent in the interaction could replace the right end of A that was destroyed by exoV when it entered A’s right end after the opening of cos. If the third A chromosome were of the same genotype as the partially destroyed one, a pair of complementary recom- binants could result (Figure 3 )
.
In this scheme, the degree of equality of complemen- tary types should be sensitive to the relative multiplicity of infection of the two parental phage types. Indeed, only when the
x +
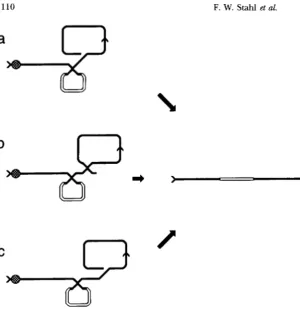
parent was in excess, were comple- mentary recombinants recovered equally ( STAHL et al. 1990b). The observed equality suggested that the lost DNA is recovered by an exchange that occurs with unit efficiency.Clustered Exchanges by RecBCD 1109
b
1 '
1
.--c=y*
1
e
3 i 2
.*
.
<4FIGURE 3.-A current interpretation of the approximately equal recovery of com- plementary X-stimulated recombinants when the X-containing parent is in excess ( STAHL
et al. 1990b)
.
( a ) As in Figure 2, terminase linearizes A. ( b ) RecBCD protein enters the linearized A at the right end ( 1 ) and digests the DNA to X, whereupon the RecD subunit is lost from the protein. The RecBC ( D - ) protein continues to travel leftward until it catalyzes a reciprocal exchange with a chro- mosome of the other infecting type or until it dissociates from the chromosome. ( c ) A single chain at the frayed doublechain end resulting from digestion by exoV invades a duplex, initiating nonreciprocal recombina- tion with a chromosome of the X-containing type. ( d ) From the resulting trimer, two complementary recombinant chromosomes can be packaged. ( e ) A X-containing re- combinant is packaged from cos3'
to cost. A non-xcontaining recombinant is packaged from cos4 to cos3. The depiction of the pro- posed reciprocal exchange ( a and b ) as oc- curring before the nonreciprocal o n e ( c and d ) is arbitrary.tion of w h e t h e r e x c h a n g e i n that pathway is ever truly Evidence supporting the schemes in Figure 4b and c reciprocal. The observed result could be obtained if over a was obtained with triparental crosses involving n e i t h e r e x c h a n g e w e r e r e c i p r o c a l ( F i g u r e 4 a ) , both two infecting A g e n o t y p e s a n d a resident plasmid that
were reciprocal (Figure 4b) or one was reciprocal and carried a segment of the A chromosome ( STAHL et al.
t h e o t h e r n o t ( F i g u r e 4 c )
.
1 9 9 0 b ) . Progeny phage particles that had p i c k e d u pI
b
n
4
d*
FIGURE 4.-Three routes compatible with the observation that two
x +
chromosomes interact with onex o
chromosome to pro- duce both a recombinant that inheritsx
and its complement. In each figure, theL right-hand end of the linearized chromo-
x
1'. ( 3 b some has been digested by exoV. ( a ) Con- Itiguous nonreciprocal exchanges are shown. ( b ) Nearby reciprocal exchanges are shown.
2.
*
, 1
3 1 I <4 nearby nonreciprocal ( c ) A reciprocal exchange ihown with a one. In b and c, the
1110 F. W. Stahl et al.
I
FIGURE 5.-Three interpretations of theclustered exchanges seen when a plasmid is
picked up in a A X plasmid X A cross. ( a )
Both exchanges in the cluster are nonrecipro- cal. ( b ) Both exchanges in the cluster are reciprocal. ( c ) One exchange is reciprocal, the other not. The schemes in B and C are not likely to be experimentally distinguishable. In
b and c, the reciprocity of the A X plasmid
exchange is symbolized by the presence of
DNA from the Ter-linearized parent to the
right of that exchange.
the plasmid in these crosses were distinguished from those that had not. In each class, recombinants for ter- minal A markers were selected and scored with respect to an intervening marker (Figure 5 )
.
If all recombina- tion occurred by nonreciprocal exchanges (Figure 5a),
plasmid pickup would require the participation of two
X chromosomes. When that event produced a plasmid- containing A recombinant, the exchange would neces- sarily be in the interval that has homology with the plasmid (S. M. ROSENBERG, personal communication)
.
When the plasmid pick-up was in the rightward of two intervals, that result was obtained, providing proof that plasmid pick-up and the A X A exchange that was se- lected occurred in a single, complex act. That result was not obtained, however, when the plasmid was picked up in the left interval. Only about one-fourth of the recombinants that had picked up plasmid were recom- binant in the interval into which the plasmid had been recombined. That result favored the view that the ex- change with the plasmid was reciprocal, rather than nonreciprocal, consonant with Figure 5, b and c, and arguing against the scheme in Figures 4a and 5a. On the other hand, the result was compatible with the possi- bility that a large minority ( 524% in the data reported) of the plasmid pick-ups was achieved by a pair of nonre- ciprocal exchanges.The plasmid pick-up experiments reported by STAHL et al. (1990b) and those described above demonstrate clustering of exchanges-the pickup of a plasmid, by
a reciprocal exchange-is often accompanied by a nearby A X A exchange, which is effectively nonrecipro- cal (Figure 5, b or c )
.
This demonstration of clustered exchanges in the RecBCD pathway puts in doubt the interpretation of experiments purporting to study “patches” ( STAHL et al. 1982, 1984, 1990b; ROSENBERG 1987, 1988; HAGEMANN and ROSENBERC 1991), z.e., short single-chain segments of DNA transfered from one chromosome to another unaccompanied by cross- ing over of flanking markers ( STAHL 1979). Because splices are demonstrably clustered in these RecBCD crosses, we must consider the possibility that most such “patches” are really close double splices.The results of experiments reported herein support the view that plasmid X A recombination leading to plas- mid pick-up is usually reciprocal by showing that the route diagrammed in Figure 5a makes no appreciable contribution to A recombination accompanying plasmid pick-up. This result is the one expected from the model tested previously, which supposes that RecBCD unwinds and hydrolyzes DNA until the RecD subunit is lost from the RecBCD holoenzyme. The loss of RecD eliminates exoV activity and renders RecBC ( D - ) protein capable
of catalyzing exchange as it continues to travel.
