Copyright 0 1996 by the Genetics Society of America
Noncomplementing Diploids From
Bacillus subtilis Protoplast Fusion:
Relationship Between Maintenance of Chromosomal
Inactivation
and
Segregation Capacity
Valerie Grandjean, Yolande Hauck, Jacques Le Derout and Luisa Hirschbein
Znstitut de Ginitique et Microbiologie, Universiti Paras-Sud, 91405 Orsay Cedex, France Manuscript received December 7, 1995
Accepted for publication August 1 , 1996
ABSTRACT
Fusions of Bacillus subtilh protoplasts from two genetically marked strains produce noncomplementing heterodiploid bacteria. These noncomplementing diploids (Ncds) carry both parental chromosomes, but only one is expressed. Fusion products of strains polymorphic for NotI restriction sites provide new physical evidence to support the conclusion that Ncds are not an artifact of cross feeding or cell adhesion. We show that reversible chromosomal inactivation can only account for the biparental trait of unstable Ncds. Two types of cells were recovered from the late progeny of unstable Ncds: Ncds with irreversible chromosome silencing (stable Ncds) and secondary recombinants that displayed a genomic mosaic NotI profile. Segregants from an unstable Ncd population gave rise to two viable haploid cell types. By contrast, stable Ncds segregated into a population of viable and inviable haploid cells. We propose that the latter are derived from irreversible chromosome silencing. Our results indicate that clonal popula- tions of stable Ncds are heterogenous and suggest that segregation and inactivation are independent parameters.
R
EVERSIBLE silencing of all or part of a chromo- some has been observed in many organisms (GOYON and FAUGERON 1989; ASSWet al. 1993; LYON 1993). Examples of the importance of the phenomenon during development can be found in mammals (Xchro- mosome inactivation in female development; GRANT and CHAPMAN 1988), in insects (inactivation of the pa- ternal set of chromosomes in somatic tissue; CROUSI et al. 1971) and in plants (silencing of foreign genes dur- ing the regeneration of transgenic plants; JORGENSEN1991). Unexpectedly, heritable inactivation of a redun- dant chromosome also occurs in prokaryotes (reviewed in THALER et al. 1990). The first evidence came from the work of HOTCHKISS and GABOR (1980). These authors observed that polyethyleneglycol (PEG)-induced pro- toplast fusion of strains of Bacillus subtilis carrying multi- ple markers produced diploid cells in which one of the two parental chromosomes was randomly silenced. These bacteria, called noncomplementing diploids
(Ncds), show the phenotype of only one parent (HOTCHKISS and GABOR 1980). The diploid nature of fusion product was demonstrated by several experi- ments. Extensive genetic experiments showed that the silent chromosome remained unexpressed until segre- gation occurred, after which it was reactivated (HOTCH-
KISS and GABOR 1980; SANCHEZ-RIVAS et al. 1982). A
diploid fusion product displaying phenotype A, grown in medium selective for parent A, typically yielded cells
Corresponding author: Luisa Hirschbein, Institut de C2nttique et
Microbiologie, Centre d’Orsay, Biitiment 409, 91405 Orsay Cedex, France. E-mail: [email protected]
Genetics 144: 871-881 (November, 1996)
of phenotype B at a frequency of 10-2-10-4. Therefore, Ncds characterized by following the inheritance of the biparental trait (BP) are unstable (SCHAEFFER and HIRSCHBEIN 1985). Purified DNA from Ncd clones ex- hibited all the transforming activities expected from a diploid donor (BOHIN et al. 1982). Most importantly, cells recombinant for parental traits A and B were iden- tified in the Ncd progeny, indicating that inactive nucleoids could coexist with active nucleoids in the same cell (GABOR and HOTCHKISS 1983; LEVI-MEYRUEIS et al. 1984). Segregating and recombinant Ncds con- verted spontaneously to a stable form (SANCHEZ-RIVAS et al. 1982; GUILLEN et al. 1985). The rate and frequency of this conversion depended on the particular clone being analyzed. The stable class of progeny displayed markers from the previously silent chromosome at a frequency of <lo”. Markers from the stabilized silent chromosome have been detected by three means: by inducing self-fusions with PEG, by assessing the trans- forming activity of purified DNA, and by analyzing the reactivation of a silent chloramphenicol acetyltransfer- ase (cat) gene (LOPEZ et al. 1986; LE DEROUT et al. 1992).
872 V. Grandjean et al.
1994). On the basis of reconstruction experiments, these authors concluded that fusion products manifest- ing Ncd properties corresponded to a mixture of paren- tal strains, and that recombinant cells resulted from the lysis of the haploid cell mixture and subsequent cell transformation. This controversy can only be resolved by using new techniques. In this paper we have com- bined pulsed-field gel electrophoresis (PFGE), PCR,
and segregation analysis of strains bearing polymorphic NotI restriction sites (ITAYA and TANAKA 1991) to obtain further evidence of the genetic and physical structure of Ncds and recombinant cells. We demonstrate that the persistence of the biparental trait of a fusion prod- uct is not observed with a mixture of parental haploids. Unstable Ncds do result from cell fusion. A primary Ncd colony harbors an heterogeneous cell population of pure segregated haploid parental cells and Ncds. In the progeny of unstable Ncds we detected recombinant cells that displayed a phenotype and a mosaic genomic NotI profile expected from the interaction between the two parental chromosomes. The segregation of both parental genomes persists in unstable Ncds, and most likely, in the stable Ncds. Failure to find the expected cell types suggests that segregation and inactivation function as independent parameters.
MATERIALS AND METHODS
Bacterial strains: Two B. subtilis strains were used in this study. The first, SI5 (purB?4, u r a l , trpC7, rfm486) (LEVI et al. 1977), requires adenine, uracil, and tryptophan, and is resis- tant to rifampicin. The second, BEST4087 (metC7, trpC2, pro, leu, c a t ) , requires leucine, tryptophan, proline, and methic- nine, and is resistant to chloramphenicol (ITAYA and TANAKA 1991). The primary advantages of these strains are their differ- ent morphologies on complete medium and their specific NotI restriction fragment length polymorphisms (RFLP)
.
Media: Media and buffer employed in fusion experiments were described in SCHAEFFER et al. (1976). Parental and Ncd strains were grown in liquid minimal medium (ANAGNOSTO- POULOS and SPIZIZEN 1961) supplemented with the necessary growth factors.
