STRUCTURAL ANALYSIS OF THE BILIARY GLYCOPROTEIN (BGP) BINDING SITE
by
Ana M aria M iranda Botelho Teixeira
A thesis subm itted for the degree of Doctor of Philosophy, U niversity of London.
I.C.R.F. D epartm ent of Medical Onclogy St. Bartholom ew’s H ospital Medical College
L ondon
ProQ uest Number: 10017415
All rights reserved
INFORMATION TO ALL U SE R S
The quality of this reproduction is d ep en d en t upon the quality of the copy subm itted. In the unlikely even t that the author did not sen d a com plete manuscript
and there are m issing p a g e s, th e se will be noted. Also, if material had to be rem oved, a note will indicate the deletion.
uest.
ProQ uest 10017415
Published by ProQ uest LLC(2016). Copyright of the Dissertation is held by the Author. All rights reserved.
This work is protected against unauthorized copying under Title 17, United S ta tes C ode. Microform Edition © ProQ uest LLC.
ProQ uest LLC
789 East E isenhow er Parkway P.O. Box 1346
ABSTRACT
The Biliary G lycoprotein (BGP) gene is a m em ber of th e C arcino- em bryonic A ntigen (CEA) Gene Family. It codes for at least 10 isoforms obtained by differential splicing of the BGP gene. Structurally they are m em bers of the Im m unoglobulin su p er fam ily (IgSF) an d function as hom otypic adhesion molecules. It is clustered as CD66a an d is m ainly present in leukocytes and in th eep ith eliu m o f bile caniculi an d colon . C H O tran sfectan ts expressing the BGPc isoform an d soluble fusion p ro tein chim eras (N-Fc, N A IB I-F c an d NA1B1A2-Fc) w ere used in b in d in g assays w here b in d in g of these tran sfectan ts to the soluble dom ain proteins w as tested. They w ere show n to bind in a hom ophilic m an n er suggesting th at the N -term inal dom ain w as in volved in the ad h esio n process. BGP isoform s w ith an ex tracellu lar N -term in al dom ain only, followed by a transm em brane dom ain and a 72 or 9 am ino acid cytoplasmic tail (BGPx, BGPx') have also been described. In order to ascertain the N -N dom ain n atu re of this adhesion a chim eric construct equivalent to the BGPx' isoform w as constructed by a tw o step PGR m eth o d using prim ers containing the sequences of the en d of the N- term in al d o m ain follow ed by the b eg in n in g of the tran sm em b ran e dom ain, together w ith prim ers for either the beginning of the BGPc sequence or the end of the cytoplasmic tail. Sequencing of this construct m atched the predicted sequence and a stable CHO-BGPx' cell line was established. Expression of this isoform at the cells surface w as confirmed by im m unofluorescence staining and w estern blot. A dhesion assays show ed th at this cell line w as able to adhere to the N-Fc construct confirm ing the N -N term in al n a tu re of this adhesion. To fu rth er localise the binding site a set of peptides spaning the C, C and FG P strands of the N -term inal dom ain w ere tested for their ability to inhibit adhesion. Both the C, C C and FG peptides w ere able to partialy inhibit adhesion w ith the FG peptide being the m ost effective. This peptide w as also able, at a higher concentration, to prom ote high levels of adhesion, using either transfected CHO-BGPc or un transfected CH O cells. These results indicate that this peptide was able to activate adhesion molecules present in the CHO cells probably integrins.
ACKNOWLEDGEMENTS
Projects like this can never be accom plished w ith o u t a w id e ran g e of su p p o rt. H ere I g ratefu lly acknow ledge the people w h o m ad e this project possible: in particular. Professors D avid Linch, Bryan Young and D r Suzanne W att for their guidance and supervision. I also th an k the P o rtu g u ese Junta N acional de Investigaçâo C ientifica e Tecnologica (JNICT) an d the Im p erial C ancer R esearch F u n d (ICRF) fo r th eir financial su p p o rt. This project w as u n d e rta k en at the ICRF M edical Oncology U nit at St. Bartholom ew's H ospital Medicall College.
M any thanks also to Dr. Paul Bates for his valuable discussions an d advice on the structure of BGP an d to Dr. Lorna Stew art for technical advice on the BGPx' vector and site directed m utagenesis. I am also m ost grateful to Drs Steve Gschm eissner an d N asser H ajibageri for the electron m icroscopy w ork, D ereck Davis an d C lare H ughes for cell sorting, M aggie Stubbs for protein purification, Peter W ilson for his help w ith the statistical analysis and Drs. Ian G oldsm ith and G erard Evan for oligonucleotide and peptide synthesis respectively.
I w o u ld also like to thank all m y colleagues in M edical O ncology for their technical advice and friendship, in p articu lar Tracy C haplin an d Joanne M eerabux, M ary Cotter, w ho assisted m e w ith the diagram s and spelling corrections, and Keith A dam s, w ho helped w ith the art work. From Portugal I w o u ld like to acknow ledge the su p p o rt of Professor M aria de Sousa for her confidence in m y scientific capabilities, and also Dr. Isabel Barbosa for her continuous support.
LIST OF H G U R E S
C hapter I
1.1 Schematic representation of the a and P integrin
subunits dom ain organisation. 22
1.2 Schematic representation of the cadherin dom ain
organisation. 24
1.3 Schematic representation of the selectins dom ain
organisation. 26
1.4 Schematic representation of the sLex carbohydrate
m oiety. 28
1.5 Ribbon m odel of the im m unoglobulin fold. 30 1.6 Schematic representation of the chrom osom al
organisation of the CEA family genes. 36 1.7 Schematic representation of the dom ain
organisation of the CEA subgroup. 38
1.8 Schematic representation of the different BGP
isoform s dom ain organisation. 46
Chapter II
2.1 Diagram of the pIG vector. 54
2.2 Schematic representation of the truncated form s of
the soluble BGP fusion proteins. 55
2.3 O utline of the m ethod used for the construction of
the BGPx’cDNA. 65
2.4 O utline of the m ethod used for site directed
m utagenesis of the BGP N -term inal dom ain. 70 2.5 Schematic representation of the adhesion assay
Chapter III
3.1 Binding of anti-CEA family antibodies to
CHO-BGPc transfectants and PMN. 8 6
3.2 Binding of CD66 and CD67 defined antibodies to
CHO, CHO-BGPc and PMN cells. 87
3.3 W estern blot analysis of the CHO-BGPc lysate. 8 8 3.4 Electron m icroscopy photographs show ing
localisation of BGPc to contact sites in aggregating
CHO-BGPc cells. 9 0
3.5 Im m unofluorescence staining of CHO-BGPc cells. 91 3.6 Protein gel electrophoresis of the labelled
soluble BGP-Fc dom ain fusion proteins. 93 3.7 Antibody binding to the BGPc-Fc soluble dom ain
fusion proteins. 9 4
3.8 Binding of several anti-CEA MAbs to the CD33-Fc
fusion protein. 9 5
3.9 Fluorescence histogram s of antibody stained CHO- BGPc and CHO-CD34 cells lifted w ith either trypsin
or EDTA. 9 9
3.10 Q uantitative binding of CHO-BGPc and CHO-CD34 transfectants to the BGPc-Fc soluble dom ain fusion
proteins at 37°C. 100
3.11 Q uantitative binding of CHO-BGPc and CHO-CD34 cells to the BGPc-Fc soluble dom ain fusion proteins
at 4 0c . 101
3.12 Time course of adhesion betw een the CHO-BGPc
cells and the BGP-Fc [NA1B1A2] fusion protein. 1Q2 3.13 Q uantitative binding of CHO-BGPc and CHO-CD34
3.14 Q uantitative binding of CHO-BGPc cells to the
BGF[N-Fc] soluble dom ain fusion protein in Puck's m edium containing EDTA, Ca^+, Mg2+ or both Ca2+
and Mg2+. ^04
C hapter IV
4.1 Agarose gel show ing the PCR products obtained
during the construction of the BGPx' cDNA. 109
4.2 BGPx' DNA and am inoacid sequence. 110
4.3 Fluorescence histogram s of the 3 cell sorting cycles
used to establish the CHO-BGPx' cell line. m 4.4 Binding of the CD66 MoAbs from the VII Leucocyte
W orkshop collection to the CHO-BGPx' cells. H2 4.5 Im m unoblot analysis of the BGPc,
CHO-BGPx', CHO-CD34 and CHO cell lysates. 113
4.6 Confocal microscopy photographs of
im m unofluorescence stained CHO-BGPx' cells. 11 4 4.7 Q uantitative binding of CHO-BGPx' and CHO cells
to the BGPc-Fc soluble dom ain fusion proteins at
3 7 0c . 116
4.8 Time course of adhesion betw een the CHO-BGPx'
cells and the BGP[N-Fc] dom ain fusion protein. 1 1 7 4.9 Q uantitative binding of CHO-BGPx' and CHO cells
to the BGPc-Fc soluble dom ain fusion proteins at
either 37^C or 4oC. 228
4.10 Q uantitative binding of CHO-BGPx' and CHO cells to the BGPc-Fc soluble dom ain fusion proteins at
37°C and in the presence of EDTA. 219
C hapter V
5.2 Inhibition of binding of CHO-BGPx' cells to BGP [N-Fc] soluble dom ain fusion protein by 3 different
BGP peptides. 126
5.3 Binding of CHO-BGPc cells to the BGP-Fc soluble dom ain fusion protein in the presence of 4
different concentrations of Peptide FG. i2 7 5.4 Binding of CHO-BGPc cells to the BGP-Fc soluble
dom ain fusion protein or to BSA in the presence
of a high concentration of Peptide FG. 129 5.5 Binding of CHO cells to the BGP-Fc soluble dom ain