Clustered Exchanges by RecBCD
TABLE 1
A genetic elements
Element Size (kb) Properties Source/Reference
1111
cop' (hy33) cos2
Asusll Msus481 Jtsl5 Jsus6
A b2 Ab519 AB Ab538 cosML AC biol int4 Aspi6 red3 gam21 0
imm434 imm"
CI 85 7
CI 26
nin5 d2 rap d5
X+D
ssus 7
Rsus5 Rts2 X+C 6 3 5 6.9 5 net A 2.5
4
net A 2.5
2.8 0.9
0.3
cos, 1 and part of 2
22-bp deletion of cos cut site Amber in terminase gene A
Amber in gene M
Heat-sensitive allele of J
Amber allele of J
No phenotype No phenotype
Deletion EcoRI sites 1-2 Deletion leftward from att
cos cloned in A in reverse orientation Deletion EcoRI sites 2-3 (Int- Red-) Int- Red- Gam-
Unconditional int allele A (re&-cIII)
Mutation in redp, polar on re&
Amber allele of gam
Immunity region of phage 434 Immunity region of phage 21
Temperature-sensitive clear mutation Unconditional clear mutation Chi mutation in cII
Deletion
Deletion in ninB-ninC ninG
Deletion of ninGninH
Chi mutation left of S
Allele of S suppresible only by SuIII Amber in lysis gene R
Heat-sensitive allele of R
FRACKMAN et al. (1984) KOBAYASHI et al. (1982)
CAMPBELL (1961) J. S. PARKINSON
BROWN and ARBER (1964)
CAMPBELL (1961)
S z m m s ~ and Szmms~l (1979) DAVIS and PARKINSON (1971)
CAMERON et al. (1975) DAVIS and PARKINSON (1971)
KOBAYASHI et al. (1982)
CAMERON et al. (1975)
HRADECNA and S Z ~ ~ S K I (1969) GINGERY and ECHOLS (1967)
G. R. SMITH, personal communication SIGNER and WEIL (1968)
ZISSLER et al. (1971)
KAISER and JACOB (1957) CAMPBELL (1971)
SUSSMAN and JACOB (1962) MESELSON (1964)
STAHL et al. (1975) COURT and SATO (1969) HOLLIFIELD et al. (1987) HOLLIFIELD et al. (1987) HOLLIFIELD et al. (1987) STAHL et al. (1975)
GOLDBERG and HOWE (1969) CAMPBELL (1961)
BROWN and ARBER (1964) Elements are read from left to right on conventional map.
ried the plasmid than was observed when X rap' was used. This observation suggests that contributions by
X to the RecBCD pathway alter the properties of that pathway. We report herein that plasmid pickup by X
in the absence of Rap function manifests clustering of exchanges like that seen with rap' phage. Surprisingly, rap and rap' phage pick up apparent patches from plas- mids equally well, or nearly so (see APPENDIX). This observation argues that apparent patches are not simply close double splices.
Other results reported herein provide additional evi- dence against the idea that an appreciable fraction of marker pickups (apparent patches) from a plasmid into X is due to double splices. In so doing, they support the concept of (true) patches catalyzed by the RecBCD pathway. That in turn strengthens the view that both patches and splices, arising in a reciprocal interaction between X and a plasmid, are accompanied by a X X X
splice nearby to the right ( STAHL et al. 1990b).
MATERIALS AND METHODS
Phage genetic elements are described in Table 1. Bacterial strains and plasmids are in Table 2. Some stocks of phage
strains carrying large deletions were prepared on the r e d re&
strain FS3578 to reduce the frequency of addition mutants in the stock. In most experiments, unadsorbed phage were "eliminated" from the lysates as follows: parental phage stocks were prepared on hosts that do not modify DNA for the K restriction/modification system; host cells for the crosses were competent for modification but not for restric- tion; yields from the crosses were plated on hosts that are competent for restriction. Other conditions of crosses and other methods are those described in STAHL et al. ( 1990b). Some crosses, as indicated, are blocked for DNA replication.
RESULTS
Plasmid pickup: Pickup opposite a deletion: Among re- combinants for a distant pair of X markers, those that have picked up a plasmid are more likely than average to have enjoyed the X X X splice in the interval that contains the bit of DNA homologous with part of the plasmid ( STAHL et al. 1990b)
.
As
described in INTRO- DUCTION, not all of the recombinants that had picked1112 F. W. Stahl et al.
TABLE 2
Bacterial strains and plasmids
Strain Relevant properties Source/Reference
594 C600 AC417 MB406 Q358 V66 FS2152 FS2258 FS2942 FS2987 FS3127 FS3137 FS3238 FS3239 FS3546 FS3578 pLTl6 pLT18
pRKl0l PCP 175
PK.39
pK40
Su- rec+
Su+ rec+ Su- recD
Suf recB2I recC22 sbcBI5 hflA hflB rec+ r- m+
Su- recF143
Su- redl1009 hflA:Tn5 594 r- m+
V66[pLT16]
Su- redl ( A Jsus6 imm434 Rsus5)
V66 [ pLT 1 81 594[pCP175] Q358[pK391 Q358 [ pK401 FS2258 [pK40] Su+ recA redl r- m-
Kpnl shrink of pRKl0l (7.2 kb)
Shrunken pBR322 (AmpR, Tets) carrying A EcoRI-B fragment
Shrunken pBR322 carrying A from bp 16,121-22,346 (8.7 kb) EcoRI-Hind111 fragment of pMB9 with 1092 bp of AcI
containing a Ser to Ile mutation at amino acid position 77. Total length, 6.1 kb.
A fragment AatII(bp14978)-NcoI(bp19329) into corresponding
sites of pMTL22 (6.8 kb)
Inverse of pK.39 (7.4 kb)
WEICLE (1966)
APPLEYARD (1954)
A. CHAUDHURY
W. R. DOVE D. T. DENHARDT
CHAUDHURY and SMITH (1984) This paper
Collection This paper a
This paper This paper This paper This paper This paper This paper This paper This paper This paper
R. D. KOLODNER R. SAUER
A. KUZMINOV
dn& transductant of a V66 dnuBts22 [pLTlG] .
grammed in Figure 5a. However, it remains possible that some of the pickups are because of coupled nonre- ciprocal splices and that these pickups account for the shift of A X A exchanges into the pickup interval. This possibility can be tested by a A X plasmid X A cross in which only one of the two A parents has homology with the plasmid (Figure 6 ) .
The cross has several features, they are: ( 1 ) A parents are marked with terminal, counterselectable makers Asus and &us, respectively;
( 2 )
the A parents have het- erospecific cos sites and terminase genes, one of which (the Asus) is mutant. This assures that packaging (and recombination ) is initiated on only the 2+ ( homologue of A + ) parent ( HOHN 1975; FRACKMAN et al. 1984) ; ( 3 )one parent is deleted for the segment of A carried by the plasmid. In a control cross, neither parent is deleted for that A segment; and (4) one parent carries a CI marker, which renders the plaques clear in contrast with the turbidity of
CI
+ plaques.The data and further details for the two crosses are in Figure
7.