Protoplast fusion and analysis of regenerants Bacterial protoplast fusions were performed according to the method of SCHAEFFER et al. (1976). Analysis of regenerants, which allows identification of unstable and stable Ncds, was per- formed as described previously (LE DEKOUT et al. 1992). Typi- cally, well-se arated regenerated colonies, obtained by plating
or 10- -fold dilutions of fused protoplasts on the nonse- lective regeneration medium, were picked at random with a toothpick and placed on media selective for the two parents (A and B media) and also on minimal medium. Colonies that grew on both parental media (A and B media) but not on minimal medium were termed “biparental” (BP) and were subjected to several more cycles of toothpick selection. An unstable Ncd of A(B) phenotype will display the following segregation pattern: colonies obtained from growth medium A will continue to manifest the BP trait. In contrast, when growth on medium B is further tested, the B colonies grow only on medium B. The unstable Ncd of phenotype B(A) will show the reciprocal segregation pattern: cells grown on medium B will exhibit the BP trait, while those grown medium A will not.
B
Evaluation of diploid cells present in an Ncd clone: A frac- tion of the cell population from an unstable Ncd clone of A(B) phenotype was resuspended, diluted, and spread on solid selective media A and B. On medium A, both the Ncd and the segregated haploid cells containing the A chromo- some will grow. On medium B, only the haploid cells con- taining the segregated chromosome B can grow. We assume that all Ncd bacteria that segregate give rise to two haploid cells, one A and one B cell, so that the titer of haploid cells (with chromosome B) growing on medium B will equal to the titer of haploid cells with chromosome A. Therefore, the titer of Ncd cells is equal to the titer of cells growing on medium A minus the titer of the cells growing in medium B. The proportion ( d ) of Ncd cells in the clonal population is given by
d = ( N A - N B ) / ( N A
+
N B ) , (1)where NA is the titer of bacteria growing on medium A and NB the titer of bacteria growing on medium B. The same method was used to quantify the Ncd cells having a B(A) phenotype.
Segregation rate per generation: The segregation rate per generation corresponds to the proportion of Ncd cells that segregate and produce two haploid cells per generation. If
No is the number of Ncd cells found in the original clonal population (NA - N B ) , and if y is the segregation rate per generation ( n ) , then the number of Ncd cells present in the next generation is given by N, = 2(N0 - Noy). After n generations, the number of Ncd cells, N,,, is given by 2”N,(1 - y ) ” . The proportion of Ncd cells ( d ) is N,, divided by the total number of haploid plus Ncd cells (2“ N o ) ; d = (1 - y)”. The segregation rate is given by
= 1 - e l n ( R / n
.
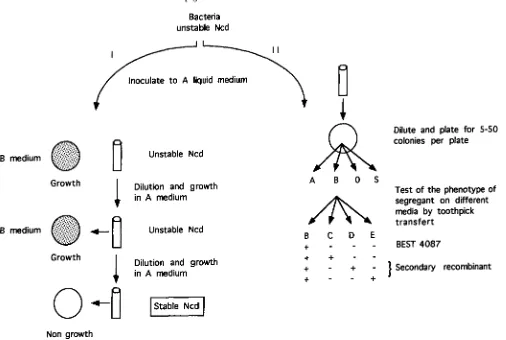
(2)Selection of secondary recombinants and stable Ncds: Sel- ection of stable Ncd and secondary recombinants is described in Figure 1.
Selection of stable Ncds: Stable Ncds were obtained as segre- gants of unstable Ncds according to procedures described by GUILLEN et al. (1985) with the following modification. A col- ony that exhibited a BP phenotype was inoculated into liquid A medium, which allows the parent S15 strain to grow but not the parent BEST4087 strain. After 20 generations, 100 ml of the culture were plated onto solid B medium. If the BP trait was retained, the culture was diluted and grown again. A clonal population was considered to be composed of stable Ncd when it did not manifest the BP trait but contained mark- ers from the stabilized silent chromosome (revealed as pre- viously described; THAI.EK 1990).
Selection of secondaly recombinants: Recombinants arising in the progeny of unstable Ncd were named secondary recombi- nants to distinguish them from directly selected primary re- combinants appearing after fusion (LEVI-MEYRUEIS and SAN-
CHEZ-RIVAS 1984). An unstable Ncd colony of strain S15 (BEST4087) that retained the BP trait through several pas- sages was subcloned by culturing for about seven generations in liquid medium selective for the S15 parent. The culture was then diluted and plated for single cells on minimal medium supplemented with S15 requirements. Any colonies that could also grow on BEST4087 medium were further tested for their growth requirements. Some clones still exhibited the BEST4087 phenotype and others exhibited a recombinant phenotype. The recombinants thus obtained were subcloned in liquid selective medium and then analyzed again.
Epigenetic Inactivation in B. subtilis
Bacteria unstable Ncd
873
Inoculate to A liquid
Growth
1
Dilution and growth in A mediumGrowth
1
Dilution and growth in A medium0
-B
I
Stable NcdI
A B O S
A
B C D E
+
-+
+
"
+
-
+
-+ " +
"
Dilute and plate for 5-50
colonies per plate
Test of the phenotype of segregant on different media by toothpick transfert
BEST 4087
}
Secondary recombinantNon growth
FIGURE 1.-Analysis of noncomplementing diploid of type A(B). (I) Selection of stable Ncd. (11) Selection of secondary recombinant. A, B, C, D, E, S and 0 correspond to the solid AUO, MLBP, MOL, MOP, LOP, complete and minimum medium, respectively. Bacteria are an inoculum of lo4 cells from a patch of an unstable Ncd of S15(BEST4087) phenotype. This patch is derived from the initial cell fusion uroduct after 40 generations of growth. A, adenine; U, uracil; v B , tryptophan; M, methionine; L, leucine; P, proline;
+,
growth; -, non-growth.lowing digestion with restriction enzymes (Boehringer Mann- heim).
Pulsed-field gel electrophoresis: High-molecular weight chromosomal DNA was prepared in agarose plugs (ITAYA and TANAKA 1991). Slices of these were incubated for 4 hr at 37" with NotI using the buffer supplied by the manufacturer. Electrophoresis was performed using a contour-clamped ho- mogeneous electric field apparatus (CHEF DRII, Biorad). Agarose gels (1%) were pre ared and electrophoresed in 0.5X TBE buffer at 12 Vcm- . Pulse time increased linearly from 5 to 20 sec over 20 hr. Concatemeric lambda DNA was used as size markers.
