in extraocular muscle function
Kenneth E. Eakins and Ronald L. Katz
The purpose of this study was to determine the possible role of the autonomic nervous system in the function of mammalian extraocular muscle. Experiments were carried out on the cat anesthetized with pentobarbital. Stimulation of the cervical sympathetic nerve and the injec-tion of epinephrine increased the tension of the superior rectus muscle. These responses were unaffected by sympathetic ^-receptor blocking agents, potentiated by cocaine, and antagonized by sympathetic a-receptor blocking agents. Similar results were observed with the nictitating membrane. The superior rectus muscle and nictitating membrane differed in that atropine blocked the response of the nictitating membrane to epinephrine, but not that of the superior rectus muscle. The implications of these remits are discussed.
.he histological observations of Boeke1 and Wolter2 indicating that the striated muscle fibers of extraocular muscles have a double innervation, motor and auto-nomic, led Alpern and Wolter3 to advance the view that the slow vergence move-ments of the eye were under autonomic control. This aspect of the control of ex-traocular muscle activity has since received little scientific attention. For this presenta-tion, I would like to deal exclusively with some results we have obtained in our laboratory concerning the effect of cervical
From the Departments of Ophthalmology Research and Anesthesiology, College of Physicians and Surgeons, Columbia University, New York, N. Y.
Supported by Fight-for-Sight Grant-in-Aid G 303-C-2-C3 of the National Council to Combat Blindness, Inc., New York, and United States Public Health Service National Institutes of Health Grant GM 09069.
Figs. 1, 2, 5, 6, and 7 are reprinted from and with the permission of The Journal of Pharma-cology and Experimental Therapeutics. In press.
sympathetic stimulation and systemically administered epinephrine on extraocular muscle tension. No attempt will be made to assess the relative importance of the role of the parasympathetic nervous system in extraocular muscle function at this stage, except to point out that we have repeatedly observed in our laboratory that the con-tracture of the extraocular muscle pro-duced by acetylcholine is unaffected by atropine, which would seem to preclude the involvement of parasympathetic muscarinic receptors in the response.
In an earlier study,4 we observed that the intravenous injection of epinephrine in the cat resulted in a small but fairly well-maintained contracture of the superior and lateral rectus muscles. This observation was of interest to us in that, as long ago as 1930, Duke-Elder and Duke-Elder1 had drawn attention to the fact that mammalian extraocular muscles behave in many re-spects like denervated mammalian'skeletal muscle; for example, both the external mus-cles of the eye and chronically denervated leg muscles respond to acetylcholine with
a contracture, and, in contrast to the lack of effect of sympathetic amines on normal peripheral skeletal muscle, these com-pounds will produce a slowly developing reversible increase in resting tension sev-eral weeks after section of the motor nerve. Bowman and Raper in 1965° reported that the response of the chronically dener-vated tibialis anterior and soleus muscles of the cat to epinephrine corresponded on AhlquistV 1948 classification to a sympa-thetic /3-receptor effect since it was selec-tively blocked by sympathetic /3-receptor, but not by a-receptor antagonists.
In this presentation, we will describe some experiments that we have carried out to determine whether the increased tension of extraocular muscles observed after sys-temic administration of epinephrine repre-sents yet another similarity between extra-ocular and chronically denervated skeletal muscle. We have examined the response of the superior rectus muscle of the cat to epinephrine and sympathetic stimulation and its modification by a variety of agents. In addition, we determined the response of the nictitating membrane under these con-ditions to monitor the behavior of the orbit-al smooth muscle.
Methods
Cats of either sex, weighing between 2.5 and 3.5 kilograms, were used in these experiments. The animals were anesthetized with sodium penta-barbital (36 mg. per kilogram) given by
intra-peritoneal injection. The trachea was cannulated and artificial respiration employed. Arterial blood pressure was recorded from the right femoral arteiy with a Statham (P23Db) pressure trans-ducer coupled to a Grass Model 5 ink-writing polygraph.
The superior rectus of one eye was separated from the globe and a suture placed through the tendon, the thread then being connected to a Grass force displacement transducer (FT-03). Contractions of the ipsilateral nictitating mem-brane were recorded by a similar force displace-ment transducer. The animal's head was immo-bilized in a stereotaxic apparatus. The cornea of the test eye was incised and the eyeball eviscer-ated and ligatured. Intra-arterial injections were made via a polyethylene cannula inserted into the ipsilateral common carotid artery. Intravenous in-jections were made via a cannula inserted into the left femoral vein.