Figure7,
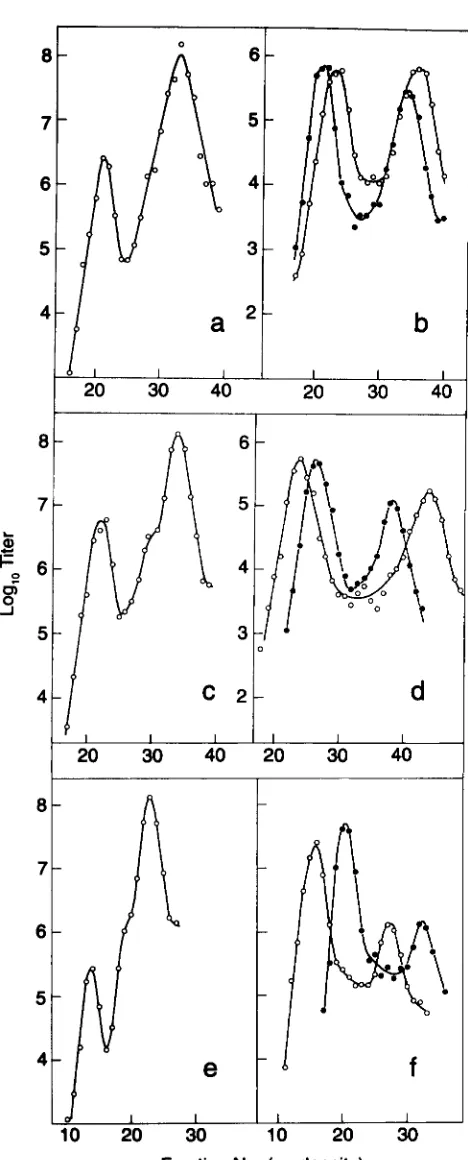
a and b, shows independent execu- tions of the control experiment, in which both parents have homology with the plasmid. Figure 7c shows the results of the cross in which only the Rsus5 parent has homology with the plasmid. In each figure, clear andturbid phage show three major peaks. Our conclusions derive from the behaviors of the heaviest and the light- est peaks. In those peaks, the relative exchange rates in the two intervals b519-
CI
and cI- R are given by the relative frequencies of the turbid and clear plaques, respectively. Estimating the frequencies of turbid phages from the heights of those two peaks gives the following results: for the controls, the frequencies of turbid plaques in the light peaks, in which the plasmid has not been picked up, are 3 and2%,
respectively, whereas the corresponding pickup peaks show 23 and 12%, respectively. This result is like that reported ear- lier for a similar cross ( STAHL et al. 1990b). For the experiment, in which the Asus parent has no homology with the plasmid, the frequencies of turbid plaques in the light and heavy peaks are 6 and 2396, respectively, similar to those seen in the control crosses.These results rule out the possibility that a major part of the shift of recombination into the A-cI interval accompanying the pickup of plasmid pLT18 is because of coupled nonreciprocal exchanges as diagrammed in Figure 5a.
Clustered Exchanges 1113 RecBCD by
a
b
CI+
+
0
pLT18FIGURE 6.-Plasmid pickup opposite a deletion in a A X plasmid X A cross. A Asusll was crossed with A Rsus5 in the presence of the plasmid pLT18. In cross a, both parents have homology with the EcoRI-B segment of A carried by the plas- mid. In cross b, the Asusll parent is missing the entire EcoRI- B segment (AB)
.
The crosses have the following additional features: ( 1 ) the Rsus5 parent carries cos and terminase genes from phage 21. The crosses are executed in an Su- host (FS3127), so packaging and recombination are initiated only on the Rsus5 parent. ( 2 ) The host is mutant in r e 8 to main- tain the plasmid in a monomer state (verified by gel elec- trophoresis of extracted DNA, data not shown). ( 3 ) Both parents are deleted for red and gam (Aspi6) so that recombi- nation can proceed by the RecBCD pathway. This deletion and the deletion Ab519 in the &us5 parent shorten the Achromosome so that phage chromosomes that have picked up the plasmid are packagable. The presence of Ab519 only in the Rsus5 parent ensures that any (unexpected) pickups in the Asus parent will tend to be lost from the analysis. The deletion Aspi6 invades cIII, rendering the plaques clear on ordinary hosts. (4) 2+ R+ recombinants were assayed on E. coli strain FS2152, which is Su- (to select sus+ recombinants), re& (to allow Aspi6 phage to make large plaques) , and hfZA
(to permit A cIII to make turbid plaques)
.
2’ R + cIII recombi- nants carrying the marker cI26make clear plaques, which are scored as recombinants in the cI-R interval. Turbid plaques formed on FS2152 are 2+R+ recombinants resulting from exchange in the A-cI interval. The Rsw parent carries x + D , which results in a relative “shortening” of the A-cI interval, making any increase in that interval associated with plasmid pickup more easily detected ( STAHL et al. 1990b).occur in a single, triparental event-among phage that had picked up the plasmid and had undergone A X A recombination, essentially all of the exchanges between the A chromosomes occurred at or to the right of the point at which the plasmid was incorporated. To distin- guish whether the A X A exchanges are at the point of pickup or to the right of that point, we performed experiments comparable to the Adv pickup experi- ments, using a plasmid (pCP175) that carries the cI gene of phage A, just left of 0-0.
The phages were rendered Red- Gam- by the substi- tution biol, which is a net deletion of about 5% of the A chromdsome. Additional DNA was removed by the deletion b2, making room for the plasmid. One parent
was marked with Jsusb and the other with Rsus5. The
&us5 parent was always imm‘. The Jsus6 parent was
either imm‘ or carried in its place the heterologous substitution imm434. Because the cI gene of A is entirely replaced by the imm434 substitution, homologous recom- bination between the plasmid pCP175 and A imm434
does not occur. Additionally, the crosses carried
x
+ C,located just to the right of imm434. The crosses are dia- gramed in Figure 8, and the results of the crosses, all carried out on the same culture, are in Figure 9.
In Figure 9A, both parents are imm‘, so both share homology with the plasmid. Among
J’
R+ recombi- nants, the fraction of phage that have picked up the plasmid is 2.0x
When the Jsusb parent is imm434(Figure 9B), the fraction of J’ R’ that has picked up the plasmid is
2.4
X lO-’l. Thus, among J’ R+ recombinants, the fraction that has picked up plasmid was not detectably diminished by eliminating the possi- bility of pickup via a pair of nonreciprocal exchanges.A second comparison supports that conclusion. When both parents are imm‘, the fraction of the
J’
x
+ C R+ phage that has picked up the plasmid is 6.7 Xl O P 3 l . When the Jsusb parent is imm434, the fraction of J’ imm‘ R’ phage, almost all of which are x + C , that has picked up plasmid is 1.3 X lo-*.
The primary conclusion of the crosses is that heterol- ogy in the
x o
parent does not diminish the fraction of J’ R’ A that has picked up the plasmid. Thus, again, the major route of plasmid pickup cannot be the one involving a pair of nonreciprocal A X plasmid ex- changes. Instead, the plasmid must be incorporated by a single reciprocal exchange. A caveat to that conclu- sion is detailed in DISCUSSION.Marker pickup: Marker pickup from a plasmid may be the result of patch formation. Such patches may represent the alternate resolution of an intermediate that can result in splices. The intermediates given most currency have been those of HOLLIDAY (1964), MESEL
SON and RADDING ( 1975) and SZOSTAK et al. ( 1983)
.