Southern blot analysis: DNA was blotted onto Hybond-N nylon membranes (Amersham) by capillary transfer (hlA-
NIATIS et al. 1982). DNA was cross-linked to the membrane at 310 nm for 5 min. Nonradioactive detection using digoxy- genin-dUTP as the labeling component was performed (Boe- hringer Mannheim). Hybridizations were carried out at 68". Stringent washes were performed twice in 0.1% SSC and 0.1 %
SDS at 60" for 15 min. Hybridized probe DNAs were detected using a chemiluminescent substrate (AMPPD).
For the identification of the recombinant 200-kb Not1 band, Not1 digests of S15 and BEST4087 chromosomal DNAs were probed with the 200-kb NotI recombinant fragment. The cat probe was prepared from the 1.8-kb BamHI-EcoRI fragment of plasmid pHV32 carrying the cat sequence (NIAUDET and EHRLICH 1979).
Localization of cut sequence by PCR assay: The chromo-
P
somal DNAs of the S15 and BEST4087 strains were digested with NotI and then subject to PFGE in low-melting-point agarose gels. An agarose plug was picked around the 60-kb NotI fragment with a Pasteur pipette and used directly for PCR amplification (SAIKI 1990). Two oligomers designed to amplify a 827-bp fragment of the cat gene in the chromo- somal DNA of B. subtilis were synthesized with an Applied Biosystems automated oligonucleotide synthesizer. The se- quence of the 27-nucleotide (nt) 5'-3' primer was CCC- ATTAGTTCAACAAACGAAAATTGG, and of the 24nt 3'-
5' primer was CCGACTGTAAAAAGTACAGTCGGC, corre-
sponding to nucleotides 1095-1122 and 1922-1946 of plas- mid pC194, respectively (HORINOUCHI and WEISBLUM 1982). Taq DNA polymerase was obtained from Appligene. Gene amplification was performed with a Hybaid Thermal Reac- tor (Cera Labo) using an initial denaturation step of 95" for 5 min followed by 30 cycles of amplification, each at 95" for 1 min/60" for 1 min/72" for 1 min, and a final incuba- tion at 72" for 10 min to extend all initiated molecules.
RESULTS
874 V. Grandjean et al.
TABLE 1
Fusions between S15 and BEST4087 strains
Efficiency of
regeneration Colony phenotype
Cross S15 X S15 BEST4087 Fusion products S15 BEST4087 BP
BEST4087 (%)
(%I
tested (%) (%) (%)1 22 25 . 300 43 53 4
2 20 15 300 53 45 2
Equal numbers of protoplasts of the two strains were fused medium so that five to 50 colonies arose on each plate.
of Ncds. Fusion crosses between S15 and BEST4087 were performed and Ncd clones of defined phenotypes were identified as described in MATERIALS AND METH- ODS. The characteristics of the two fusion experiments carried out for this study are described in Table 1. The properties of the fusion products are detailed below.
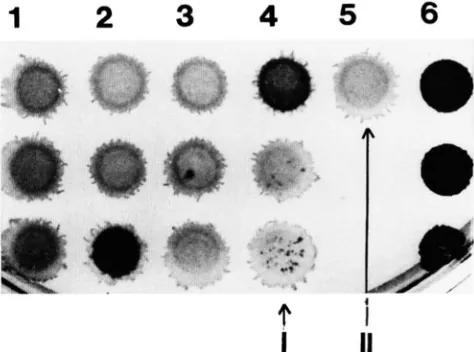
Sector-forming colonies are present only in Ncd p o p ulations: When plated on complete medium, the mor- phology of BEST4087 and S15 colonies was different. BEST4087 colonies were transparent and irregular, whereas the S15 colonies were opaque and round. When plated on selective medium, there were no ob- servable morphological differences between the two strains. A single Ncd colony from the regeneration plates, characterized by a BP trait and a BEST4087(S15) phenotype, was resuspended, diluted and plated on minimal medium supplemented with the BEST4087 re- quirements to obtain 10-20 colonies per plate. About
300 random single colonies were analyzed on the two parental selective media and on complete medium. All these colonies had BEST4087 morphology, although 20% of them displayed opaque sectors (Figure 2).
These sectoring colonies contained bacteria that grew on both parental selective media but not on minimal medium.
In a reconstruction experiment, S15 and BEST4087 bacteria were cocultivated for five generations on me- dium that allowed the growth of both strains. The mix- ture was then spread on minimal medium suitable for growth of BEST4087 cells. Three hundred single colo- nies were analyzed by plating on each parental medium
as well as on complete medium. All colonies had a pure haploid BEST4087 phenotype and failed to show any sectoring. Thus, only cells originating from fusion crosses were capable of giving sectors and forming colo- nies on either parental media. No bacteria could grow on minimal medium. This implies that sectors that arise during growth of an Ncd of BEST4087(S15) phenotype are due to the segregation and the reexpression of the previously silent chromosome.