fusion protein or to BSA in the presence of a high
concentration of Peptide FG. 130
5.6 Blocking of BGP hom ophilic adhesion by MoAbs
HD-11 and AH-11. 131
C hapter VI
6.1 Aminoacid sequence alignm ent of BGP and rat 136 CD2 N -term inal dom ains.
6 .2 Agarose gel show ing the PCR products obtained during the production of the m utated BGP cDNA
constructs. I3 7
6.3 Sequence trace data for the m utated BGP cDNA
constructs. 138-40
6.4 N -term inal dom ain am inoacid sequence alignm ent
of the m utated BGP[NA1B1]-Fc fusion proteins. 1 4 2 6.5 Binding of MoAbs 3B10, AD-11 and AH-11 to the
BGP[NA1B1]-Fc m utated fusion proteins. 143-44 6 .6 Binding of CHO-BGPc cells to the m utated
LIST OF TABLES
C hapter I
1.1 The CEA gene family m embers. 37
1.2 The rodent CEA-related gene family. 40
Chapter II
2.1 Restriction enzym es and buffers used in DNA
digestion. 58
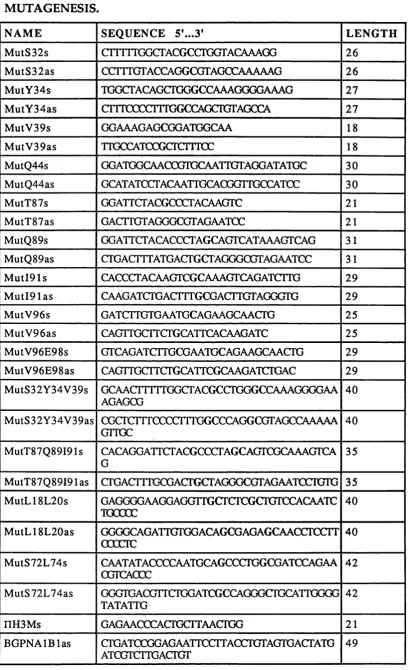
2.2 List of prim ers used in the production of the BGPx’
splice variant. 62
2.3 List of prim ers used in PCR screening and / o r
sequencing. 64
2.4 List of prim ers used for the site directed
m utagenesis 69
2.5 A ntibodies used in confocal m icroscopy 76 2.6 A m inoacid sequence of the N -term inal dom ain
BGP peptides. 82
Chapter III
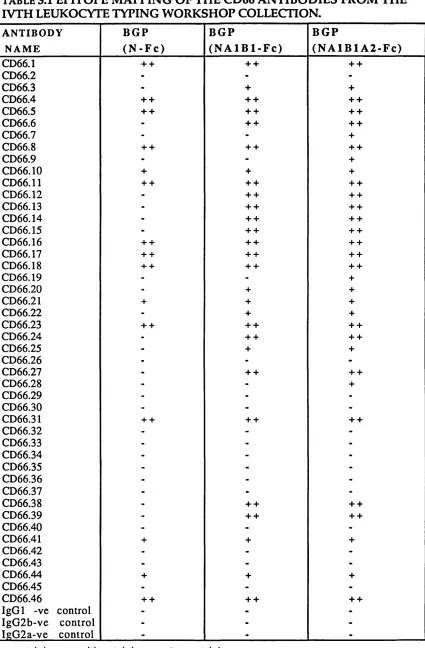
3.1 Epitope m apping of the CD66 antibodies from the
IVth leukocyte typing w orkshop collection. 96 C hapter V
5.1 Statistical analysis of the peptide inhibition
adhesion assays. 125
C hapter VI
6.1 Statistical analysis of the adhesion to the m utated
ABBREVIATIONS
ALL Acute lym phoblastic leukaem ia
AML Acute m yeloid leukaem ia
BGP Biliary glycoprotein
Bgp
Mouse biliary glycoproteinbp base pair
BSA Bovine serum album in
C C onstant
cDNA com plem entary DNA
C-CAM cellular cell adhesion m olecule
CAMs Cell adhesion molecules
CD Cluster of differentiation
CEA Carcinoem bryonic antigen
COM CEA group m em ber
CHO Chinese ham ster ovary
CML Chronic m yelogenous leukaem ia
CRD C arbohydrate recognition dom ain
dATP 2'-deoxyadenosine-5'-triphosphate
dCTP 2’-deoxycytidine-5'-triphosphate
DEAE-Dextran D iethyl-am ino-ethyl dextran
dCTP 2'-deoxyguanidine-5'-triphosphate
DMEM Dulbecco's m im im al essential m ed iu m
DMSO D im ethyl sulphoxide
dXTP 2'-deoxytim idine-5'-triphosphate
DTT D ith io th reito l
BCD E-cadherin EDTA EGF E-selectin EtBr FACS ECS FITC FN GlyCAM GPI Hams-FlO HD HIV ICAM Ig IgC2 IgSF IgV kb kDa LB LFA-1 LMP L-selectin M
Extracellular dom ain Endothelial cadherin
Fthylenodiam inotetra acetic acid Epiderm al grow th factor
Endothelial selectin E thidium brom ide
Fluorescence activated cell sorter Foetal calf serum
Fluoresceinyl isothiocyanate Fibronectin
G lycoprotein cell adhesion m olecule G lycosyl-phosphatidyl-inosytol
N u trien t m ixture H am 's-form ulation 10 H odgkin's disease
H um an im m unodificiency virus Intercellular cell adhesion m olecule Im m u n o g lo b u lin
Im m unoglobulin constant like dom ain type 2 Im m unoglobulin su p er fam ily
Im m unoglobulin variable like dom ain kilobase pair
kilo D alton L-broth
Leukocyte function associated m olecule 1 Low m elting p o in t
m M m illi m olar
p.M micro m olar
M AdCAM Mucosal adressin cell adhesion m olecule
MAG Myelin associated glycoprotein
pME p-m ercaptoethanol
M oAb M onoclonal antibody
m R N A m essenger RNA
N C A Non-specific crossreactive antigen
N -C ad h erin N eu ral cadherin
N C A M N eural cell adhesion m olecule
n an o g ram
NG S N orm al goat serum
N K N atu ral killer
PBS Phosphate buffered saline
P-cadherin Platelet cadherin
PCR polym erase chain reaction
PECAM-1 Platelet endothelial cell adhesion m olecule
P-selectin Platelet selectin
PSG Pregnancy specific glycoproteins
Ph Philadelphia chrom osom e
PM A Phorbo-12-m yristate-l 3-acetate
PM N Polym orphonuclear cells
PMSF phenyl m ethyl sulphonyl fluoride
R N ase E. coli ribonuclease A
rp m revolutions p er m inute
SCR Short consensus repeats
SDS-PAGE Sodium dodecyl sulphate polyacrylam ide gel
Taq Therm us aquaticus
TBE Tris borate buffer
T -cadherin T runcated cadherin
TEMED N ,N ,N ,N -T etram eth y leth y len ed iam in e
V Variable
VCAM vascular cell adhesion m olecule
VLA Very late antigen
U V U ltra violet light
WBC W hite blood counts
TABLE OF CONTENTS
ABSTRACT...2
ACKNOWLEDGEMENTS... 3
U S T O F n C U R E S... 5
U ST OF TABLES... 9
ABBREVIATIONS... 10
CHAPTER I 1.1. INTRODUCTION... 20
1.2. ADHESION MOLECULES FAMILIES...20
1.2.1. The integrin fam ily...20
1.2.2. The Cadherin fam ily... 23
1.2.3. The selectin fam ily...25
1.2.4. The im m unoglobulin super fam ily...28
1.3. THE CARCINOEMBRYONIC ANTIGEN (CEA) GENE FAMILY... 32
1.3.1. Historical perspective... 32
1.3.2. The CEA family subgroups... 33
1.3.3. The rodent CEA gene fam ily... 39
1.3.4. Tissue and cell distribution of the CEA subgroup proteins...41
1.3.5. The CEA related molecules as adhesion m olecules...42
1.4. THE BILIARY GLYCOPROTEIN (BGP) MOLECULES... 44
1.4.1. The BGP isoforms... 44
1.4.2. Possible functional roles of BGP... 45
1.4.3. BGP in m alignancy... 47
1.5. AIMS OF THIS THESIS... 48
CHAPTER II 2.1 MATERIALS...50
2.2.1. GENE CONSTRUCTS AND SEQUENCE... 51
2.2.1.1. Expression cloning and seq u en cin g... 51
2.2.1.2. Production o f solub le BGPc d om ain constructs...52