Operationally, the distinction between a patch and a pair of close splices is difficult. When the hypothesized pair of splices involves three chromosomes, however, there is no ambiguity.STAHL et al. (1990b) demonstrated that pickup of a marker from a plasmid into X (an apparent patch) is accompanied by a nearby A X A splice. Such a correla- tion can be variously interpreted. The following two
interpretations strike us as the most simple: (1) A has picked up a true patch from a plasmid. This event is accompanied by a nearby, but separate, A X A splice (Figure 1OA). ( 2 ) A nonreciprocal splice between A and plasmid is followed by degradation of the plasmid. The degradation is halted by a second nonreciprocal splice, involving a second A chromosome (Figure l o b ) .
1114 F. W. Stahl et al.
6
L
5
i= d
E 4
0 )
5 3
2
30 40 20 30 40
50
60 iFraction No. (+density)
FIGURE 7.-Equilibrium density gradient distributions of progenies from crosses described in Figure 6. In each gradient, the heaviest peak is comprised of phage particles that have picked up the plasmid, pLT18, and have the genotype cosz1 Ab519Aspi6
R + . In a and b, the lightest peak is comprised of particles of that same genotype except that they lack the plasmid; also in a and b, which are replicates from the control cross diagramed in Figure 6a, the peaks of intermediate density have a more complex origin. The clear plaques ( 0 ) arise from phenotypically dense particles of the predominant genotype in the light peak, cosz1 Ab519Aspi6 cI26 R + . These particles are demonstrably dense as a result of having the immature capsids that characterize a fraction of A particles that have packaged short chromosomes ( STERNBERG and WEISBERG 1977). Particles picked from plaques made by the clear phage in this peak of intermediate density band with the density characteristic of the typical cosz1 Ab519 Aspi6 particles. The turbid plaques ( 0 ) are due primarily to recombinant phage of genotype cosz1 Aspi6 cI+ R + . In c, which corresponds to the cross in Figure 6b, both the clear and turbid phage of intermediate density are phenotypically dense variants of the phages in the major light peak. In c, the light peak of turbid phage has a light shoulder of the recombinant genotype
cosz1 AB Aspi6 cI+ R + .
a
b
>
J~~~ b2 biof Cl++
@
PCP175>
+
b2 b i 0 l cl'X+c
Rsus>
FIGURE 8.-Plasmid pickup opposite a substitution in a A
X plasmid X A cross. A Jsus6 was crossed with A Rrus5 in the presence of the plasmid pCP175, which carries the cI gene of A. The lysates were centrifuged in CsFormate, and J + R+
recombinants were plated selectively. Each phage was Int- Red- Gam- by virtue of the substitution biol. Additional space for the plasmid was provided by the deletion 62. In one cross ( a ) both parents were homologous with the lasmid. In the other cross ( b ) , the Jsusbparent carried imm4", a substitution that only rarely recombines with immk. The Rrus5 parent car- ried x + C , at base pair 38,483, which is 238 bp rightward of the right end of imm434.
x
+ C stimulates pickup of pCP175, allowing its quantification by density gradient centrifugation (Figure 9 ) .30 40
C
-
4050
Fraction No. (t density)
FIGURE 9.-Equilibrium density gradient distributions of progenies from crosses described in Figure 8. The crosses diagrammed in Figure 8 were executed in bacterial strain FS3137. In a, density fractions were plated on AC417, a Su- re& host that optimizes the distinction between clear ( 0 )
Clustered Exchanges by RecBCD 1115
a
b
m2
(rll
+
1
- ) 2
I
2
1
1
- )2
if the second A chromosome (Figure 8 b ) lacks the ho- mology that permits recombination between the plas- mid and the first X. This loss would be detected as a reduction in the frequency of apparent patches among A progeny particles recombinant for markers flanking the marker pickup point.
To assess the possibility that a large fraction of the marker pickups in A X plasmid X A crosses arise by the second route, we compared the outcomes of two crosses. The plasmid pCPl75, which is too large to be found spliced into full-length progeny X particles, car- ries a clear mutation in the cloned A
GI
gene. The J’ parent was immA and carried x + D , located near the right end of the J- R interval.x
+ D can stimulate recom-bination in the cI gene
(LAM
et al. 1974) . The presence of x + D ensures that most of the detected marker pick- ups are the result of events initiated on the J’ parent. In one cross, the R+ parent was also immA; in the other cross, the R’ parent carried the heterologous substitu- tion imm434 (Figure 11 ).
The results of the two crossesFIGURE 10.-Two ways in which marker pickup from a plasmid could stimulate nearby A X X exchange. In a, X-stimulated patches are swapped reciprocally between
A and a plasmid. A (nonreciprocal) ex- change to the right results in a complete A
recombinant chromosome. In b, a pair of
(nonreciprocal) splices occurs within the
A segment carried by the plasmid. The dia- gram is noninformative regarding the het- eroduplex nature of the patches and splices, which may be detected or not de- pending on the method of analysis.
(Table 3 ) reveal an equal frequency of clear plaques among J’ R+ recombinants whether the R parent was immA or imm434. Thus, no large fraction of the marker pickups among J’ R+ recombinants can be the result of a pair of close, triparental splices. This conclusion is congruent with the conclusion by STAHL et al. ( 1984) that
x
“gives” and “takes” patches equally, which im- plies thatx
stimulates reciprocal patching.X’s rap function: A’s gene rap is known to influence RecBCD-mediated recombination between A and a plas- mid with which X shares homology (HOLLIFIELD et al. 1987). We examined plasmid pickup by a A rap mutant to see whether the clustering of exchanges seen with rap’ phage was dependent on Rap function.
We crossed A phages marked at M, cI and R in the presence of plasmid pK40, which contains a wild-type J gene. Both the A phages in the cross carried a ts mutation in J (Figure 1 2 ) . In one cross, the phages were Rap’, in the other they were Rap ~ by virtue of the d 5 deletion.
F. W. Stahl et al.
1116
a
b
>
+
int red gam cl+ x+DRtsFIGURE 11.-Marker pickup in A X plasmid X A crosses. int4 red3gamZlOderivatives of A Jtsl5 and of A x + D RtsZwere
crossed in strain FS3137, which carries plasmid pCP175. This plasmid carries a mutant allele of the cI gene of A. In one cross, both parents were imm” and hence were homologous with the A segment carried by the plasmid. In the other cross,
the Jts R + parent carried the heterologous substitution i m ~ ~ ~ ’ ~ .
that had picked up the plasmid from those that had not. In accord with expectation ( HOLLIFIELD et al. 1987), the pickup of plasmid into A (splicing) was reduced by the rap mutation. The reduction in pickup was 17-fold. Patching of the J’ marker into A was reduced 2.5-fold. This differential effect of rap on splices and patches in A X plasmid recombination is further documented in the APPENDIX ( see Figure A2)
.