The number of counter-selected bacteria increases dur-
ing growth of Ncd clones in selective medium: The fol- lowing experiments were devised to test the possibility that the BP trait of Ncds was actually due to crowfeeding
with polyethylene glycol, diluted, and plated on regeneration
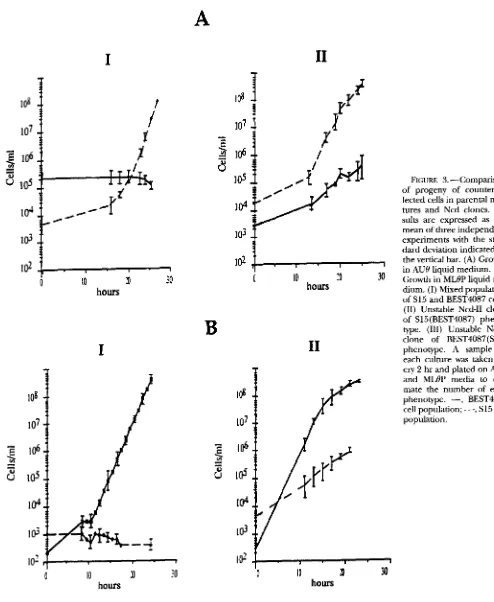
of parental strains. In a reconstruction experiment, equal numbers of parental S15 and BEST4087 cells were mixed and grown on each of the two selective media. As shown in Figure 3, A-I and B-I, the number of counter-selected bacteria remained stable or decreased in both selective media after 25 hr of growth (20 generations), the pre- dicted result in absence of crowfeeding. Figure 3A-I1
shows the evolution of the BEST4087 and S15 cell popula- tions for a characterized unstable Ncd (Ncd-11) clone of phenotype S15(BEST4087). This clone was obtained from a purified regenerated colony, and was grown for 15 cell divisions in liquid medium selective for the S15 strain. The number of S15 cells increased as expected, but an increase in the number of BEST4087 cells was also o b served. The same pattern was observed starting with an independent unstable Ncd clone of reciprocal phenotype
F~CURE 2.-Morphology of a biparental (BP) colony on
108
107
I
t
f
i
0IO
r) 30hours
Epigenetic Inactivation in B. subtilis
A
B
I
0 10
n
:0hours
I1
f
I1
875
FIGURE 3.-Comparison of progeny of counter-se- lected cells in parental mix- tures and Ncd clones. Re- sults are expressed as the mean of three independent experiments with the stan- dard deviation indicated by
IO2
lr----”--
in AUB liquid medium. (B) the vertical bar. (A) Growth0
I O
30J0
Growth in MLBP liquid me-hours
dium. (I) Mixed population of S15 and BEST4087 cells. (11) Unstable Ncd-I1 cloneof S15(BEST4087) p h e n e type. (111) Unstable Ncd-I clone of BEST4087(S15) phenotype. A sample of
each culture was taken ev- ery 2 hr and plated on AUB and MLBP media to esti- mate the number of each phenotype. -, BEST4087 cell population; -
-
-, S15 cell popdation.BEST4087(S15), clone Ncd-I (Figure 3EII). This experi- These experiments demonstrate that the peculiar BP trait ment was repeated six times with different unstable Ncd of Ncd clones can only be due to segregation and reex- clones that arose from unrelated fusion crosses. They all pression of the previously silent chromosome. They vali- gave the same segregation pattern: an increasing number date the diploid nature of Ncd cells and exclude the of cells bearing the phenotype of the counter-selected possibility of a cross-feeding phenomenon.
876 V . GranGjean P I 01.
TABLE 2
Estimation of the Ncd proportion and segregation rate in BP clones
Initial no. o f bacteria exhibiting the phenotype of
Expressed BP
Unexpressed
chromosome chromosome Ncd proportion in Segregation
clones N:, N B BP clone (%) rate
Ncd-I 5 x 10:' 7 x 10' 75 1 . 1 x 10-2
Ncd-I1 3 x 10:' 3 x 10' 82 9.9 x 1 0 - 3
Ncd-I11 3 x 10' 2.8 x 10' 98 7 x 10-4
Aliquots o f each o f the three original independent Ncd clones (Ncd-I, Ncd-11, Ncd-111) characterized by the
BP trait were resuspended, diluted and spread on solid selective media to score the cell number of each phenotype. The cell number corresponding to the expressed and unexpressed chromosome phenotype are noted X,, and NII, respectively. The percentage of Ncd bacteria present in the population (d) is given by Equation 1 and the segregation rate (y) is given by Equation 2 (see MATERIALS AND METHODS). As the formation of one colony from a single cell requires 23 cell divisions, n is equal to 2.5.
segregate the previously unexpressed chromosome im- plies that the cell population is heterogeneous, com- posed of haploid and diploid cells. The distribution
of each type of cell can be evaluated by analyzing the phenotype of cells derived from single clones. If segre- gation of an Ncd cell of phenotype A(B) produces two
viable haploid cells of genotypes A and €5, the propor- tion of Ncd cells (d) can be calculated from Equation
1 (see MATERIALS AND METHODS). Table 2 illustrates the percentage of Ncds in three original independent Ncd clones and shows the calculated segregation rate per generation. For these particular clones, the proportion of Ncd cells within the clonal population after 25 gener- ations should be 75%, 82% and 98%, respectively. These relatively high estimates are based on the assump tion that the segregation of an Ncd produces two viable haploid cells of eadh parental phenotype. To test this assumption, we have analyzed the Not1 RLFP pattern of unstable Ncds.
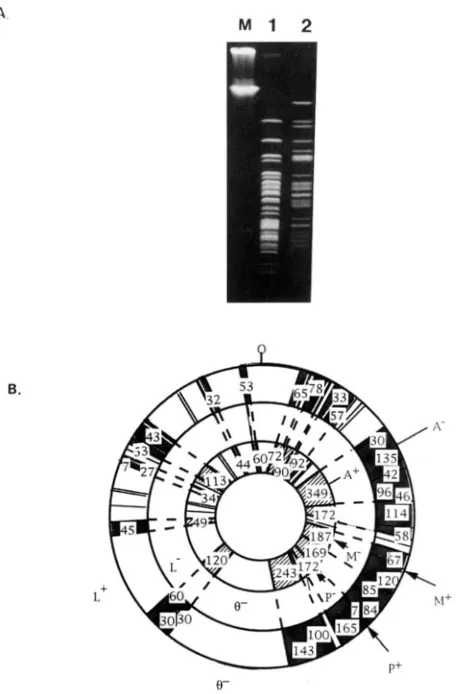
The S15 and BEST4087 chromosomes have distinct
Not1 restriction patterns, sharing only 46% common fragments (Figure 4). The progeny of unstable Ncds should display the RFLP pattern of both parental strains if 10% or more of cells in that culture are Ncds. This DNA detection limit in PFGE was deduced from recon- struction experiments using different proportions of each parental haploid strain (data not shown). The un- stable original Ncd-I, Ncd-I1 and Ncd-111 clones were grown for 40 generations. Their chromosomal DNA was then extracted, digested with Not1 and analyzed by PFGE. The patterns were identical to either the S15 parental strain for Ncd-I1 and Ncd-111, o r to the BEST4087 parental strain for Ncd-I (not shown). This result indicates that, despite genetic evidence for the presence of both parental phenotypes, unstable Ncds were present in quantities below the 10% threshold detectable by PFGE. M'e conclude that the proportion
of Ncd cells within a single colonv is greatly overesti- mated when it is determined uniquely from segregation rates.
A
B.