Nerve stimulation. For preganglionic stimula-tion, the sympathetic trunk was cut just above the sternum and laid on a pair of silver electrodes. For postganglionic stimulation, the superior cer-vical ganglion was exposed, and postganglionic nerve isolated and laid on a pair of silver elec-trodes. The preparation was prevented from dry-ing with warm liquid paraffin. Rectangular pulses with parameters determined in each experiment were delivered by a Grass S4 stimulator in con-junction with a stimulus isolation unit.
Drugs used in this study were 1-epinephrine chloride (Parke, Davis & Co.), phenoxybenza-mine hydrochloride (Dibenzyline, Smith, Kline & French Labs.), phentolamine (Regitine, Ciba Pharmaceutical Products, Inc.), pronethalol (Al-derlin, Ayerst Labs.), propanolol (Inderal, Ayerst Labs.), atropine sulfate (Burroughs Wellcome & Co.), and cocaine hydrochloride. The drugs were diluted in 0.9 per cent weight per volume saline. All doses refer to the. salts.
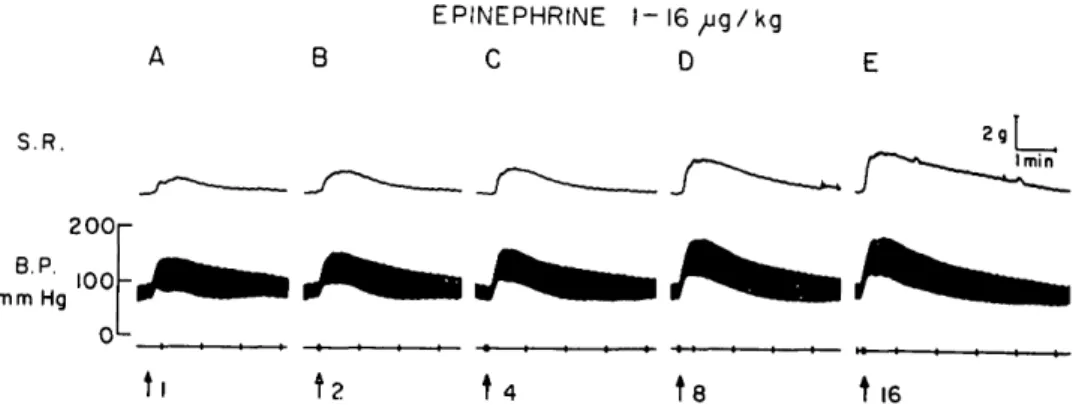
EPINEPHRINE t — 16 C D kg S.R. B.P. mm Hg 2OOr tl
t
t8 t 16Fig. 1. Cat anesthetized with pentobarbital. Effect of increasing intravenous doses of epi-nephrine on the superior rectus muscle (S.R.). Calibrations 2 g tension and 1 minute.
Results and discussion
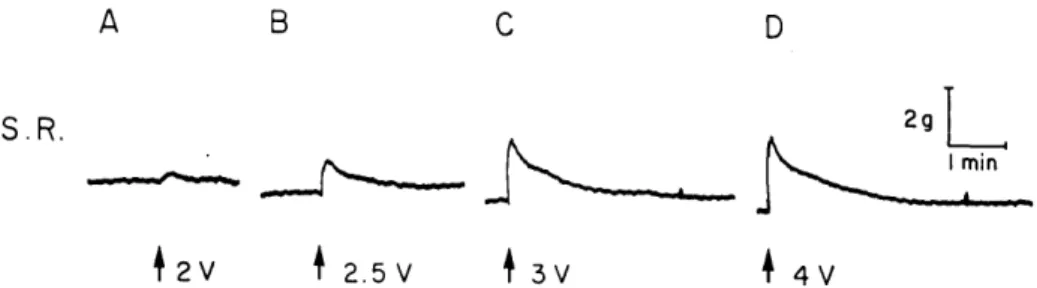
Intravenous injections of epinephrine (1 to 16 /xg per kilogram) were repeatedly found to produce an increase in the rest-ing tension of the superior rectus muscle. A typical result is illustrated in Fig. 1. In these experiments, injections were made every 5 to 10 minutes; with more frequent administration a marked decrease in the response was observed. The response of the extraocular muscle to epinephrine did not appear to be associated with cardiovascular changes, since an increase in muscle ten-sion was always noted, irrespective of whether the dose of epinephrine resulted in a rise or fall in the arterial blood pres-sure. Fig. 2 illustrates the voltage-depen-dent increase in tension of the superior rectus muscle to electrical stimulation of the ascending cervical sympathetic nerve carried out at a frequency of 20 cycles per second. It was of importance to determine whether these responses of the superior rectus muscle were the result of a direct action of sympathetic stimulation and epinephrine on this striated muscle or if the changes in extraocular muscle tension were secondary to an effect on intraorbital smooth muscle. Consequently, we com-pared the responses of both the superior rectus muscle and the nictitating mem-brane to these procedures in the presence of various compounds known to modify the response of smooth muscle. First of all, we studied the sympathetic a- and /^-receptor blocking agents. The ^-receptor blocking agents, pronethalol (5 mg. per kilogram
intravenously) and propanolol (1 mg. per kilogram intravenously), did not antagonize the response of either muscle to sympa-thetic stimulation or epinephrine. Ex-amples of these results are seen in Figs. 3 and 4. The doses of the blocking agents used in these experiments are well known to produce substantial sympathetic /3-re-ceptor blockade elsewhere. The increased pressor response to epinephrine after the dose of propanolol in Fig. 4 indicates a high degree of /^-receptor blockade in the ani-mal.