We plated the gradient fractions for M + R+ recombi- nants at low temperature, which enumerated all A X A recombinants, and we plated for MiJi Ri recombi- nants at high temperature, which identified those A X A recombinants that were J’ by virtue of recombination with the plasmid that resulted in either a splice (heavy peak) or a patch (light peak)
.
The M’R’ recombi- nants in each peak at each temperature were scored for the cI marker. The data (Table 4 ) show that A X A exchange is increased in the M-cI interval by both the splicing and the patching of J’ from the plasmid into the A and that the effect is similar in degree for the Rap - and the Rap+ crosses.We make two conclusions from the Rap experiments. The properties of RecBCD recombination described herein are not qualitatively dependent on X Rap func- tion, By that token, they are more apt to characterize the RecBCD pathway uninfluenced by A gene functions. Second, the differential reduction in splices and patches resulting from the rap mutation augments the evidence that “patches” in the RecBCD pathway are not simply the result of closely spaced double splices.
DISCUSSION
STAHL et al. ( 1990b) demonstrated a relationship be- tween plasmid pickup and exchange between two A
TABLE 3
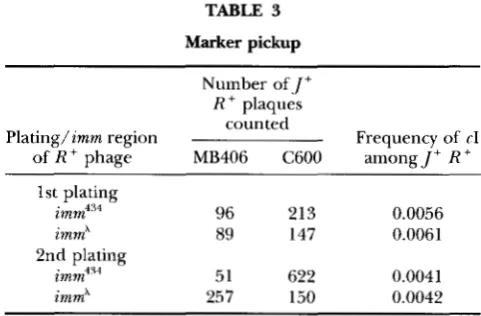
Marker pickup
Number of, J’
R + plaques
counted
of R + phage MB406 C600 among J’ R
’
Plating/imm region Frequency of rI1st plating
imm434 13 96 2
0.0056
immA 89 147 0.0061
2nd plating
imm434 51 622 0.0041
imm” 150 0 257
.0042
After appropriate dilution of the cross lysate, total .J+R+ recombinants were assayed twice on strain C600 at 42”,
whereas J’ R + recombinants that have picked up the cI
marker from the plasmid were assayed at 42” on the hfl strain MB406, upon which A cIi particles do not make plaques.
phage chromosomes recombining by the RecBCD path- way. When a plasmid is picked up by A and particles are selected for having enjoyed a A X A exchange event, the exchange is almost always at or to the right of the site of plasmid pickup. By this criterion, plasmid pickup and X X X exchange occur “in the same act.” Results presented here, in which one A parent is nonhomolo- gous with the plasmid, argue that the A X A exchange rarely occurs at the site of the plasmid pickup. This demonstration strengthens the view that plasmid pickup reflects a single reciprocal exchange event. Ever more complex pathways for the origin of any recombi- nant type in any genetic system can always be proposed. An example is presented later in this DISCUSSION. Thus, a proof of reciprocality satisfying to all is probably unat- tainable. Similarly, our results show that marker pickup, which is also frequently accompanied by A X X ex- change ( STAHL et al. 1990b) , is rarely, if ever, the result of a pair of splices involving two different phage chro- mosomes. That demonstration strengthens the case for interpreting marker pickup from a plasmid into A as representing (reciprocal) patch formation. HAGEMANN
and ROSENBERG ( 1991 ) argued for patches, as opposed to closely paired splices, in RecBCD-mediated A X plas- mid crosses. Our observation (APPENDIX) that a rap mu- tation reduces plasmid pickup by a greater factor than it does marker pickup similarly argues that patches in the RecBCD pathway are not the consequence of clus- tered splices.
Clustered Exchanges RecBCD by 1117
Msus J t s b 2 biol cl ( d s )
+
FIGURE 12.-Plasmid and marker pickup in a Rap- cross. The indicated phages were crossed in the presence of pK40 in the host FS3239. In one cross, both parents were nin’; in
the other both carried the Rap- deletion d5. The phages are Int- Red- Gam- by virtue of the substitution biol. That substitution is a net 5% deletion. When biol is combined with the deletion b2, the phages carry a net deletion of 17%, which provides enough space to pick up the plasmid. The plaque- morphology marker cI26, in the Msus parent, divides the M- R interval in two. Because biol has a leftward
x
sequence, we addedx
+ D to the Rsus parent to assure that comparablenumbers of M + R+ recombinants would arise in the two inter- vals. Lysates resulting from such crosses were centrifuged in cesium formate to separate particles that have picked up the plasmid from those that have not. The M + S R+ recombinants in each of the two peaks were scored for cI. Clear plaques are from recombinants in the M cZ internal.
and KUPIEC (1994) for yeast and reported herein for the RecBCD pathway of E. coli, leave no doubt as to the complexity of some recombination events. No single, simple model ( HOLLIDAY 1964; MESELSON and RADDINC
1975; SZOSTAK et al. 1983) can cope.
For the RecBCD pathway, we entertain two mecha- nisms to explain the triparental clusters seen here and in STAHL et al. (1990b). First, the bimolecular recipro- cal exchange event, which accounts for plasmid pickup and for the equality in complementary types in a X X X cross when the
x
+ parent is in excess, could be verylike the pathway proposed by HOLLIDAY ( 1964). In its present incarnation, we envision a RecAdependent four-chained structure ( CONLEY and WEST 1989) that can be acted upon by an endonuclease activity of a travelling RecBC ( D - ) enzyme to produce a Holliday junction. Resolution of the junction, by a “resolvase” that could be RecBC ( D - )
,
completes the reciprocal exchange event (or produces a pair of patches). The nonreciprocal, coupled event, which leads to the re- quirement for twox
+ parent chromosomes to produceone
x
+ recombinant, could be like that proposed fora number of systems, including the Red pathway of X ( STAHL et al. 1985; SIDDIQI et al. 1991 )
.
In particular, single chained DNA generated by the unwinding activ- ity of the RecBCD protein becomes coated with RecA protein and initiates a 3“ended invasion of an intact duplex (SMITH et dl. 1984; SIDDIQI et al. 1991 ).
Appro- priate nuclease and ligase activities complete the nonre- ciprocal break-join reaction ( STAHL et al. 1990a). Ro-MAN et al. ( 1991 ) offer evidence for such events arising from the joint in vitro activities of RecBCD and RecA proteins.
Our second model is derived from that of ROSENBERC
TABLE 4
Rap and reciprocality
cI marker among selected Rap type/ genotype*
selected
genotype Peak C C+ Mottledt
b P +
M + R + Light 513 (52) 466 (47) 20 (2)
M’J’ R + Light 63 (63) 33 (33) 4 (4) Heavy 142 (71) 58 (29)
Heavy 75 (75) 25 (25) Rap-
M + R + Light 594 (37) 929 (57) 62 (4)
M’J’ R + Light 114 (57) 72 (36) 14 (7)
Heavy 100 (67) 50 (33)
Heavy 72 (72) 28 (28)
*Values are no. of plaques examined with percentage in
Among phage that have picked up the plasmid, hetero- parentheses.
duplexes at the cI locus were not scored.
and HASTINGS (1991). This model supposes that
RecBCD converted to RecBC ( D - ) by its encounter with
x
retains helicase activity. As the altered enzyme continues traveling leftward it “splits” the end of the duplex ( RINKEN et al. 1992) and each of the resulting single chains invades a different homologue. The 3’-ended chain behaves as described above. The 5‘ended chain engages a homologue somewhere to the left and provokes a reciprocal event, either patching or splicing
(and see MYERS and STAHL 1994).