M 1 2
e-
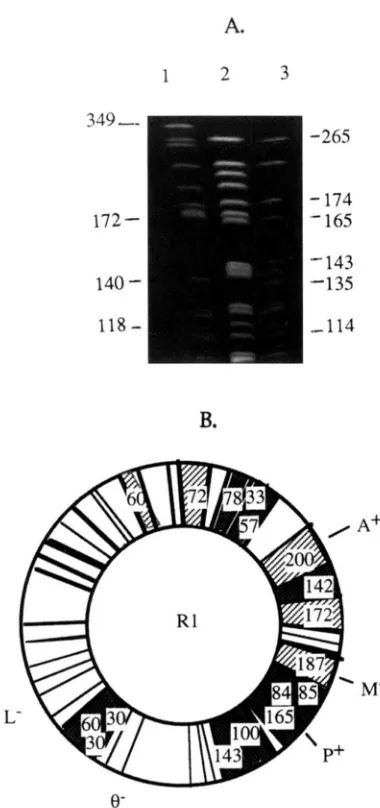
FIGURE 4.-PFGE analysis of chromosomal DNA of SI5 and BEST4087 strains. (A) Ethidium bromide staining. Lane M, concatemeric lambda DNA size markers; lane 1 , Not1 digest of S15; lane 2, Nor1 digest of BEST4087. (B) Comparison of
Epigenetic Inactivation in R. s~tbtilis 877
143kb NotI fragment, are specific to the S15 strain. Other fragments, such as the 187-kb NotI fragment, are characteristic of the BEST4087 strain. In addition, two
fragments of -200 and 142 kb are not found in either parental Not1 profiles. " h e n t h e 200-kb fragment was used as a probe, it hybridized to bands from both paren- tal strains. The new NotI fragment (200 kb) originated from a recombination event between hvo homologous regions located on the 349-kb Not1 fragment of parental strain BEST4087 and the 135-kb Not1 fragment of strain S15 (Figure 5). At least six crossovers are required to account for the observed NotI profile of clone R1. The sum of all the restriction fragments was -4000 kb, which corresponds approximately to the size of the chromosome of a
R.
s ~ b t i l i s haploid cell.A.
1 2 3
B.
A+
"
e
FIGURE 5.-No/l pattern of the R I recombinant. (A) PFGE analysis of chromosomal DNA of the MLB recombinant clone. Lane I , Not1 digest of the REST4087 chromosomal DNA, lane
2, No/I digest of' the RI chromosomal DNA; lane 3, No/I digest of the SI5 chromosomal DNA. (R) lVo/I restriction map of
the RI clone.
Secondary recombinants arise from croswver events that involve fragments of DNA larger than I00 k b Sec- ondary recombinant clones (defined in MATERIALS AND METHODS) were recovered from the progeny of the un- stable Ncd-I, Ncd-I1 and Ncd-111 clones. This is evidence that Ncds persist in the progeny of a fusion product. Primary BP colonies were repeatedly plated, and recom- binant clones were identified in the unstable Ncd prog- enies after 40 generations (Figure 1). The recombi- nants were analyzed after purification by restreaking and further growth in liquid selective medium (see MA- TERIALS AND METHODS).
The Not1 restriction pattern of one of these recombi- nants, clone R1, is illustrated in Figure 5. The genomic Not1 profile of clone R1 appears as a mosaic of S15 and BEST4087 parental Not1 fragment., and correlates with the observed phenotype. Some fragments, such as the
Stable Ncd clonal populations are composed mainly of haploid cells: One of the most serious limitations in examining the clonal composition of stabilized Ncds
(GUII.L.EN d nl. 1985; LOPEL Pt nl. 1986) is that, by defi-
nition, stabilized Ncds cannot be distinguished from haploid cells by phenotype alone. This is not true for the unstable class. Their peculiar segregation pattern clearly supports the existence of transient diploid cells. To circumvent this problem, we used PFGE to analyze the DNA of putative stable Ncd clones that were derived from the progeny of the unstable Ncds clones, Ncd-I, Ncd-I1 and Ncd-111, after 100 generations of growth. Chromosomal DNA of 10 putative stable Ncd clones with a S15(BEST4087) phenotype was extracted, di- gested by NotI, and analyzed by PFGE. These clones
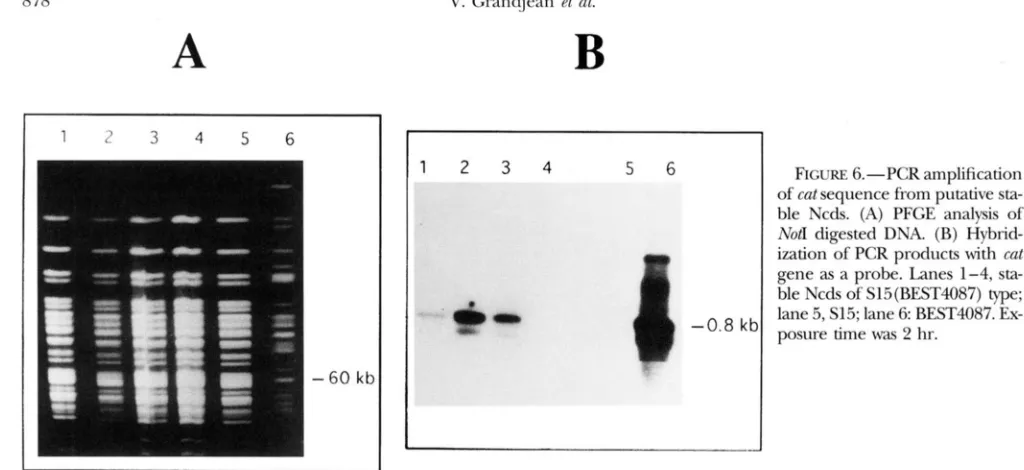
all exhibited the NotI pattern that corresponds to the expressed chromosome (data not shown). M'e searched for the unexpressed chromosome to determine whether the Not1 DNA patterns corresponded to a pure haploid population or to a heterogeneous progeny that could include a minority of stabilized Ncds. Two a p proaches were taken. The first was the identification of a DNA sequence from one parental strain that is not expressed in the putative stable Ncd clone, the cat gene sequence of parent BEST4087. This sequence is present only in the BEST4087 chromosome and is located within the 60-kb Not1 fragment. Figure 6 shows the re- sults of one of these experiments. The DNA from four putative stable Ncd with a S15(BEST4087) chloram- phenicol-sensitive phenotype was subjected to PFGE after Notldigestion (Figure 6A). A gel plug containing the 60-kb fragment was used for PCR amplification. N o DNA amplification was observed when the 60-kb fragment from the PFGE gel corresponding to S15 was used as a template. By contrast, amplification of the
cat sequence was obtained when the 60-kb fragment- containing gel plugs from three out of four putative stable Ncds and the BEST4087 parental strain were used
8 f 8
A
V. GrancIjean PI nl.
1 2 3 4 5 6
L
-
60 kbJ
the intensity of the hvbridization cat signal is weaker than that obtained from the hybridization with the 60-
kb Not1 fragment from the BEST4087 parental strain (Figure 6B).