Substantial antagonism of the responses of both muscles to epinephrine and sym-pathetic stimulation was observed after treatment with the sympathetic a-receptor blocking agents, phenoxybenzamine (1 to 2 mg. per kilogram intravenously) and phentolamine (2 to 3 mg. per kilogram in-travenously). Fig. 5 illustrates the effect of an intravenous injection of 1 mg. per kilo-gram of phenoxybenzamine on the response of the nictitating membrane and superior rectus muscle to postganglionic sympa-thetic stimulation and intra-arterial injec-tion of 3 [xg of epinephrine. It can be seen that all the responses were markedly de-pressed by this sympathetic a-receptor blocking agent.
Epinephrine is well known to modify skeletal neuromuscular transmission in the cat (see Bowman and Rapers for refer-ences); it has not previously been demon-strated to alter resting tension in normally innervated mammalian striated muscles, •although chronically denervated
mammali-S.R.
2V 2.5 V 3V 4V
Fig. 2. Cat anesthetized with pentobarbital. Effect of electrical stimulation of the cervical sympathetic nerve on the superior rectus muscle (S.R.) 20 per second, 3 msec, duration, Calibrations 2 g tension and 1 minute .
N.M. 10 g I min S.R. ig Imin 15V
t
15V Pronethalol 5mg/kgFig. 3. Cat anesthetized with pentobarbital. Lack of eflect of pronethalol on the response of the superior rectus (S.R.) and nictitating membrane (N.M.) to sympathetic stimulation. Calibrations in g tension and minutes as indicated.
an striated muscle will respond to epine-phrine with a slowly developing reversible increase in resting tension."19> 10~13 It has been reported that the response of the chronically denervated tibialis anterior and soleus muscles corresponds on Ahl-quist's7 classification of catecholamine re-ceptive mechanisms to a sympathetic /?-receptor effect, since the response was most effectively produced by Levisoprenaline and was selectively blocked by the sympa-thetic /3-receptor blocking agents, dichlori-soproterenol and pronethalol.0 The results obtained after systemic administration of epinephrine in these experiments indicate that the response of the striated superior rectus muscle to the catecholamine differs from the response obtained from chron-ically denervated mammalian skeletal mus-cles, since the response was unaffected by the sympathetic /3-receptor blocking agents, but was abolished by the sympathetic a-receptor blocking agents.
A N.M. S . R . 200r BP mm Hg 100 0L tAd
B
t Ad Propanalol I mg/kgFig. 4. Cat anesthetized with pentobarbital. Response of the superior rectus (S.R.) and nictitat-ing membrane (N.M.) to epinephrine (Ad) before (A) and after (B) 1 mg. per kilogram of propanolol. All drugs given intravenously. Note increase in effect of epinephrine on arterial blood pressure (B.P.) after the propanolol. Calibrations in g tension and minutes as indicated.
S.R. I min N.M. Imin Postgang.stim.
•
Ad Postgong.stim.•
Ad Phenoxybenzamine Img/kgFig. 5. Cat anesthetized with pentobarbitone. Inhibition of the responses of both muscles to sympathetic stimulation and intravenous epinephrine (Ad) by phenoxybenzamine. (A) Control responses of superior rectus (S.R.) and nictitating membrane (N.M.); (B) responses after phen-oxybenzamine. Calibrations in g tension and minutes as indicated.