2.2.1.3. Production o f the solu ble CD33-Fc d efin ed m o lecu le...53
2.2.1.4. P roduction o f CHO-BGPc and C H O -C D 34 stable transfectants... 56
2.2.2. PREPARATION OF PLASMID cDNA ON CSCL GRADIENTS...56
2.2.2.1. Production o f com petent bacteria... 56
2.2.2.2. Transform ation o f MC1061 bacteria...57
2.2.2.3. CsCl plasm id preparations... 57
2.2.3. DIGESTION WITH RESTRICTION ENZYMES...58
2.2.4. PRODUCTION OF THE SOLUBLE BPC DOMAIN FUSION PROTEINS... 59
2.2.4.1. Transfection o f COS 7 cells... 59
2.2.4.2. Labelling o f the fu sion p rotein s... 59
2.2.4.3. Protein A sepharose pu rification... 60
2.2.4.4. SDS g el electrophoresis...60
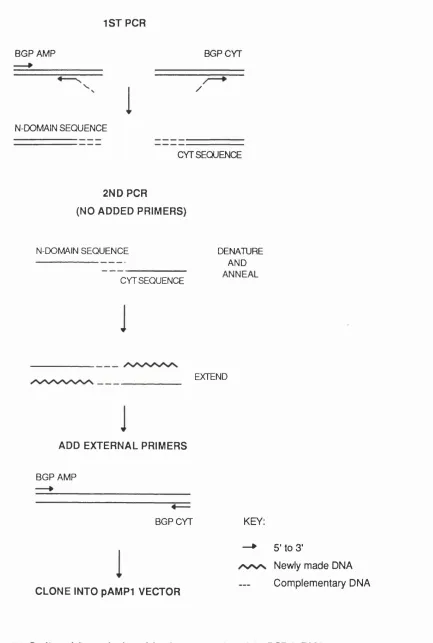
2.2.5. CONSTRUCTION OF THE BGPx* SPLICE VARIANT BY PCR... 61
2.2.5.1. Prim ers and PCR con d ition s...61
2.2.5.2. C lonin g into the pA M P 1 vector... 63
2.2.5.3. C olon y screen ing... 63
2.2.5.4. S eq u en cin g... 64
2.2.5.5. C lonin g into the exp ression vector p C D N A I/A m p ...6 6 2.2.5.6. Production o f a CHO-BGPx* stable cell lin e ...67
2.2.6. SITE DIRECTED MUTAGENESIS OF THE BGP N-TERMINAL DOMAIN 67 2.2.6.1. P roduction o f the d ifferent m u tan ts u sin g a PCR b a sed m eth o d ... 67
2.2.6.2. M utant analysis and seq u en cin g ... 78
2.2.7. CELL SEPARATION AND GROWTH OF CELL LINES... 71
2.2.9. FLOW CYTOMETRY... 72
2.2.10. WESTERN BLOTTING OF THE CHO CELL LINES... 72
2.2.10.1 CHO-BGPc and CHO-CD34 lysate preparations... 72
2.2.10.2 L ysate im m u n op récip itation ... 73
2.2.10.3. SDS-PAGE gel electrophoresis and w estern b lo t...73
2.2.11. ELECTRON MICROSCOPY... 74
2.2.11.1. Pre-Em bedding L abelling... 74
2.2.11.2. Post-E m bedding L abelling... 75
2.2.12. CONFOCAL MICROSCOPY OF THE CHO-BGPc, CHO-BGPx’ AND CHO-CD34 TRANSFECTANTS...76
2.2.13. EPITOPE MAPPING USING SOLUBLE BGPc-Fc DOMAIN FUSION PROTEINS... 77
2.2.13.1. Protein and concentration determ in ation ... 77
2.2.13.2. ELISA A ssa y ... 77
2.2.13.3. BGP [NA1B1]-Fc m utant fu sion proteins ELISA...78
2.2.14. 3h-THYMIDINE LABELLING OF CELLS AND ADHESION ASSAYS USING IMMOBILISED Fc DOMAIN FUSION PROTEINS... 79
2.2.14.1. Preparation o f the ad h esion assay w e lls ...79
2.2.14.2. Labelling o f the CHO transfected cells... 79
2.2.14.3. A d h esion assay... 79
2.2.14.4. C H O -B G P x’ a d h e sio n to the BGPc so lu b le d o m a in fu sio n p ro tein s... 81
2.2.14.5. Inhibition o f BGPc h om oph ilic ad h esio n b y p e p tid e s... 82
2.2.14.6. In h ib itio n o f BGP h o m o p h ilic a d h e s io n b y C D6 6 a n tib o d ies... 82
2.2.14.7. C H O -BG Pc a d h esio n to th e m u ta n t B G P[N A 1B 1]-Fc fu sio n p ro tein s... 83
CHAPTER III INTRODUCTION...85
3.1. P h en o ty p in g of the CHO-BGPc stable tran sfectan ts an d
polym orphonuclear cells (PMN)... 85
3.2. W estern bloting of the CHO-BGPc and CHO cell line lysates...88
3.3. Electron microscopy of the CHO-BGPc cells... 89
3.4. Confocal exam ination of CHO-BGPc cells stained w ith the YTH.71.3.2 a n tib o d y ... 89
3.5. P ro d u ctio n an d P rotein A Sepharose p u rificatio n of th e BGPc soluble dom ain fusion proteins... 92
3.6. E pitope m ap p in g of the BGPc-Fc soluble d o m ain fusion p ro tein s... 92
3.7. ADHESION OF THE CHO-BGPC CELLS TO THE IMMOBILIZED BGPc-Fc FUSION PROTEINS... 97
3.7.1. In v o lv e m e n t of th e N -te rm in a l d o m a in in BGP hom ophilic adhesion properties...97
3.7.2. BGPc hom otypic adhesion tem perature d e p e n d e n c y ... 98
3.7.3. BGPc adhesion time course... 98
3.7.4. BGPc adhesion cation dependency... 98
DISCUSSION... 105
CHAPTER IV INTRODUCTION... 108
RESULTS... 108
4.1. Construction of the BGPx' splice variant using PCR... 108
4.2. Characterization of the CHO-BGPx' stable cell line... 110
4.3. FUNCTIONAL ANALYSIS OF THE BGPx' ISOFORM... 115
4.3.1. A dhesion of the CHO-BGPx' cells to the BGP soluble dom ain fusion p ro tein s... 115
4.3.2. BGPx' time course of adhesion...115
4.3.3. Tem perature dependency of adhesion... 117
4.3.4. BGPx' cation dependency of adhesion... 119
CHAPTER V
INTRODUCTION...123
RESULTS... 123
5.1. INHIBITION OF BGP HOMOPHILIC ADHESION BY SELECTED PEPTIDES... 123
5.1.1. P ep tid e selection and purification... 123
5.1.2. P ep tid e inh ibition o f BGP h o m o p h ilic a d h esio n ...124
5.2. Integrin activation b y the FG p ep tid e ... 128
5.3. A n tib o d y inh ibition o f BGP h o m o p h ilic a d h esio n ... 128
DISCUSSION... 132
CHAPTER VI INTRODUCTION... 135
RESULTS... 135
6.1. M utant selection and p rod u ction ... 135
6.2. Purification and analysis the o f m utant chim eric p ro tein s...141
6.3. CHO-BGPc ad h esio n to the m u tated BGP [N A IB I-F c] fu sio n p r o te in s...144
DISCUSSION... 149
CHAPTER VII 7.1. GENERAL DISCUSSION... 152
APPENDIX 1 ...163
APPENDIX II...168
APPENDIX m ...175
APPENDIX IV...178
APPENDIX V ...179
CHAPTER I:
1. INTRODUCTION 1.1 Introduction.
This chapter starts w ith a brief review of the m ain adhesion m olecule families as a setting for the m ore detailed description of the CEA family. It is follow ed by a review on the stru ctu re, tissue d istrib u tio n and function of the Biliary G lycoprotein (BGP; CD6 6a) and ends w ith a description of the aims of this thesis.