The second approach was to use PEGinduced self fusion of the same stable Ncds, which should lead to the destabilization of both chromosomes and reactiva- tion of the previously unexpressed markers (GLVLLEN
P t nl. 1985; LOPEZ pt 01. 1986). Table 3 illustrates the
results of this experiment. After self-fusion of the puta- tive Ncd protoplasts and regeneration on nonselective plates, recombinants and BP colonies were recovered, revealing the diploid nature of the Ncds. However, the percentage of destabilized cells was never >3%. The great majority of the cells within the regenerated colo- nies were haploids, displaying only one of the parental phenotypes.
Together, the amplification and self-fusion results in- dicate that, although heterodiploids are persistent within the clonal population, only a minority of Ncd cells are present within a so-called stable Ncd colony.
DISCUSSION
Instability tests have been used to define Ncds
(HOTCHKISS and GABOR 1980). Colonies were consid-
RCURE 6.-PCR amplification of cut sequence from putative sta- ble Ncds. (A) PFGE analysis of Not1 digested DNA. (B) Hybrid- ization of PCR products with cut
gene as a probe. Lanes 1-4, sta- ble Ncds of Sl5(BEST4087)
w ;
lane 5, S15; lane 6: BEST4087. Ex- posure time was 2 hr.
ered to be Ncds if, during growth, fusion products of one parental phenotype segregated bacteria that ex- pressed all the genetic markers of the other parent. Also, it was necessary that neither parent grew on mini- mum medium and that each had different nutritional requirements Only those colonies showing persistence of this biparental behavior after subcloning in liquid medium were considered to be unstable Ncds. “ S u b cloning in liquid medium” meant that patches from putative Ncd colonies were inoculated into rich liquid medium and grown overnight. Suitable dilutions were then spread onto plates, so that <lo0 colonies arose on a single plate. The strong inference was that each colony that arose by this procedure was the product of a single cell. The capacity of late Ncds to segregate recombinants with markers from both original parents provided the first proof that a subset of regenerated products contained two complete, genetically distinct, genomes, one of which had been phenotypically unex- pressed (HOTCHKISS and GABOR 1980; Ltw et al. 1980). In the present work, we demonstrate by precise recon- struction experiments in liquid selective medium that the BP trait of unstable Ncds is not an artifact arising from cross-feeding or cell adhesion. This result is in
TABLE 3
Reactivation of nonexpressed chromosome after Ncd self-fusion
Phenotypes obtained among regenerated cells
No. of colonies Recombinants Biparentals Parental
Strain analyzed (%) (9%) (%)
Ncd 120
-
-
100Ncd
+
P E G 416 0.5 97.11 2.4Single colonies from an Ncd clone of phenotype S15, classified as a stable Ncd (segregation rate < lo”)
were picked, diluted, and plated onto SI5 selection medium. Then 25-30 colonies were picked and grown in liquid medium before performing the PEG self-fusion experiments.
Epigenetic Inactivation in B. subtilk 879
apparent contradiction with that of HAUSER and KARA
MATA (1992, 1994). However, these authors used solid media for their reconstruction experiments, which would not exclude cross-feeding. The BP behavior they observed probably resulted from artifactual cell adhe- sion, as they have postulated.
We provide additional proof that the BP trait o b
served during growth in selective liquid medium results from segregation and reexpression of the previously silent parental chromosome. The presence in the late progeny of single Ncd clones of recombinant bacteria with complex crossover patterns provides further evi- dence for the persistence of Ncds during cell division. We have shown that the genetic composition of late recombinant bacteria is determined by interactions be- tween the unexpressed and expressed chromosomes in the same cell. The genomic structure of each recombi- nant cell contained RFLPs characteristic of both paren- tal chromosomes. The mosaic NotI pattern that we ob-
served could only arise from multiple crossovers involving DNA exchanges of an average size of 100-
350 kb (Figure 5). The hypothesis that DNAse-resistant transformation accounts for this mosaic NotI pattern seems highly unlikely. Normally, transformation in B. subtilis requires recipient cells to be in a competent state, and is RecA (formerly RecE)-dependent (DUBNAU
and CIRIGLIANO 1972; DUBNAU et al. 1973). The growth medium used in the current work is not conductive to cell competence, and at no time during growth were the cells in a competent state. Furthermore, Ltw- MEYRUEIS et al. (1980) and FTOUHI and GUILLEN (1990)
showed that fusion of parental strains with a RecA geno- type did not prevent the generation of recombinant cells in the segregating progeny. Thus, the recA product seems not to be required for recombination occurring between chromosomes in the diploid bacteria. How- ever, we cannot rule out entirely that bacteriophage- encoded recombination functions could be responsible for this RecA independence. The B. subtilis strains 168
and the majority of its derivatives carry resident pro- phages such as SPP, as well as the defective phage, PBSX
(ZAHLER 1993).
We have considered the possibility that illegitimate recombination, which is RecA independent (FRANKLIN 1967), may account for the recombinant cells obtained among the late progeny. Illegitimate recombination oc- curs at a rate of lo-’ (HOFEMEISTER et al. 1983; GRANDJEAN et al. 1993). B. subtilis recombinants from fusion experiments arose at the much higher rate of 5
X to 2 X lo-’ (GABOR and HOTCHKISS 1983;
FTOUHI and GUILLEN 1990).
The fact that perfect sets of reciprocal recombinants have been recovered from individual postfusion ~ 0 1 0 -
nies further supports our argument that recombination does not result from transformation events. In particu- lar, all possible combinations of eight parental markers, and their respective reciprocals, were found in the prog-
eny of individual unstable Ncds (GABOR and HOTCHKISS 1983; reviewed by HOTCHKISS and GABOR 1985). This finding proved that the expressed and silent chromo- somes were able to undergo multiple rounds of recom- bination during celldivision.
According to previous studies (HOTCHKISS and GA- BOR 1980), an individual Ncd colony that had displayed a BP trait for 50-100 generations, at a segregation rate of lo-‘ to should exhibit a large proportion of diploid cells. The fraction of diploids should be suffi- ciently large that two different RFLP patterns will be seen in NotI digests, characteristic of the two parental chromosomes. In the current work, however, all pri- mary Ncd colonies analyzed displayed only one Not1
RFLP pattern. The pattern detected was identical to that of the parental chromosome that was expressed in the population. Thus, the number of Ncd cells per col- ony, calculated from the segregation rate, differed sig- nificantly from the estimated number of Ncd cells per colony determined by PFGE analysis. This discrepancy can be explained by an unequal viability of the segreg- ants.