A B D N.M. S.R. Pregang stim.
t
Ad Pregang Adt t
stim. COCAINE 5mg/kgFig. 6. Cat anesthetized with pentobarbital. Effect of cocaine on the responses of the superior rectus (S.R.) and nictitating membrane (N.M.) to preganglionic' sympathetic stimulation and epinephrine (Ad) (A) and (B) control responses; (C) and (D) responses after cocaine. All drugs injected intravenously. Calibrations in g tension and minutes as indicated.
The effect of cocaine (5 mg. per kilo-gram intravenously) on these responses is seen in Fig. 6. Notice that the responses of both muscles to preganglionic sympathetic stimulation were slightly depressed, but that the epinephrine responses were defi-nitely enhanced. Further studies showed
that these doses of cocaine always poten-tiated the effects of postganglionic sympa-thetic stimulation on both muscle systems. This difference between the effect of cocaine on the responses to pre- and post-ganglionic sympathetic stimulation has been reported by other workers, the
im-portant point here being that both muscles responded in the same way. Lastly, we examined the effect of atropine (1 to 2 mg. per kilogram intravenously) on these responses. A typical result is seen in Fig. 7. It can be observed that the injection of 1 mg. per kilogram of atropine differen-tiated the responses to some extent. The response of the superior rectus muscle and nictitating membrane to sympathetic stimu-lation and the response of the nictitating membrane to epinephrine were depressed by atropine, whereas the response of the superior rectus muscle to epinephrine was unaffected.
The identical behavior of the nictitating membrane and superior rectus muscle to sympathetic stimulation through all the procedures reported here raises the possi-bility that a part of the response of the superior rectus muscle produced by stimu-lation may be a pseudocontracture sec-ondary to contraction of intraorbital smooth muscle. However, it is possible that the sympathetic nervous system may be in-volved with something other than the con-trol of the vasculature in the extraocular muscles. We hope that studies now in progress (in collaboration with Dr. R.
Barrett of Columbia University), utilizing the fluorescence technique developed by Hillarp and Falck for the histological local-ization of catecholamines in tissues, will yield more precise information on this problem. In addition, we also have to rule out the possibility that the extraocular mus-cles, already rather unusual in terms of their structure as compared with other skeletal muscles,14'15 may possess smooth muscle elements which may be responsible for the present observations.
It is also difficult to explain the response of the superior rectus muscle to epineph-rine entirely in terms of an effect on orbital smooth muscle in view of the differential action of atropine. It has been demon-strated before that atropine is capable of antagonizing the response of the smooth muscle cells of the nictitating membrane to both epinephrine and sympathetic stimu-lation.10' 17 Thus, the observation that atropine did not reduce the response of the superior rectus muscle to epinephrine argues against the total extraocular muscle response being secondary to changes in tone of the intraorbital smooth muscle.
This effect of epinephrine on the superior rectus muscle may be related to its
anti-B
S.R.
N.M. 5g
Imin
•
t A dPostgang stim. Postgang stim. Atropine
I mg/kg
Ad
Fig. 7. Cat anesthetized with pentobarbital. Differential action of atropine. (A) Control re-sponses of the superior rectus (S.R.) and nictitating membrane (N.M.) to sympathetic stimula-tion and intra-arterial epinephrine (1 Hg); (B) responses after intravenous atropine. Note all responses depressed except the response of S.R. to epinephrine. Calibrations in g tension and minutes as indicated.
curare action in partially curarized striated muscles.18'21 In common with the present response of the superior rectus muscle, the anticurare action is blocked by sympathetic a-receptor blocking agents.21"23 This anti-curare effect of epinephrine is thought to be due to an increase in acetylcholine release resulting from a hyperpolarizing action of epinephrine on motor nerve end-ings.-'1- 25 Thus, mobilization of acetyl-choline from prejunctional storage sites in the superior rectus muscle by epinephrine would result in an increase in tension in the muscle, the acetylcholine most prob-ably affecting the "slow" or multiply inner-vated muscle fibers. Since the contracture of the superior rectus muscle produced by acetylcholine is unaffected by atropine this would also explain the lack of effect of atropine on the response to epinephrine described in this paper. However, these experiments do not rule out two further possibilities which could also explain the response of the extraocular muscle to epinephrine: (1) the presence of smooth muscle elements within the extraocular muscle, as suggested above, and (2) that epinephrine may be acting directly on the neuromuscular junctions, most probably the multiple endplates of the "slow" fibers. We hope that work currently in progress in our laboratory will yield answers to some of these problems.