The reciprocality claimed herein for the event that incorporates a plasmid into X is, by the nature of the experiments, “genetic” reciprocality. At the molecular level, a pair of nonreciprocal events involving sister du- plexes, as in the the unisex circle model ( STAHL 1979), could underlie this reciprocality.
We considered the possibility that our results on re- combination between X and a plasmid via the RecBCD pathway were determined by the involvement of X’s rap gene ( HOLLIFIELD et al. 1987)
.
In agreement with those authors, we found that the loss ofRap
function im- paired the ability of X to pick up a plasmid with which it has homology. Surprisingly (see APPENDIX),
this im- paired ability is not simply because of a reduced overall rate of recombination, because the ability of X to pick up patches of information from a plasmid is relatively less impaired by the rap deletion. In rap mutants, the association of X XX
recombination withX
X plasmid recombination was the same as in rap’ X. Although that result gives little insight into the role of Rap function, it does increase the likelihood that our results on clus- tered exchange apply to the bacterial RecBCD pathway as well as to a X-modified RecBCD pathway.1118 F. W. Stahl et al.
duplex regions flanking a plasmid picked up into rap’
A
by the action of the RecBCD pathway are not precise complements of each other. This nonreciprocality at the microscopic level does not contradict our conclu- sion that plasmid pickup is reciprocal at the macro- scopic level.Numerous workers have demonstrated reciprocality in the RecBCD pathway operating on the E. coli chromo- some (e.g., HERMAN 1965, 1968; MESELSON 1967; hlA
HAN and ROTH 1989) Our demonstration that a single
pass by RecBCD can catalyze both a reciprocal and a nonreciprocal exchange suggests a framework of thought for those observations. KUZMINOV et al. (1994) proposed that RecBCD can enter DNA at a double- chain break arising in an arm of a replication fork. Digestion of that arm is halted by
x,
and the “frayed end” invades its sister, reestablishing the broken fork. In the meanwhile, the X-activated enzyme, traveling within the originally broken arm, can catalyze a recipro- cal exchange between that arm and a homologue. This view has significant common elements with the pro- posal of MAHAN and ROTH ( 1989).
ROBERT SAUER and FLORA BANUETT kindly provided strains. This work was supported by National Science Foundation grant DMB- 8905310 and National Institute of General Medical Sciences grant GM-33677. F.W.S. is American Cancer Society Research Professor of Molecular Genetics.
Note added in prooj For rap mutant phage, patches are lowered as much as splices (about 20-fold) when DNA replication is blocked.
LITERATURE CITED
AMUNDSEN, S. K., A. F. TAYLOR, A. M. CHAUDHURY and G. R. SMITH, 1986 r e d , the gene for an essential third subunit of exonuclease V. Proc. Natl. Acad. Sci. USA 83: 5558-5562.
APPLEYARD, R. K., 1954 Segregation of new lysogenic types during growth of a doubly lysogenic strain derived from Escherichia coli K12. Genetics 39: 440-452.
BROWN, A,, and W. ARBER, 1964 Temperature-sensitive mutants of coliphage lambda. Virology 2 4 237-239.
CAMERON, J., R. S. PANSENKO, I. R. LEHMAN and R. W. DAVIS, 1975
In vitro construction of bacteriophage A carrying segments of the Eschmkhia coli chromosome: selection of hybrids containing the gene for DNA ligase. Proc. Natl. Acad. Sci. USA 72: 3416-3420. CAMPBELI., A,, 1961 Sensitive mutants of bacteriophage lambda. Virol-
ogy 1 4 22-32.
CAMPBEI.I., A,, 1971 Genetic structure, pp. 13-14 in TheBactm’qphage Lambda, edited by A. D. HERSHEY. Cold Spring Harbor Labora- tory, Cold Spring Harbor, N Y .
CHAUDHURY, A. M., and G. R. SMITH, 1984 A new class of Eschachia coli recBC mutants: implications for the role of RecBC enzyme in homologous recombination. Proc. Natl. Acad. Sci. USA 81:
CONLEY, E. C., and S. C. WEST, 1989 Homologous pairing and the formation of nascent synaptic intermediates between regions of duplex DNA by RecA protein. Cell 56: 987-995.
COURT, D., and K. SATO, 1969 Studies of novel transducing variants of lambda: dispensability of genes N a n d Q. Virology 39: 348-
352.
DAVIS, R. W., and J. S. PARKINSON, 1971 Deletion mutants of bacterio- 7850-7854.
phage lambda. 111. Physical structure of at@. J. Mol. Biol. 56: 403-423.
FRACKMAN, S., D. A. SIEGELE and M. FEISS, 1984 A functional domain of bacteriophage A terminase for prohead binding. J. Mol. Biol. 180: 283-300.
GINGERY, R., and H. ECHOLS, 1967 Mutants of bacteriophage lambda unable to integrate into the host chromosome. Proc. Natl. Acad. Sci. USA 58: 1507-1514.
GOLDBERG, A. R., and M. HOW, 1969 New mutations in the Scistron of bacteriophage lambda affecting host cell lysis. Virology 38: 200-202.
HAGEMANN, A. T., and S. M. ROSENBERG, 1991 Chain-bias in Chi- stimulated heteroduplex patches in the A ren gene is determined by the orientation of A cos. Genetics 129: 611-621.
HERMAN, R. K., 1965 Reciprocal recombination of chromosome and F-merogenote in Escherichia coli. J. Bacteriol. 90: 1664-1668. HERMAN, R. IC, 1968 Identification of recombinant chromosomes
and F-merogenotes in merodiploids of Escherichia coli. J. Bacte- riol. 96: 173-179.
HOHN, B., 1975 DNA as a substrate for packaging into bacteriophage lambda in vitro. J. Mol. Biol. 98: 93-106.
HOLLIDAY, R., 1964 A mechanism for gene conversion in fungi. Genet. Res. 5 282-304.
HOLLIFIELD, W. C., 1988 Tetranemic analysis: the equivalent of tetrad analysis in Escherichia coli applied to RecABGmediated phage- plasmid recombination. Ph.D. Thesis, Washington University, St. Louis, MO.
HOLLIFIELD, W. C., E. N. -LAN and H. V. HUANG, 1987 Efficient RecABCdependent homologous recombination between coli- phage lambda and plasmids requires a phage ninR region gene. Mol. Gen. Genet. 210: 248-255.