Stable Ncds were defined by their stabilized pheno- type, and it was assumed that a clonal population was homogeneous (GUILLEN et al. 1985). However, we have shown, using PFGE analysis, PCR amplification and PEGinduced self-fusion destabilization, that the cell populations from each of the stable Ncd colonies were heterogeneous. We found from the PFGE analysis that, in unstable Ncd clones, most of the cells were haploids and < l o % were diploids. This result suggests that, dur- ing growth, cell segregation occurs both in unstable and in stable Ncds. Thus, in contrast to predictions based on classical genetic experiments, the clonal popu- lation from a single stable Ncd colony will always be heterogeneous.
Our results lead to a new understanding of the com- position of the clonal populations that arise from single stable and unstable Ncds. We have confirmed that Ncds persist even when segregation and bacterial replication takes place for a considerable number of generations. We also infer from our results that only a minority of the cells within a primary Ncd colony actually have the Ncd genotype. After continuous subcloning and re- peated growth in selective medium, the population de- riving from a primary colony is composed in majority of haploid cells of the expressed parental phenotype. Haploid recombinants and reversible or irreversibly si- lenced Ncds account for only a minority of the colony- forming units.
880 V. Grandjean et al.
our data, we propose that, upon segregation, an irre- versibly silenced genome will not be able to support cell growth. Thus, an Ncd cell with an inactive chromo- some that has reached the irreversible stage will segre- gate to produce viable haploid cells bearing only one parental phenotype and inviable cells. Two sets of re- sults support this assumption. First, only a few cells re- covered from single clones were reciprocal recombi- nants. As judged from colony counts, the two allelic types were present in unequal proportions with a rela- tive frequency of 1 per 100 (GABOR and HOTCHKISS 1983). Second, self-fusion experiments suggested that diploids represent only a small fraction within the stable Ncd clonal population (GUILLEN et al. 1985; LOPEZ et al. 1986; this work). Continuous segregation and the different viability of the segregants may explain why PEG self-fusion of progeny derived from Ncds by re- peated subcloning and growth in selective medium failed to show evidence for diploid cells (LOPEZ 1985).
The fact that Ncds colonies are heterogeneous may account for previous results that challenged the exis- tence of Ncds (HAUSER and KARAMATA 1994). Indeed, inactive alleles were difficult to detect by hybridization (reviewed in THALER et al. 1990). Using PCR, we have been able to detect the presence of silenced alleles (this work; and J. LE DEROUT and L. HIRSCHBEIN, in prepara- tion). Thus, our conclusions concerning the fate of the unexpressed DNA within an Ncd colony have important consequences for the design and interpretation of ex- periments in this field. In vivo evidence for the propaga- tion of gene silencing over large coding regions through nucleoprotein structures has recently been ob-
tained in E. coli (LYNCH and WANC 1995). Nucleopro- tein structures are also implicated in heritable gene silencing in fusion products of Gram-positive bacteria (BOHIN et al. 1982; GUILLEN et al. 1985). Our results also imply that it is necessary to select a homogenous cell population on which to perform accurate biochemi- cal and structural studies of chromosomal inactivation in Ncds.
We thank B. HALL, J. A r . 0 ~ ~ 0 , D. THAI.ER and M. BLOT for discus- sions and comments on this manuscript, and M. ITAYA for his gift of the BEST4087 strain. This work was supported by Centre Nalional d e la Recherche Scientifique (URA 1354), Institut National de la Santi et de la Recherche Midicale grant no. 900106 and Association Pour la Recherche Sur le Cancer grant no. 6794 to L.H. V.G. was supported by the Ministere de la Recherche et de la Technologie.
LITERATURE CITED
ANAGNOSTOPOULOS, C., and J. SPIZIZEN, 1961 Requirements for transformation in Bacillus subtilis. J. Bacteriol. 81: 741-746.
ASSAAD, F. F., K L. TUCKER and E. R. SIGNER, 1993 Epigenetic re- peat-induced gene silencing (RIGS) in Arabidopsis. Plant Mol. Biol. 22: 1067-1085.
BOHIN, J. P., K. BEN KHALIFA, N. GUILLEN, P. SCHAEFFER and L. HIRSCHBEIN, 1982 Phenotypic expression i n vivo and trans- forming activity in vitro: two related functions of folded bacterial chromosome. Mol. Gen. Genet. 185: 65-68.
CROUSI, H. V., A. BROWN and B. C. MUNFORD, 1971 L-chomosome
inheritance and the problem of “imprinting” in Sciara (Sciaridae. Diptena). Chromosome 3 4 324-339.
DUBNAU, D. A., and C. CIRIGLLANO, 1972 Fate of transforming DNA following uptake by competent Bacillus subtilis. 111. Formation and properties of products isolated from transformed cells which are derived entirely from DNA. J. Mol. Biol. 6 4 9-29.
DUBNAU, D. A., R. DAVIDOFF-ABELSON, B. SCHER and C. CIRIGI.IANO,
1973 Fate of transforming deoxyribonucleic acid after uptake by competent Bacillw subtilis: phenotypic characterization of ra- diation sensitive recombination deficient mutants. J. Bacteriol.
FTOUHI, N., and N. GUILLEN, 1990 Genetic analysis of fusion recom- binants in Bacillus subtilis: function of the recE gene. Genetics FRANKLIN, N., 1967 Extraordinary recombinational events in Esche- richia coli. Their independence of the rec’ function. Genetics GABOR, M. H., and R. D. HOTCHKISS, 1983 Reciprocal and nonrecip- rocal recombination in diploid clones from Bacillus subtilis proto- plast fusion: association with the replication origin and terminus. Proc. Natl. Acad. Sci. USA 80: 1426-1430.
GOYON, C., and G. FAUGERON, 1989 Targeted transformation of As-
cated DNA sequences. Mol. Cell. Biol. 9: 2818-2827.
cobolus immesus and de novo methylation of the resulting dupli- GRANDJEAN, V., J. NGLWEN, Y. HAUCK and L. HIRSCHBEIN, 1993 Estab lishment of a new replicon generated from an integrational plas- mid and a cryptic pUBllO origin-like region in Bacillus subtilis.
Plasmid 30: 1-13.
GRANT, S. G., and V. M. CHAPMAN, 1988 Mechanisms of X-chromc- some replication. Annu. Rev. Genet. 22: 199-233.