REFERENCES
.1. Boeke, J.: Die Beziehungen cler Nervenfasern zu den Bindegewebselementen und Tastzel-len, Ztschr. mikr.-anat. Forsch. 4: 448, 1926. 2. Wolter, J. R.: Morphology of the sensory
nerve apparatus in the striated muscle of the human eye, Arch. Ophth. 53: 201, 1955. 3. Alpern, M., and Wolter, J. R.: The relation
of horizontal saccadic and vergence move-ments, Arch. Ophth. 56: 685, 1956. 4. Eakins, K. E., and Katz, R. L.: The action
of succinylcholine on the tension of extra-ocular muscle, Brit. J. Pharmacol. 26: 205, 1966.
5. Duke-Elder, W. S., and Duke-Elder, P. M.: The contraction of the extrinsic muscles of the eye by choline and nicotine, Proc. Roy. Soc. (B). 107: 232, 1930.
6. Bowman, W. C , and Raper, C.: The effects of sympathomimetic amines on chronically denervated skeletal muscles, Brit. J. Pharma-col. 24: 98, 1965.
7. Ahlquist, R. P.: A study of the adrenotropic receptors, Am. J. Physiol. 153: 586, 1948. 8. Bowman, W. C , and Raper, C : Effects of
sympathomimetic amines on neuromuscular transmission, Brit. J. Pharmacol. 27: 313, 1966.
9. Euler, U. S. von, and Gaddum, J. H.: Pseudo-motor contractures after degeneration of the facial nerve, J. Physiol. (Lond) 73: 54, 1931. 10. Biilbring, E., and Bum, J. H.: The
Sherring-ton phenomenon, J. Physiol. (Lond) 86: 61, 1936.
11. Luco, J. V., and Sanchez, P.: Spontaneous activity and contractile responses to adrena-line of denervated auricular muscles, Acta physiol. latinoam. 6: 171, 1956.
12. Luco, J. V., and Sanchez, P.: The effect of adrenaline and noradrenaline on the activity of denervated skeletal muscles. Antagonism between curare and adrenaline-like substances, in Curare and curare-like agents, Amsterdam, 1959, Elsevier Publishing Company, pp. 405-408.
13. Bowman, W. C , and Zaimis, E.: The action of adrenaline, noradrenaline and isoprenaline on the denervated mammalian muscle, ]. Physiol. (Lond.) 158: 24, 1961.
14. Hess, A., and Pilar, C : Slow fibres in the extraocular muscles of the cat, J. Physiol.
(Lond.) 169: 780, 1963.
15. Dietert, S. E.: The demonstration of different types of muscle fibres in human extraocular muscle by electron microscopy and
cholin-esterase staining, INVEST. OPHTH. 4: 51, 1965.
16. Cervoni, P., West, T. C , and Fink, L. D.: Autonomic postganglionic innervation of the nictitating membrane of the cat, J. Pharmacol. & Exper. Therap. 116: 90, 1956.
17. D'Alena, P., and Featherstone, R. M.: Effects of atropine and related alkaloids on epineph-rine induced contractions of the cat nic-titating membrane, J. Pharmacol. & Exper. Therap. 149: 351, 1965.
18. Panella, A.: Action due principe actif sur-renal sur la fatigue musculaire, Arch. ital. biol. 48: 430, 1907.
19. Rosenblueth, A., Lindsley, D. B., and Mori-son, R. S.: A study of some decurarizing sub-stances, Am. J. Physiol. 115: 53, 1936. 20. Wilson, A. T., and Wright, S.: Anticurare
effect of potassium and some other substances, Quart. J. Exper. Physiol. 26: 127, 1937. 21. Brown, G. L., Goffart, M., and Dias, M. V.:
The effects of adrenaline and of sympathetic stimulation on the demarcation potential of
mammalian skeletal muscle, J. Physiol. (Lond.) I l l : 184, 1950.
22. Maddock, W. O., Rankin, V. M., and You-mans, W. B.: Prevention of the anticurare action of epinephrine by dibenamine, Proc. Soc. Exper. Biol. & Med. 67: 151, 1948. 23. Bowman, W. C , Goldberg, A. A. J., and
Raper, C : A comparison between the effects of a tetanus and the effects of sympathomi-metric amines on fast- and slow-contracting
mammalian muscles, Brit. J. Pharmacol. 19: 464, 1962.
24. Krnjevic, K., and Miledi, R.: Some effects produced by adrenaline upon neuromuscular propagation, J. Physiol. (Lond.) 141: 291, 1958.
25. Krnjevic, K., and Miledi, R.: Presynaptic failure of neuromuscular propagation in rats, J. Physiol. (Lond.) 149: 1, 1959.