1.2. Ad h e s io n m oleculef a m iu e s
M ost adhesion m olecules belong to one of the fo u r m ajor adhesion m olecule fam ilies. These com prise the integrins, the cadherins, the selectins and the im m unoglobulin super family. A dhesion interactions can be su b d iv id e d into four d ifferen t types: h o m o p h ilic w h en an adhesion molecule is able to bind to a sim ilar molecule on the surface of an o p posing cell; heterophilic adhesion w hen tw o different m olecules are able to bind to each other; hom otypic adhesion w hen cells of the sam e ty p e are able to form aggregates w hich m ay in v o lv e eith er h o m o p h ilic o r h e te ro p h ilic in te ra c tio n s b e tw e e n th e a d h e sio n m olecules m ed iatin g the agg reg atio n process; h etero ty p ic ad h esio n w hen different cell types are able to aggregate in a process th at could also be m ediated either by hom ophilic or heterophilic interactions.
1.2.1 The in teg rin family.
one type of p su b u n it |making functional classifications m ore appropriate.
The m olecular w eight of integrin p subunits ranges from 90 to 110 kD w ith th e exception of P4 w hich is 210kD. They all sh o w stro n g hom ology at th e am ino acid level (35-55%). T hey all co n tain 56 conserved cysteines (except P4 which has 48) arranged in four repeating units. C om parison of the extracellular regions of p l - p 7 revealed a hig h ly conserved region at the N -term inal half of the protein. The cytoplasmic dom ains are 40-50 residues long, except for p4 w hich has an u n u su a lly long (1018 am ino acids) cytoplasm ic tail containing four fibronectin type III repeats (H ogervost et al, 1990; Suzuki an d N aitoh, 1990). The P subunits also contain five cysteine rich repeats close to the transm em brane region. C onsensus tyrosine phosphorylation sequences ap p ear in the cytoplasm ic tails of p i, P3, p5, p6 an d p7. A lthough P2 (Hibbs et al, 1991) and p4 (Sacchi et al, 1989) lack such a sequence they are also p h o sp h o ry lated on se rin e /ty ro sin e residues. F u rth er stru ctu ral diversity of the P subunits is provided by the existence of variant forms w hich contain alternative cytoplasmic dom ains or lack four cysteine rich repeats.
The integrin a subunits have a m olecular w eight that ranges from 150 to 200 kD w ith an am ino acid sequence hom ology of 20-60%. All contain sev en hom ologous d om ains of a p p ro x im ately 60 resid u es. These contain either 4 or 3 divalent cation binding sites. Subunits a l , a2, aL , aM and aX all contain an extra 200 am ino acids inserted betw een repeats 2 an d 3 know n as the I dom ain. The o th er a su b u n its u n d e rg o p ro te o ly tic cleavage in to h eav y a n d lig h t frag m e n ts w h ich are d isu lp h id e linked except for the a 4 su b u n it. A d d itio n al stru ctu ral diversity w ithin the integrin a subunits can arise by alternative splicing. A schem atic representation of the dom ain organisation of the a and p integrin subunits is show n in Fig. 1.1.
in intercellular a d h esion by b in d in g to so m e of the m em b ers of the im m u n o g lo b u lin su p er fa m ily su ch as th e in tercellu la r a d h esio n m o lecu les ICAM -1, ICAM -2, ICAM -3 and the vascular cell ad h esion m olecule VC AM-1. Other ligands inclu de the proteins belon gin g to the desin tegrin fam ily. O riginally d iscovered in snake venom 's as p otent inhibitors of platelet aggregation m ediated by allb p 3 , cellular form s have since been id en tified (W eskam p and Blobel, 1994). M ost integrins are able to bind m ore than on e ligand (e.g.. a i p i binds lam inin or collagen and a 3 p l bin ds collagen , lam inin and fibronectin) and so m e exhibit alternative sp ecificities d ep en d in g on the cell typ e in w h ich th ey are expressed (Elices et al, 1990).
EF-hand-like dom ains
a I domain
Tm
I
IV V VI VIICysteine-rich
region Tm
Conserved
region
F i g .l .l Schem atic representation of the a and P integrin subunits dom ain organisation.
D om ains V-VII in I d om ain containing integrins (or IV-VII in others) h ave EF-hand-like
cation binding sequences. Tm, transmembrane dom ain.
m ore global conform ational changes in the ligand. H ow ever n o t all integrins bind this ligand sequence as it has been show n for fibrinogen (Farrell and T hiagarajan, 1994) an d the leukocyte lig an d s VC AM-1 (M akarem et al, 1994) and ICAM-1 (Randi an d H ogg, 1994). Binding is cation dependent and requires both subunits. It also requires activation of the integrin receptor by specific signals (e.g. T cell activation by antigen presentation or antibodies) resulting in a conform ational change w hich enables the integrins to bin d their ligands. They play a central role in m echanism s such as cell differentiation, leukocyte trafficking, platelet aggregation and T cell activation.
1.2.2 The C adherin family.
linking the successive extracellular dom ains, conferring a rod-like shape to the cadherin m olecules.
The prototypes of the cadherin fam ily are the w ell characterised E, N and P (p latelet) cadherins. The fa m ily has h o w e v e r gro w n to in clu d e m em bers w ith add ition al extracellular repeats (Sano et al, 1993), no cy to p la sm ic region (O kazaki et al, 1994) or m o d ifie d or d istin ctly different cytoplasm ic dom ains (Berndorff et al, 1994). These in clu de the T (truncated)-cadherin, m olecu les o f the d esm osom al, d esm ocollin and d esm o g lein subfam ily and the ret proto-on cogen e (Buxton et al, 1992; Takeichi, 1993).
P r o te a s e
C le a v a g e S ite
4
2+ 2+
Cà Ch Ca
a b a b a b
2+
TM
A m ino
1 P m T9$)Qn EC1 EC2 EC3 EC4
T erm in u s |■ Cys Cyt Tail
Cell binding
L
Carboxyl
T erm inus
J
C atenin
binding
Fig.1.2 Schem atic representation of the consensus structure of classical cadherins. Mature
cadherins are transm em brane (TM ) proteins derived from a precursor b y cleavage o f the
pre-region. The extracellular region is co m p o sed of four extracellular repeats (E C l-4 )
w ith calcium b in d in g m otifs (a and b ) and a c y ste in e con tain in g region (C y s). The
cytoplasm ic tail (Cyt tail) has a catenin bin d in g region.
in m aintaining the integrity of m ulti cellular structures. E-cadherin and N -cadherin particip ate in the form ation of the zonula adherens, their cytoplasm ic dom ains interacting w ith a , p, and y catenins th at link the cadherin molecules to the actin based cytoskeleton (Ozawa an d Kemler, 1992; Kemler, 1993). Gene pro d u cts of the src proto-oncogene fam ily have also been found expressed at the zonula adherens (Takeichi et al, 1991) and m ay be responsible for cadherin phosphorylation raising the possibility th at cadherin m ed iated cell junctions m ig h t be u sed for in tercellu lar signalling (M atsuyoshi et al, 1992; Saffel et al, 1992). C adherins have also been show n to be expressed d u rin g early m uscle developm ent and are dow n regulated b u t n o t absent in m atu re m uscle (Moore an d W alsh, 1993; C ifuentes-D iaz et al, 1994) w ith N -cadherin participating in m yoblast fusion an d being d o w n reg u lated by nerve activity after the form ation of the n eu ro m u scu lar contacts (H ahn and Covault, 1992; Fredette et al, 1993). M any cadherins are also present in neural tissues (Redies et al, 1993). It has been suggested th at they m ay participate in the segregation of sensory neurons into functional groups (Fredette et al, 1994; Shim am ura et al, 1992). H o w ev er w ith the exception of N -cadherin, w hich p ro m o tes n e u rite g ro w th (Rathjen, 1991), little is know n about the function of o th er cad h erin s in the nervous system.
1.2.3 The selectin family.
transm em brane d om ain is fo llo w e d by a short cy to p la sm ic tail. A sch em a tic rep resen ta tio n of the sele c tin s d o m a in o r g a n isa tio n is d epicted in Fig. 1.3. The high d egree of h o m o lo g y fou nd b e tw een the selectins strongly suggests that they w ere produced by duplication of an ancestral gene, follow ed by exon diversification and duplication. This is supported by the fact that the genes for all three proteins are clustered over a short region o f hum an and m o u se chrom osom e 1 (rev iew ed by
Vestweber, 1992; Lasky, 1992).