GUILLEN, N., M. H., GABOR, R. D. HOTCHKISS and L. HIRSCHBEIN,
1982 Isolation and characterization of the nucleoid of non- complementing diploids from protoplast fusion in Bacillus subti-
lis. Mol. Gen. Genet. 185: 69-74.
GUILLEN, N., M. AMAR and L. HIRSCHBEIN, 1985 Stabilized non- complementing diploids (NCD) from fused protoplast products of B. subtilis. EMBO J. 4: 1333-1338.
HAUSER, P. M., and D. K A R A M A T A , 1992 Ploidy of Bacillus subtilis
exfusants: the haploid nature of cells forming colonies with bipa- rental or prototrophic phenotypes. J. Gen. Microbiol. 138: 1077-
1088.
HAUSER, P. M., and D. KARAMATA, 1994 No experimental support for diploidy and chromosomal silencing. Microbiol. 140: 2-3.
HOFEMEISTER,B.D.,M.ISRAELI-RECHES~~~D.DUBNAU,~~~~ Integra-
tion of plasmid pE194 at multiple sites of the Bacillus subtilis
chromosome. Mol. Gen. Genet. 189: 58-62.
HORINOUCHI, S., and B. WEISBLUM, 1982 Nucleotide sequence and functional map of pC194, a plasmid that specifies inducible chloramphenicol resistance. J. Bacteriol. 150: 815-825.
HOTCHKISS, R. D., and M. H. GABOR, 1980 Biparental products of bacterial protoplast fusion showing unequal parental chromc- some expression. Proc. Natl. Acad. Sci. USA 77: 3553-3557.
HOTCHKISS, R. D., and M. H. GABOR, 1985 Protoplast fusion in
Bacillus and its consequence, pp. 109-149 in The Molecular Biology ofthe Bacilli, edited by D. A. DUBNAU. Academic Press, New York. ITAYA, M., and T. TANAKA, 1991 Complete physical map of the Bacil- lus subtilis 168chromosome constructed by a genedirected muta- genesis method. J. Mol. Biol. 220: 631-648.
JORGENSEN, R., 1991 Silencing of plant genes by homologous trans- genes. AgBiotech News Inform. 4: 265N-273N.
LE D ~ R O U T , J., D. S. THAI.ER, N. GUILL.EN and L. HIRSCHBEIN, 1992
The spoOA gene is implicated in the maintenance of non-comple- menting diploids in Bacillus subtilis. Mol. Microbiol. 6: 1495- 1505.
L ~ v I , c . , C. SANCHEZ-RIVAS and P. SCHAEFFER, 1977 Further genetic studies on the fusion of bacterial protoplasts. FEMS Microbiol. Lett. 2: 323-326.
LEVI, c., C. SANCHEZ-WAS and P. SCHAEFFER, 1980 Formation de bactiries diploides stables par fusion de protoplastes de Bacillus subtilis et effet d e mutations rec- sur les produits d e fusion formes. C.R. Acad. Sci. Ser. D 291: 67-70.
L~vI-MEYRUEIS, C., and C. SANCHEZ-RIVAS, 1984 Complementation and genetic inactivation: two alternative mechanisms leading to prototrophy in diploid bacterial clones. Mol. Gen. Genet. 196:
488-493.
LOPEZ, F., 1985 Utilisation d’un plasmide pour l’itude de l’expres- 114: 273-286.
126 487-496.
Epigenetic Inactivation in B. subtilis 88 1
sion d u gknome diploide des Ncd stables issus de la fusion d e protoplastes de Bacillus subtilis. These d e Docteur 3eme cycle Microbiologie. UniversitC Paris-Sud, centre d’Orsay, France. LOPEZ, F., N. GUILLEN and L. HIRSCHBEIN, 1986 Further character-
ization of the inactive chromosome from Bacillus subtilis stabi- lized non-complementing diploids, pp. 73-86 in Bacillus Molecu- lar Genetics and Biotechnology Applications, edited by A. T. GANESAN and J. A. HOCH. Academic Press, New York.
LYON, M. F., 1993 Epigenetic inheritance in mammals. Trends Genet. 9: 123-128.
LYNCH, A. S., and J. C. WANC, 1995 SopBprotein-mediated silencing of genes linked to the sopC locus of Escherichia coli F plasmid. Proc. Natl. Acad. Sci. USA 92: 1896-1900.
MAXIATIS, T., E. F. FRITSCH and J. SAMBROOK, 1982 Molecular Clon- ing: A Labmatoly Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
NIAUDET, B., and S. D. EHRIJCH, 1979 In vitro genetic labeling of
Bacillus subtilis cryptic plasmid pHV400. Plasmid 2: 48-58. SAIKI, R. K., 1990 Amplification ofgenomic DNA, pp. 13-20 in PCR
Protocols, edited by M. A. INNIS, D. H. GELFAND, J. J. SNINSKY and T. J. WHITE. Academic Press, New York.
SANCHEZ-RIVAS, C., and C. LEVI-MEYRUEIS, 1994 Ploidy of Bacillus subtilis exfusants: a controversy. Microbiol. 140: 1-2.
SANCHEZ-WAS, C . , C. LEVI-MEYRUEIS, FLAZARD-MONIER and P.
SCHAEFFER, 1982 Direct selection of complementing diploids from PEGinduced fusion of Bacillus subtilis protoplasts. Mol. Gen. Genet. 185: 329-333.
SCHAEFFER, P., and L. HIRSCHBEIN, 1985 Bacillus subtilic protoplast fusion, nucleoids and chromosome inactivation, p.77-88 in Fun- damental and Applied Aspects of Bacterial Spores, edited by G. J. DRING, D . J . E I .and G. W. GOUID. ~ Academic Press, London. SCHAEFFER, P., B. CAMI and R. D. HOTCHKISS, 1976 Fusion of bacte-
rial protoplasts. Proc. Natl. Acad. Sci. USA 73: 2151-2121. THAI.ER, D. S., J. R. ROTH and L. HIRSCHBEIN, 1990 Imprinting as
a mechanism for the control of gene expression, pp. 445-456 in The Bacterial Chromosome, edited by K. DRI.ICA and M. RILEY. American Society for Microbiology, Washington, DC.
ZAHLER, S. A, 1993 Temperate bacteriophages, pp. 831-841 in Bacil- lus Subtilis and Other Gram-Positive Bactm’a, edited by A. L. SO- NEPU’SHEIN, J. A. HOCH and R. LOSICK. American Society for Micro- biology, Washington, DC.