HRADECNA, Z., and W. S Z ~ ~ S K I , 1969 Electron micrographic maps of deletions and substitutions in the genomes of transducing coliphages Adg and Abio. Virology 38: 473-477.
KAISER, A. D., and F. JACOB, 1957 Recombination between related temperate bacteriophages and the genetic control of immunity and prophage localization. Virology 4: 509-521.
KOBAYASHI, I., H. MURIALDO, J. M. CRASEMANN, M. M. STAHI. and F. W. STAHL, 1982 Orientation of cohesive end site cos deter- mines active orientation of Chi sequence in stimulating r e d - recBC-mediated recombination in phage A lytic infections. Proc. Natl. Acad. Sci. USA 79: 5981-5985.
KOBAYASHI, I., M. M. STAHL, D. LEACH and F. W. STAHL, 1983 The interaction of cos with Chi is separable from DNA packaging in recA-recBC-mediated recombination of bacteriophage lambda. Genetics 104 549-570.
KOBAYASHI, I., M. M. STAHI., F. R. FAIRFIELD and F. W. STAHL, 1984 Coupling with packaging explains apparent nonreciprocality of Chi-stimulated recombination of bacteriophage lambda by R e d and RecBC functions. Genetics 108: 773-794.
KUSANO, IC, Y. SUNOHARA, N. TAKAHASHI, H. YOSHIKURA and I. KOBAY- ASHI, 1994 DNA double-strand break repair: genetic determi- nants of flanking crossing-over. Proc. Natl. Acad. Sci. USA 91: 1173-1177.
KUZMINOV, A,, E. SCHABTACH and F. W. STAHI., 1994 How E. coli uses RecBCD nuclease both for the degradation of foreign DNA and for the repair of chromosomal DNA. EMBO J. 13: 2764-2776. LAM, S. T., M. M. STAHL, K. D. MCMIIJN and F. W. STAHI., 1974
Rec-mediated recombinational hot spot activity in bacteriophage lambda. 11. A mutation which causes hot spot activity. Genetics
W A N , M. J., and J. R. ROTH, 1989 Role of recBCfunction in forma- tion of chromosomal rearrangements: a two-step model for re- combination. Genetics 121: 433-443.
MESELSON, M. S., 1964 On the mechanism of genetic recombination between DNA molecules. J. Mol. Biol. 9: 734-745.
MESELSON, M. S., 1967 Reciprocal recombination in prophage lambda. J. Cell. Physiol. 1 (Suppl.) : 113-118.
MESELSON, M. S. and C. M. RADDING, 1975 A general model for genetic recombination. Proc. Natl. Acad. Sci. USA 72: 358-361. MYERS, R., and F. W. STAHI., 1994 Chi and the RecBCD enzyme of
Eschm’chia coli. Annu. Rev. Genet. 28: 49-70.
RAY, A,, N. MACHIN and F. W. STAHL, 1989 A DNA double chain break
Clustered Exchanges by RecBCD 1119
stimulates triparental recombination in Saccharomyces cereuisiae. Proc. Natl. Acad. Sci. USA 8 6 6225-6229.
RINKEN, R., B. THOMS and W. WACKERNAGEL, 1992 Evidence that recBCdependent degradation of duplex DNA in E s c h c h i a coli redl mutants involves DNA unwinding. J. Bact. 174: 5424-5429. ROMAN, L. J., D. A. DIXON and S. C. KOWALCZYKOWSKI, 1991 RecBCD- dependentjoint molecule formation promoted by the Escherichia coli RecA and SSB proteins. Proc. Natl. Acad. Sci. USA 88: 3367- 3371.
ROSENBERG, S. M., 1987 Chistimulated patches are heteroduplex, with recombinant information on the phage A r chain. Cell 48:
ROSENBERG, S. M., 1988 Chain-bias of Escherichia coli Rec-mediated A patch recombinants is independent of the orientation of A cos.
Genetics 1 1 9 7-21.
ROSENBERG, S. M., and P. J. HASTINGS, 1991 The splitend model for homologous recombination at double-strand breaks and at Chi. Biochimie 73: 385-397.
SANG, H., and H. L. K. WHITEHOUSE, 1983 Genetic recombination at the buff spore color locus in So-rdaria brm'collis. 11. Analysis of flanking marker behavior in crosses between buff mutants. Genetics 103: 161-178.
SAWITZKE, J. A,, and F. W. STAHL, 1992 Phage A has an analog of Escherichia coli rec0, recR and recFgenes. Genetics 1 3 0 7-16. SIDDIQI, I., M. M. STAHL and F. W. STAHL, 1991 Heteroduplex chain
polarity in recombination of phage A by the Red, RecBCD, RecBC ( D - ) and RecF pathways. Genetics 128: 7-22.
SIGNER, E. R., and J. WEIL, 1968 Recombination in bacteriophage A. I. Mutants deficient in general recombination. J. Mol. Biol. 34:
261-271.
SILBERMAN, R., and M. KUPIEC, 1994 Plasmid-mediated induction of recombination in yeast. Genetics 137: 41-48.
SMITH, G. R., S. K. AMUNDSEN, A. M. CHAUDHURY, K. C. CHENG, A. S. PONTICELLI et al., 1984 Roles of RecBC enzyme and Chi sites in homologous recombination. Cold Spr. Harbor Symp. Quant. Biol. 39: 485-496.
SMITH, G. R., and F. W. STAHL, 1985 Homologous recombination provided by Chi sites and RecBC enzyme of Escherichia coli. BioEs- says 2 244-249.
STAHL, F. W., 1979 Genetic Recombination, Thinking about it in Phage and Funga. W. H. Freeman, San Francisco, CA.
STAHI., F. W., and M. M. STAHL, 1985 Non-reciprocal crossing over in phage A. J. Genet. 64: 31-39.
STAHL, F. W., J. M. CRASEMANN and M. M. STAHL, 1975 Rec-mediated recombinational hot spot activity in bacteriophage A. 111. Chi mutations are site-mutations stimulating Rec-mediated recombi- nation. J. Mol. Biol. 9 4 203-212.
STAHI., F. W., M. M. STAHL, L. YOUNG and I. KOBAYASHI, 1982 Chi- stimulated recombination between phage A and the plasmid Adv. Genetics 102: 599-613.
STAHL, F. W., M. LIEB and M. M. STAHL, 1984 Does Chi give or take? Genetics 108: 795-808.
STAHL, F. W., I. KOBAYASHI and M. M. STAHL, 1985 In phage A, cos
is a recombinator in the Red pathway. J. Mol. Biol. 181: 199- 209.
STAHL, F. W., M. S. FOX, D. FAULDS and M. M. STAHL, 1990a Break- join recombination in phage A. Genetics 1 2 5 463-474. STAHL, F. W., L. C. THOMASON, I. SIDDIQI and M. M. STAHL, 1990b
Further tests of a recombination model in which
x
removes the RecD subunit from the RecBCD enzyme of Escherichia coli. Genetics 126: 519-533.STERNBERG, N., and R. WEISBERG, 1977 Packaging of coliphage lambda DNA. 11. The role of the gene D protein. J. Mol. Biol. 117: 733-759.