L-Se lectin P-Selectin E-Selectin
KEY:
V - C- type lectin domain - EGF domain
0 - SCR - short c o n se n su s repeats
Fig.1.3 Schematic representation of the dom ain organisation of the selectin m olecu les.
biological relevance, the additional b inding activity of the selectins to various su lp h ated carbohydrates suggests the possibility of alternative carbohydrate ligands (Green et al, 1992; N elson et al, 1993). So far, the m ost potent naturally occurring carbohydrate ligands for both L- and E- selectin are Le^ and Le^ derivatives, in w hich the hydroxyl gro u p on carbon 3 of galactose is su lp h a te d ra th e r th en sialy lated . W hen im m obilised in the form of glycolipids, 3'-sulphated Le^ an d Le^ w ere show n to su p p o rt direct binding of all 3 selectins at least as strongly as sLe%/& (Green et al, 1992; Yuen et al, 1992; B randley et al, 1993). A diag ram of the sLex m olecule is show n in Fig. 1.4. A lth o u g h these oligosaccharides are often p a rt of the glycosylation patterns of proteins, only a few biological ligands for selectins have been identified. These are GlyCAM-1, a lectin-like receptor glycoprotein, (Imai et al, 1991; Lasky et al, 1992), CD34 (Imai et al, 1991; Baum hueter et al, 1994), the P-selectin glycoprotein ligand 1 (PSGL-1) (Sako et al, 1993; N orgard et al, 1993) and MAdCAM -1, a m ucosal vascular adressin (Berg et al, 1993). Ail have extracellular dom ains w ith a m ucin organisation, i.e. s e r in e /threonine rich regions th at are densely su b stitu ted w ith O -linked carbohydrate chains (Shimizu and Shaw, 1993). A num ber of other potential ligands, including BGP, have been described (Aruffo et al, 1991; N o rg ard et al, 1993; Stocks et al, 1993; W alcheck et al, 1993; Lenter et al, 1994;), b u t for these fu rth er studies are necessary to establish w h eth er they serve as biological ligands for the selectins or if they are m erely cross-reactive as a result of their fortuitous expression of carbohydrate motifs. The three- dim ensional structure of the lectin and EGF dom ains of E-selectin has been resolved (Graves et al, 1994), b u t so far a co-crystal w ith a b o u n d oligosaccharide is not yet available.
G alactose
ring G lucosam ine
ring OH
OH HO,
NHCOCH OH
OH HoC
OH Sialic
acid ring
F u cose ring
Fig.1.4 Schem atic representation of the sLex carbohydrate m oiety.
The e n o r m o u s p o ten tia l o f ca rb o h y d ra tes for e n c o d in g sp e c ific inform ation w hich cou ld be u sed in cell recogn ition had lo n g been postulated. The discovery of the selectin fam ily has played a major role in v a lid a tin g th e v ie w that c a rb o h y d r a te s d e fin e r e c o g n itio n d eterm in a n ts in s p e c ific c e ll-c e ll a d h e sio n e v e n ts (V arki, 1994). A lth o u g h co n sid era b le p rogress has b een m ad e in id e n tify in g the m inim al structures for ligan d recognition, m uch rem ains to be learnt about the context in w hich they are presented to confer sp ecificity and h o w selectin m ediated interactions are interlinked w ith other adh esion pathways.
1.2.4 The im m unoglobulin super family.
an d can be viral an d grow th factor receptors. They also function as intercellular adhesion molecules an d signal transducing receptors or as both (reviewed by Buck, 1992).
S tru ctu rally the im m u n o g lo b u lin hom ology u n it consists of 70-110 am ino acids organised into 7-9 P strands. These contain 5-10 am ino a d d s and are juxtaposed in an anti-parallel m anner such th at the hydrophobic side chains are facing one another on the interior of the m olecule w ith the hydrophilic side chains exposed to the external side. The p strands form a sandw ich stabilised by the interior hydrophobic groups an d in m ost cases by a disulphide bridge form ed betw een specific p strands. A m odel of the im m unoglobulin fold is show n in Fig.1.5. The hom ology units have been classified as V or C according to their sim ilarity to the hom ology units found on the variable (V) or constant (C) dom ains of the im m unoglobulin molecule. In the V dom ain, the p sandw ich is fo rm ed by tw o P sheets, one of 3 an d the o th er of 4 P s tra n d s. Sequestered betw een these two P sheets are an extra tw o p strands, C and C", w hich in antibodies form the second hypervariable loop. In the C dom ain these two strands are m issing. The C dom ain has further been subclassified into C l and C2 type. The C2 type dom ain is m ainly found in the IgSF adhesion m olecules an d is generally m ore com pact th an eith er the V or C l dom ains. The genetic o rg an isatio n of the IgSF m olecules suggests a sophisticated m echanism for the ev o lu tio n ary co n serv atio n of stru c tu ra l a n d fu n ctio n al d iv e rsity (W illiam s an d Barclay, 1988). Each Ig hom ology u n it is encoded by a single exon, thus p reserv in g each dom ain as a functionally im p o rtan t elem ent of any receptor. The exon organisation is such th at the 3' end of an exon ends w ith the first base of the first codon in the im m ediately follow ing hom ology u n it and the 5' end of each exon begins w ithin the last tw o bases of the codon of the im m ed iately p rec ed in g h om ology un it. Exceptions to this are all the Ig-like dom ains of n eu ral cell adhesion molecule (NCAM), and dom ain 1 of CD4, w here each dom ain is encoded by two exons. This m ay reflect the possibility th at the im m unoglobulin fold w as itself form ed by the d u p lic atio n of an an cestral d o m ain (Bourgois, 1975).
with interactions betw een non-identical family members, such as CD2 and LFA-3, being an extension of this m echanism . The three dimensional structure of the first two domains of CD2 has been resolved by X-ray crystallography (Jones et al, 1992) and the presence of crystals involved in hom ophilic binding in the solution further confirms both the adhesion capacity of the im m unoglobulin fold and the fact that hom ophilic b inding m ay have been the first type of interaction developed by the IgSF.
C - T e r m i n u s T e r m i n u s
F ig .1 .5 Structure of an im m u n o g lo b u lin light chain variable (Vl) and con stan t (C L )d o m a in s (from the X-ray crystallograp h ic stu d ie s for a B ence-Jones protein
(Schiffler et al, 1973). O ne face of each dom ain is com p osed of four chains (grey arrows)
arranged in an anti-parallel b-pleated structure stabilized by interchain hydrogen bonds
betw een the CO and N H groups running along the p ep tid e backbone, and the 3 chains
(green arrows) in the other face of the dom ain; the dark bar represents the intra-chain
d isu lp h id e bond; the stripped loop s represent the hypervariable regions w hich are the
Ligands for the IgSF now com prise non-IgSF m em bers such as LFA-1 (C D 18/C D lla) and Mac-1 (C D 18/C D llb) which bind ICAM-1 (Marlin and Springer, 1987; D iam ond et al, 1990) VLA-4 ( a 4 /|3 l) an d a 4 / p 7 w hich bin d to the vascular adhesion m olecule 1 (VCAM-1) (Elices et al, 1990; Ruegg et al, 1992). All these belong to the integrin family. Com ponents of the extracellular m atrix are also am ong the counter receptors for the IgSF. For exam ple collagen and h ep arin are recognised by the m yelin associated glycoprotein (MAG) and h ep arin an d h e p aran su lp h ate are recognised by NCAM (Cole and Glaser, 1986; Reyes et al, 1990) and the platelet endothelial cell adhesion m olecule, PECAM-1 (N ew m an et al, 1990; A lbelda et al, 1991). It has been proposed th at proteoglycans could m o d u la te th e h o m o p h ilic m e d ia te d a d h e sio n of IgSF m em b ers. PECAM-1 tran sfected L cells have been sh o w n to u n d e rg o C a^ + d ependent PECAM-1 m ediated aggregation (Albelda et al, 1991). This co u ld h o w e v er be in h ib ite d by h e p a rin , h e p a ra n s u lp h a te an d chondroitan sulphate or sim ply by proteoglycanase digestion of the cells (Delisser et al, 1993). Also the observation th at HIV binding to CD4 can be blocked by dextran su lp h ate and h ep arin (L ederm an et al, 1991) fu rth er points to a role in the m odulation of the function of the IgSF molecules by proteoglycans.
Buck, 1990; Springer, 1990; Butcher, 1991). For example, their role in the developm ent of the nervous system is particularly evident (D oherty et al, 1991; E delm an, 1992). A g ro u p of m olecules w ith m u ltip le Ig dom ains coupled to a varying num ber of fibronectin type III repeats, is involved in cell m igration, the stim u latio n an d inhibition of n eu rite o u tg ro w th an d th e adhesion of n eu rites (Rathjen an d Jessel, 1991). H om ologues of this group have been fo u n d in species as diverse as insects an d m an em phasising the fu n d am en tal im p o rtan ce of these molecules. C ellular interactions w ithin the im m une system are no less complex than those found w ithin the nervous system and here too IgSF m em bers are w idely used. Exam ples are the interactions of resting T cells w ith antigen presenting cells (involving ICAM-1 binding to LFA-1) (Springer, 1990); adhesion of neutrophils and m acrophages to activated endothelial cells and subsequent transm igration th ro u g h post capillary venules (Springer, 1990; W illiam s, 1991); m acrophage m ovem ent into the skin and peritoneum , and for the hom ing of lym phocytes to specific lym ph nodes (Albelda and Buck, 1990; Butcher, 1991; M akgoba et al, 1989; Shim izu et al, 1991).
The stru c tu ra l featu res of the IgSF m olecules h av e allo w ed th e acquisition of a w ide range of functional d iv ersity resu ltin g in the evolution of im p o rtan t m olecules for m o rp h o reg u lato ry processes as well as the evolution of a highly sophisticated im m une system in w hich they function as p art of a well coordinated series of events involving both cell-cell and cell-matrix adhesion.
1.3 THE CARCINOEMBRYONIC ANTIGEN (CEA> GENE FAMILY
1.3.1 Historical perspective.
before its com plicated m olecular n atu re w as defined. Later, CEA w as show n to be present at low concentrations in body fluids an d colonic m ucosa of healthy individuals (Chu et al, 1972; Egan et al, 1977; Fritsche and Mach, 1977). It also became apparent that CEA was not an entity, b u t rath e r a m em ber of a heterogeneous fam ily of related cross-reactive glycoproteins th at could n o t be clearly d istin g u ish ed by the technical m eans available at the time. In 1972 the first of these glycoproteins to be described (von Kleist et al, 1972) received the uncom itting nam e of non specific cro ss-reactiv e an tig en o r NCA . Successful clo n in g a n d ch aracterisatio n of the N C A s cDN As sh o w ed th a t th e C EA -gene rep re se n ts th e p ro to ty p e of a larg e gene fam ily (T hom pson an d Z im m erm ann, 1988) and in 1989 the CEA gene-fam ily w as officially constituted at an International Society for O ncodevelopm ental Biology and M edicine, ISOBM, W orkshop (Barnett an d Z im m erm ann, 1990).
1.3.2 The CEA fam ily subgroups.
A ccording to sequence hom ology, the CEA family w as initially divided into tw o subgroups, the m em brane b o u n d or CEA su b g ro u p and the p regnancy specific protein (PSG) or secreted subgroup. W ithin each su b g ro u p the different m em bers possess a hig h level of sequence hom ology (80-95% hom ology at the nucleotide level), w hereas betw een s u b g ro u p s th is h o m o lo g y is a ro u n d 65-75% (T h o m p so n a n d Z im m erm ann, 1988).
gene), CD6 6c (NCA50/90), CD6 6d (CGMl) and CD6 6e (CEA) (Schlossman et al, 1994).
The secreted, pregnancy specific glycoproteins (PSG) subgroup contains at least 11 different m em bers (PSGl to PSG8 an d P S G ll to PSG13) w hich w ere independently described by Tatarinov and M asiukevich (Tatarinov an d M asiukevich, 1970) and Bohn (Bohn, 1971) w ith PSG9 an d PSGIO latter fo u n d to be allelic v arian ts of PSG4 an d PSG6 resp ectiv ely (Teglund et al, 1994). Several of these are differentially spliced leading to the production of m any protein isoforms (Streydio et al, 1990). The PSGs are m ainly p ro d u c e d by the p lacen tal sy n cy tio tro p h o b lasts d u rin g pregnancy (Tatarinov and M asiukevich, 1970; Bohn, 1971; Lei et al, 1992) and can be detected after 2-3 weeks of gestation (Lin et al, 1974). They have also been found in a num ber of nonplacental tissues, nam ely in testis (Borjigin et al, 1990), salivary glands (Zoubir et al, 1990), fetal liver (Zim m erm an et al, 1989), Hela cells (Chan et al, 1988) and m yeloid cell lines (Oikawa et al, 1989a; Barnett et al, 1990). H ow ever, the im portance of this extraplacental expression is not clear. In m alignancy they are found m ainly in trophoblastic tum ours, b u t have also been fo u n d in b rea st an d colonic carcinom as, in m u ltip le m yelom as an d in the leukaem ia cell line HL-60, as well as in the sera of patients w ith various gynaecological carcinomas (Oikawa et al, 1989a). PSG serum levels have also been used to m onitor patients w ith choriocarcinom a (Tatarinov and Sokolov, 1977) and certain PSGs, especially PSG6, seem to be associated w ith hydatidiform m ole (Leslie et al, 1990). F urther indication of the im portance of PSGs in pregnancy is the fact th at low serum levels of these proteins correlate w ith threatened abortion (Bischof, 1984) raising the possibility th at PSGs m ay have a role in protecting the developing foetus from the m aternal im m une system.
More recently a th ird new subgroup of the CEA gene fam ily has been identified (Teglund et al, 1994). It com prises 6 m em bers, C G M l3 to C G M l8, which show a 94-99% nucleotide sequence id en tity w ith each other, w hile the degree of hom ology w ith either the PSG or the CEA subgroup is betw een 70-75%. W hether these new genes are functional is not know n at the m oment.
6 in the n ew su b g ro u p . They are lo cated on th e lo n g arm of chrom osom e 19 at positions 19ql3.1-2 (Zim m erm an et al, 1988; Olsen et al, 1994) close to the genes encoding for the Transform ing G row th Factor (TG F-fil) a n d cy to ch ro m e P450 fam ily IIA (Schonk e t al, 1990). C onsidering the extensive analysis of the CEA gene fam ily region (Olsen et al, 1994; Teglund et al, 1994) it seems likely that m ost if n o t all of the CEA fam ily genes have now been identified. A list of these genes is show n in Table 1.1 and a schematic representation of the chrom osom al distribution of the CEA gene-family is show n in Fig.1.6.
_ < 2
8 8 8 8 S '
8
o | | | i 88088 88080 80880 800
C û O O O ü Q.Ü.ÜÛ.CL CLCLÜQ. Ü O . Ü Û . C L Ü Û . Ü Ü/ -
y -
150 kbV
CEN TEL
F ig.1.6 Schem atic representation of CEA g en e fam ily organization in the ch rom osom e
19ql3.2 region. The location o f the 29 individual g en e fam ily m em ber is indicated. All
gen e s are d ep icted in an arbitrarily un iform size. The sh a d in g d istin g u ish e s g en es
d etected w ith N -d o m a in probes on ly (w h ite b o x es), c on stan t d o m a in p rob es o n ly
(stippled boxes), or both N -d om ain and constant d om ain probes (black boxes). CEN
d en o tes centrom ere and TEL denotes telom ere. W here d eterm in ed , the d irection of
transcription is indicated by the arrows beneath the b oxes, (adapted from O lsen et al,
1994).
v a ry in g len g th d e p e n d in g on th e d ifferen t splice v a ria n ts b ein g transcribed (H inoda et al, 1988). Fig.1.7 show s the dom ain organisation of the m ain CEA subgroup members. All m em bers of the PSG subgroup and the third subgroup lack the transm em brane dom ain w hich probably has functional consequences in respect to the secretion, m ost PSGs being easily secreted (Zheng et al, 1990).
The com plexity of the CEA fam ily is fu rth er increased by differential splicing and post-translational m odification (e.g. glycosylation) of som e of its m em bers (Thom pson and Z im m erm ann, 1988).
TABLE 1.1 - HUMAN CEA GENE FAMILY MEMBERS.