SUSSMAN, R., and F. JACOB, 1962 Sur un systeme de repression ther- mosensible chez le bacteriophage A d'Escha'chia coli. C. R. Acad. Sci. 254: 1517-1519.
SZOSTAKJ. W., T. L. ORR-WEAVER, R. J. ROTHSTEIN and F. W. STAHL, 1983 The double-strand-break repair model for recombination. Cell 33: 25-35.
SZkBALSKt, E. H., and W. SZYBALSKI, 1979 A comprehensive molecular map of bacteriophage lambda. Gene 7: 217-270.
THALER, D. S., M. M. STAHL and F. W. STAHL, 1987 Doublechain- 855-865.
cut sites are recombination hotspots in the Red pathway of phage A. J. Mol. Biol. 195 75-87.
THALER, D. S., E. SAMPSON, I. SIDDIQI, S. M. ROSENBERG, F. W. STAHL et al., 1988 A hypothesis: Chi-activation of RecBCD enzyme involves removal of the RecD subunit, pp. 413-422 in Mechanism and Comepaces o f D N A Damage Processing, edited by E. FRIEDBERG
and P. HANAWALT. Alan R. Liss, New York.
THALER, D. S., E. SAMPSON, I. SIDDIQI, S. M. ROSENBERG, L. C. THOM-
ASON et al., 1989 Recombination of bacteriophage A in recD nutants of Escherichia coli. Genome 31: 53-67.
WEIGLE, J., 1966 Assembly of phage lambda in vitro. Proc. Natl. Acad. Sci. USA 5 5 1462-1466.
WEIL, J., 1969 Reciprocal and non-reciprocal recombination in bacte- riophage A. J. Mol. Biol. 4 3 351-355.
ZISSLER, J., E. SIGNER and F. SCHAEFER, 1971 The role of recombina- tion in growth of bacteriophage lambda. I. The gamma gene, pp. 455-468 in The Bacteriophage Lambda, edited by A. D. HER-
SHEY. Cold Spring Harbor Laboratory, Cold Spring Harbor, N y .
Communicating editor: P. J. PUKKILA
APPENDIX: THE rap MUTATION OF PHAGE A
HOLLIFIELD et al. (1987) identified a gene in the nin5 region of A that facilitated recovery of cointegrates arising by homologous recombination between A and a plasmid. They called the gene rap for recombination adept with plasmids.
We confirmed the observation of HOLLIFIELD et al. for our strains and conditions. Our rap mutation was the deletion d 5 . To control for the consequences of the (slightly) reduced size of A's chromosome, we used two rap' phages, one of which was wild type in the nin5 region and the other of which carried the nearby dele- tion d 2 , which is longer than d 5 ( HOLLIFIELD et al. 1987;
SAWITZKE and STAHL 1992). Our phage were A b 2 biol,
which renders them Red- Gam- and provides room to pick up one copy of the plasmid pLTl6. That plasmid is 7.2 kb in length and has homology to A left of the b2
deletion. We infected at room temperature a culture of
recBW+ bacteria (FS2942) carrying pLT16 separately with each of the three phages at a mutiplicity of seven. After 30 min to allow for adsorbtion, we filtered the infected cultures to remove unadsorbed phage. We re- suspended the infected cells recovered from the filter, aerated them at 37" and lysed them (egg white lysozyme and chloroform) after 75 min. We centrifuged the re- sulting lysates to equilibrium in a cesium formate gradi- ent to separate the phage particles according to their density, which is determined by their DNA content. Plaque-assay of fractions collected from each gradient revealed a large peak corresponding to A phage that had not picked up plasmid and a smaller, denser peak of cointegrate particles. The ratio of the peak heights in a gradient is a measure of the fraction of phage that have picked up plasmid. We found (Figure A1 ) that the nin+
and d 2 control lysates contained cointegrate particles at
1120
t
i
E
5
4
a
7
zl
2?
E 6
0
-I
5 4
8
7 6 5 4
F. W. Stahl et al.
0
6 -
5 -
4 -
3 -
d
20
30
4020
30
40c 2
r
I I I I I I
20
30 40 2030
40I I 1
10
20
30Fraction No. ( c density)
FIGURE A1.-Affect of Rap on cointegrate frequency and stability. A b2 biol was grown for one cycle in FS2942, which carries pLT16. Unadsorbed phage were assayed and found to be a negligible fraction of the total yield. Infecting phages varied according to whether they were nin’ ( a and b ) o r Ad2
( c and d ) or Ad5 ( e a n d f )
.
Resulting lysates were centri- fuged to equilibrium, and density-fractions were assayed forfrequencies of 0.02 and 0.04, respectively. The d 5 (Rap-) lysate, on the other hand, contained such parti- cles at a frequency of 0.002. This 10- to 20-fold effect of the rap mutation was reproducible.
The demonstrated Rap effect could mean either that the pickup of a plasmid is poor in rap mutants or that cointegrates involving A rap are less stable. To distin- guish these possibilities, we plated from the heavy gradi- ent peaks on strain C600. From each plating, we picked and pooled 20 young ( 6 h r ) plaques and spun the particles in cesium formate. Each plaque pool gave rise to two density peaks of phage corresponding to cointe- grates and to A particles from which the plasmid has been lost, respectively. The relative heights of the two peaks in a gradient is a measure of stability of the cointe- grate. We found (Figure A1 ) that the Rap - cointegrate
was more, not less, stable than the nin+ and d 2 cointe- grates. We repeated the procedure to demonstrate re- producibility (Figure A1 )
.
Our result implies that the loss of this plasmid from a cointegrate is reduced by the rapdeletion, as is its original pickup. Thus, hypothe- sized instability of the Rap- cointegrate cannot account for the primary Rap phenotype. Following a suggestion by DAVID S. THALER (personal communication), we asked whether a rap mutation reduced the ability of A to pick up a marker from a plasmid ( a patch) as it did the ability to pick up the entire plasmid ( a splice).
We made two plasmids (pK39 and pK40) that carry A’s J gene in opposite orientations (total size, 6808 bp).
We separately infected hosts carrying these plasmids with A A d5,
Ad 2 or n in+. Each phage was A b2 bio and carried an amber mutation ( s u s 6 ) in J. We distinguished plas- mid pickups from marker pickups by centrifuging the lysates in a cesium formate density gradient and plaque- assaying relevant fractions from the gradient. Results with the two kinds of plasmids were entirely equivalent (Figure A2).
They show, as before, that Rap - phagehave impaired ability (
-
1OX ) to pick up plasmid. How- ever, the ability of the Rap- phage to patch a marker from the plasmid to A was reduced only about twofold.HOLLIFIELD et al. (1987) reported that Rap is specific to the RecBCD pathway, as measured by the frequency of cointegrates formed between A and a plasmid. We have confirmed that conclusion by showing a lack of rap dependence for cointegrate formation in A’s Red