CEA SUBGROUP / CD PSG SUBGROUP THIRD SUBGROUP
CEA CD6 6e PSGl CGM13
NCA50/90 CD6 6C PSG2 CGM14
CGM6 or NCA95 CD66b PSG3 CGM15
BGP CD6 6a PSG4 CGM16
CGMl CD6 6d PSG5 CGM17
CGM2 PSG6 CGM18
CGM7 PSG7
CGM8 pg PSG8
CGM9 P S G ll
CGMIO pg PSG12
C G M ll pg PSG13
CGM12 pg
CEA SUBGROUP
CEA
NCA-95
BGPa
NCA-50/90
CGM 1a
CGM 1c
CGM7
L N
m
a s
L N
N
L N
M
l
A1 B1 A2 B2 A3 B3
C C 0 C C C C C C C C 0 GPI
A1 B1
C C C C GPI
A1 B1 A2 TTVI Cyt
C C C CC C C
A1 B1
0 C 0 C
L N TTy/1 C y t
TTR
L N m Cyt
M
i i i
L N TVI Cyt
m m s m
A1
C O ■ GPI
F ig .1 .7 Schem atic representation of the d om ain org a n isa tio n of the CEA su b grou p
m em bers. N , IgV -like N -term inal d om ain, A and B, IgC2-like dom ain s typ e A or B; c
in sid e the b o x rep resen ts the nu m b er o f c y ste in r e sid u e s in that d o m a in ; T M
tran sm em b ran e d o m a in ; C y t, cy to p la sm ic d o m a in (the d ifferen t p atterns on the
cytoplasm ic dom ain represent different alternative sp licin g even ts for this dom ain); GPI,
1.3.3 The rodent CEA gene family.
Probes from the hum an CEA gene fam ily m em bers w ere used to isolate an d characterise ro d en t CEA related m em bers (Beauchemin et al, 1989; Kodelja et al, 1989; Lin and Guidotti, 1989; Rebstock et al, 1990; Turbide et al, 1991). Based on sequence com parisons these w ere d iv id ed into 3 subgroups.
W ithin a subgroup, the genes reveal 80-90% sequence sim ilarity at the nucleotide level, whereas betw een subgroups it drops to 55-60%. To date 11 ro d en t CEA -related genes have been described (although som e of these m ay represent splice variants of the sam e gene) and are listed in Table 1.2. Rat C-CAM has al least two splice variants, one producing a 71 (C-CAMl) and the other a 10 (C-CAM2) am ino acid cytoplasmic tail.
Individual counterparts betw een the ro d en t and the h u m an CEA gene fam ily m em bers w ere im possible to determ ine based only on sequence com parisons. This is d u e to the fact th at a 50-60% sequence sim ilarity exists betw een the two families irrespective of the subgroups (review ed by Thom pson et al, 1991) and assignm ent of counterparts h ad to be done based on dom ain organisation, tissue expression and function.
1990). These expression patterns are as expected for counterparts of the hum an CEA subgroup. Both BgpB and rat C-CAM have been show n to m ed iate ad h esio n (T urbide et al, 1991; O cklind an d O brink, 1982; T ingstrom et al, 1990). F u rth er m ore th e in tracellu lar cytoplasm ic dom ain of C-CAM l has been show n to be involved in the tran sp o rt of bile salts through the canicular dom ain of hepatocytes (Sippel et al, 1993), in p h o sp h o ry latio n events activated by the in su lin receptor tyrosine kinase (Najjar et al, 1993) and in binding of calm odulin (Blikstad et al, 1992).
R odent subgroup 2 m em bers com pletely lack cytoplasm ic dom ains an d are directly secreted from the cells. T ranscripts for rnC G M l have been found in rat placenta (Rebstock et al, 1990; Beauchemin et al, 1989) and equally, transcripts for mmCGM5 have been found in m ouse placenta (review ed by T hom pson et al, 1991) in d icatin g th at this su b g ro u p encodes the rodent PSG counterparts.
The rodent subgroup 3 genes have only been partially characterised at the sequence level and little is know n about their expression patterns or function. Based on th eir high sequence sim ilarity , rnCG M 2 an d mmCGM8 are thought to be hom ologous genes.
TABLE 1.2 THE RODENT CEA-RELATED GENE FAMILY
SUBGROUP 1 SUBGROUP2 SUBGROUP 3
Mouse / Isoforms Mouse Mouse
Bgp BgpA and E m mCGM S
BgpB and C m m C G M6 m m C G M8
BgpD and F m m C G M 7
BgpH m m CG M 9
Rat / Isoforms Rat Rat
C-CAM C-CAMl rnC G M l
C-CAM2 rnCGM3
rnCGM4
1.3.4 Tissue and cell d istrib u tio n of the CEA subgroup proteins.
U ntil recently m ost antibodies available for detecting proteins belonging to the CEA subgroup w ould cross-react w ith the different m em bers, d u e to their great am ino acid sequence sim ilarity, m aking differential tissue expression studies difficult to perform . H ow ever using N o rth ern blot analysis. Polym erase Chain Reaction (PCR), in situ hybridisation as well as im m unocytostaining, it has been possible to discrim inate betw een the different transcripts of the CEA fam ily. D espite their g reat sequence sim ilarity, the different m em bers of the CEA su b g ro u p reveal very different expression patterns. It appears th at CEA and N C A -50/90 are coexpressed in a num ber of colorectal adenom as or derived cell lines as well as in norm al colonic m ucosa (H igashide et al, 1990). They are also detected in the m icrovilli of sw eat glands, in m ultivesicular bodies in sw eat ducts and in syringom as and porocarcinom as (Metze et al, 1996). N C A -50/90 is also expressed in cutaneous tum ours (M etze et al, 1992) and in lung and breast cancer (C ournoyer et al, 1988). CEA has been reported to be present in oesophagus, stom ach and pancreas, tum ours of gastrointestinal origin as well as in lung, ovary and b lad d er carcinom as (Pavelic et ai, 1991).
up-regulated d u rin g neutrophilic developm ent b u t is sw itched off d u rin g m onocytic differentiation.
BGP was first discovered in the epithelium of bile canaliculi (Svenberg et al, 1976), b u t is also expressed in norm al granulocytes (Berling et al, 1990), norm al colon (Barnett et al, 1989), fetal liver (H inoda et al, 1990), in vesicular structures of higher differentiated sebocytes (M etze et al, 1996) and in squam ous cell carcinom as of the lu n g (C ournoyer et al, 1988). M ore recently the availability of a specific BGP m onoclonal antibody m ade possible a m ore thorough stu d y of BGP localisation in hu m an tissues (Prall et al, 1996). BGP w as found in epithelium and e n d o th e liu m of a w id e ran g e of n o rm al h u m a n tissu es. A pical m em branous staining of epithelia w as found in oesophageal glands, in the d u o d en u m B runner's glands, in the superficial absorptive cells of the colon, in large and sm all ducts of the pancreas, in bile ducts, in gallbladder, in prostate, in the cervix, in uterine glands an d in breasts ducts. Endothelial staining was found in the capilliaries in islets of the pancreas, in glom erular capillaries and vasa recta of the kidney, in small arteries and venules of the prostate, in sm all en d o m etrial vessels, in small arteries, capillaries and venules of the placenta and in sinusoids of the adrenal gland. Staining was also found in enterocytes of the small intestine, in goblet cells of the colon an d in hepatocytes along bile canaliculi. Bone m arrow staining of granulocytes an d their precursors, including myelocytes confirm ed the presence of BGP in these cells (Watt et al, 1991 and 1994). O ther authors have also reported the presence of BGP on T cells and on a sub-population (CD16"CD56'^) of natural killer (NK) cells (Moller et al, 1996). Both BGP an d N C A 50/90 have also been found on B cell lym phom as (Khan et al, 1993) and on bone m arrow and peripheral blood cells of acute lym phoblastic leukaem ia (ALL) patients (H anenberg et al, 1994) w ith either T or B cell origin. BGP as also been found in cell lines of B-cell origin (Khan et al, 1993; H anenberg et al, 1994).
1.3.5 The CEA related m olecules as adhesion molecules.
The fact that the constant-like dom ains of the CEA family m em bers w ere of the C2 set, w hich includes a n u m b er of cell adhesion m olecules, n am ely NCAM (C unningham et al, 1987), CD2 (Sewell et al, 1986), in tercellu lar ad h esio n m olecules I a n d II (ICAM-1 a n d ICAM-2) (Staunton et al, 1988 and 1989) and the m yelin-associated glycoprotein (MAG) (Salzer et al, 1987), led to the experim ents aim ed at testing the adhesion properties of the CEA subgroup m em bers.
So far h om otypic ad h esio n has been estab lish ed in vitro for CEA (Benchimol et al, 1989), NCA -50/90 (Oikawa et al, 1989b) and BGP (Rojas et al, 1990; Teixeira et al, 1994; W att et al, 1994) w ith each one of these interacting also w ith one another in a heterophilic m anner (Oikaw a et al, 1992). CEA and NCA hom ophilic adhesion is tem perature and cation in d ep en d en t (review ed by T hom pson an d Z im m erm ann, 1988). BGP hom ophilic adhesion was reported to be both calcium and tem perature d ep en d en t (Rojas et al, 1990; Teixeira et al, 1994), how ever its cation d e p e n d e n c y is still co n tro v e rsial since re p o rts of calcium an d tem perature independency are also available (Oikawa et al, 1992).
Specific binding of CEA, BGP and N CA -50/90 to certain strains of E. coli has also been reported (Leusch et al, 1990 an d 1991). Binding of CEA p resen t in the norm al colonic m ucosa to bacteria could function as a m echanism to facilitate bacterial colonisation of the colon w hile the shedding of CEA into the lum en could regulate the steady state level of bacteria in the colon. In addition, NCA present in the granulocytes has been show n to bind invading bacteria thus facilitating their phagocytosis
(Leusch et al, 1991).
A dhesion to collagen type I and activated endothelium have also been reported for CEA and BGP respectively (Pignatelli et al, 1990; Kuijpers et al, 1992). In m etastasising tum our cells and in m igrating em bryonic cells th is m ay be im p o rta n t in m e d ia tin g in te ra c tio n s w ith b a sem e n t m em branes. In granulocytes this could be im p o rtan t in the process th at m ediates the adhesion of these cells to endothelium in o rd er to allow them to penetrate tissues and reach the sites of inflam m ation.
d ifferen t im m u n o th erap y ap p ro ach es an d th eir sid e effects can be analysed (Eades-Perner and Zim m erm ann, 1995).
1.4 THE BILIARY GLYCOPROTEIN (BGP) MOLECLTLES
1.4.1 The BGP isoforms.
A t least eight transm em brane BGP m olecules, (generated by differential splicing of a single BGP gene), have been identified by cloning and PCR analyses (Barnett et al, 1989 and 1993; W att et al, 1994). The existence of three soluble forms has also been suggested (Kuroki et al, 1991), although this has recently been d isp u ted (Barnett et al, 1993) and their protein products have not as yet been identified. Studies have show n th at the several m ature BGP molecules can occur by alternative splicing (Barnett et al, 1989 and 1993). The largest BGP molecule, BGPa, consists of a 108 am ino acid N -term inal dom ain, tw o 178 am ino acid IgC-like dom ains (A l and Bl follow ing the nom enclature agreed in the XVIIth M eeting of the International Society for O ncodevelopm ental Biology an d M edicine [Barnett et al, 1990]), a 100 am ino acid A2 dom ain, follow ed by a 32 am ino acid transm em brane region and a 71 am ino acid cytoplasm ic tail (Barnett et al, 1989). The several isoforms obtained by alternative spicing show the following dom ain structure: BGPb lacks the th ird A2 dom ain while BGPc keeps the A2 dom ain b u t has only a short cytoplasmic tail of 9 amino acids generated by a 53 nucleotide deletion of exon cytl. As 53bp is not a m ultiple of a codon triplet, nucleotides after the deletion site yield a new read in g fram e th at ru n s in to a sto p codon after the tran slatio n of six new am ino acids. This generates a 9 am ino acid cytoplasmic tail instead of the 71 am ino acid tail th at is generated w hen both cy tl and cyt2 exons are translated. BGPd lacks both the 3rd A2 dom ain an d the long cytoplasm ic tail (Fig.1.8). The BGPb an d BGPd isoform s have a fifth cysteine after their A lB l dom ains th at does n o t seem to be involved in the form ation of an intrachain disu lp h id e bridge b u t m ay be involved in dim erization (Barnett et al, 1989).
A t the am ino acid level, the N -term inal an d the A lB l BGP dom ain sequences exhibit m ore than 70% id en tity w ith o th er m em bers of the CEA subfamily, (including CEA itself and the non-specific cross reacting antigen, N CA -50/90), w hereas the A2 dom ain show s only 45% identity (H inoda et al, 1988; Barnett et al, 1989). In contrast, the Y2 and Z2 dom ains are specific for the BGP family, displaying significant hom ology at the nucleotide level w ith an in v erted an d tru n ca te d h u m an A lu
repeat sequence (Barnett et al, 1993). A schem atic dom ain representation of the different BGP splice variants is show n in Fig.1.8. All isoforms are heavily N -glycosylated, approxim ately 40% by w eight (B arnett et al, 1989). T he la rg e r c y to p lasm ic tail sh o w s se v e ra l p o te n tia l p h o s p h o ry la tio n sites w hich w e re sh o w n to be c o n stitu tiv e ly phosphorylated on serine, threonine and tyrosine (Afar et al, 1992). A close association betw een the BGPs an d m em brane associated protein tyrosine kinases has also been rep o rted (Skubitz et al, 1995). These fin d in g s su g g est th a t BGPs m ay be lin k ed to signal tran sd u ctio n pathw ays regulated by the alternative splicing of the cytoplasmic tail. 1.4.2 Possible functional roles of BGP.
Like all the other members of the CEA family the role of BGP in vivo is still u n d er investigation. In vitro BGP has been show n to be involved in hom ophilic and heterophilic adhesion (Rojas et al, 1990; O ikaw a et al, 1992). The latter m ay involve interactions w ith other m em bers of the CEA subgroup (Oikawa et al, 1992), w ith integrins (Stocks et al 1995) or w ith bacteria and endothelium (Leusch et al, 1991; Stocks et al, 1990). The presence of the CD15 epitope or sialyl Lewis X (sLe^ ) on BGPa (Stocks et al, 1990; Stocks and Kerr, 1993) suggests that adhesion of BGP to E-selectins on activated endothelium m ay be m ed iated by the CD15 epitope an d p articip ate in the firsts steps of n e u tro p h il adhesion to e n d o th e liu m .
BILIARY GLYCOPROTEIN ISOFORMS
BGPa
BGPb
BGPc
BGPd
BGPx
BGPx'
BGPy
BGPy’
BGPz
BGPz'
L N A l B l A 2 TM C y tl C yt2
I
m e m :
L N A1 B1 TM Cyt1 C yt2
L N A l B l TM Cyt2
L N TM C ytl C yt2
m - \ / / / / / / .
L N TM C yl2
L N A l B l Y TM C y tl Cyt2
L N A l B l Y TM C yt2
L N A l B l Z TM C y tl Cyt2
L N A l B l Z TM C yt2
Fig.1.8 Schem atic rep resen tation o f the d iffere n t BGP sp lic e varian ts. L, lea d er
sequence; N , IgV-like N -term inal dom ain; A and B, IgC2-like dom ain s typ e A or B; Y and
Z ex tra c ellu la r d o m a in s w ith a se q u e n c e sim ila r to an A lu -ty p e rep eat; T M
transmembrane dom ain; C y tl and 2, nucleotide sequences from tw o different exons coding
hu m an BGP m ight be involved in the tran sp o rt of bile acids as it w as show n for the rat hom ologue (Sippel et al, 1993). In m ice, the BGP h om ologues have also been fo u n d to acts as an h ep atitis recep to r (Dveksler et al, 1993).
As discussed previously BGP can also bin d som e strains of E. coli and Salm onella (Leusch et al, 1990). This b in d in g seem s to be m annose- d ep en d en t and a lectin/ carbohydrate interaction and m ay facilitate the phagocytosis of bacteria by the granulocytes expressing the BGP.
The phosphorylation of the cytoplasm ic tail of the BGP m olecules also suggest th at these m olecules m ay be involved in signal tran sd u ctio n p athw ays as has been show n for other m em bers of the IgSF m olecules (Veillette et al, 1988; Samelson et al, 1990). For exam ple MAG, another m em ber of the Ig super family, has significant hom ology to BGP in their respective cytoplasm ic dom ains. MAG also has altern ativ e splicing v arian ts th at generate isoform s w ith differen t cytoplasm ic dom ains (Frail and Braun, 1984; Salzer et al, 1987). In both cases the isoforms w ith the sh o rt cytoplasm ic d om ains show lo w or u n d e te ctab le in vivo
phosphorylation w hile the isoforms w ith the long cytoplasm ic tails are extensively phosphorylated on serine and tyrosine residues (Afar et al, 1990 an d 1992). The p h o sp h o ry latio n sites are conserved in these adhesion molecules suggesting that phosphorylation/ d ep h o sp h o ry latio n events m ay be im portant in regulating their biochemical and biological properties nam ely in the m odulation of their adhesive properties. More recently, BGP has been show n to associate w ith tyrosine kinases of the src fam ily, nam ely Lyn an d H ck (Skubitz et al, 1995) in activated granulocytes and w ith pp60-src in HT-29 cell lines a n d granulocytes (Brummer et al, 1995).
1.4.3 BGP in m alignancy.





![Fig. 3.13 BGPc-Fc [N, NAIBI and NAIBIA2] soluble domain fusion proteins and CD33-Fc](https://thumb-us.123doks.com/thumbv2/123dok_us/9000733.1437795/104.596.89.546.75.524/fig-bgpc-naibi-naibia-soluble-domain-fusion-proteins.webp)
