- S1 -
Supporting Information for
Development of engineered ferredoxin reductase systems for the efficient
hydroxylation of steroidal substrates
Zhangliang Zhu†, Xin Gao†, Zhan Song†, Chao Li†, Fuping Lu†*, Masaru Tanokura†, ‡* and
Hui-Min Qin†*
† Key Laboratory of Industrial Fermentation Microbiology of the Ministry of Education; Tianjin
Key Laboratory of Industrial Microbiology; College of Biotechnology, Tianjin University of Science and Technology; National Engineering Laboratory for Industrial Enzymes; Tianjin 300457, P. R. China
‡ Laboratory of Basic Science on Healthy Longevity, Department of Applied Biological
Chemistry, Graduate School of Agricultural and Life Sciences, The University of Tokyo, 1-1-1 Yayoi, Bunkyo, Tokyo 113-8657, Japan
*Corresponding authors: College of Biotechnology, Tianjin University of Science and Technology, F. Lu: [email protected]; M. Tanokura: [email protected]; H.-M. Qin: [email protected].
Tel: +86-22-60602949. Fax: +86-22-60602298
Number of pages: 20 Number of tables: 2 Number of figures: 8
- S2 -
Table of contents
1.
Bacterial strains, plasmids, and chemicals
2.
Plasmids construction, and protein expression and purification
3.
Construction of modified KshB and TDO
4.
Construction of whole-cell catalysts for the transformation of AD
5.
Bioconversion of AD to produce 9OHAD
6.
Structural modeling of KshB
7.
Differential scanning fluorimetry
8.
Supplementary tables and figures
- S3 - Bacterial strains, plasmids, and chemicals
All strains and vectors used in this study are listed in Table S1. Escherichia coli JM109 was used for cloning, and E. coli BL21 (DE3) was used for protein expression. pET28a, pCold I, and pETDuet-1 were used for the expression of target proteins. 4-Androstene-3,17-dione (AD) and 9α-hydroxy-4-androstene-3,17-dione (9OHAD) were purchased from Shanghai Macklin Biochemical Co. Ltd. (China). DNA polymerase (Q5 High Fidelity Polymerase), restriction enzymes, T4 DNA ligase, and an NEBuilder high-fidelity DNA assembly cloning kit were purchased from New England BioLabs (Ipswich, MA, USA). Oligonucleotides and genes were synthesized by Genewiz (Suzhou, China). Other reagents and chemicals used in this study were of the highest grade available.
Plasmids construction, and protein expression and purification
Gene sequences encoding KshA (Protein ID: WP_003419211.1), FDH (Protein ID: OWB80117.1), DMR1 (Protein ID: NP_001188537.1), DMR2 (Protein ID: NP_609942.1), and DMR3 (Protein ID: NP_611419.1) were codon-optimized for E. coli, synthesized by Genewiz, then ligated into vector pET28a between NdeI and EcoRI sites. To produce pCold-KshB, the sequence encoding KshB (Protein ID: WP_003900102.1) were codon-optimized for E. coli, synthesized by Genewiz, and ligated into the pCold I between NdeI and EcoRI sites. The sequence of “TDO” contains a reductase (TDO-R; Protein ID: WP_012052598.1) and a Rieske [2Fe–2S] cluster protein (TDO-F; Protein ID: WP_012052599.1)1 linked by peptide
EGRGSLLTCGDVEENPGP. To produce pCold-TDO, the gene encoding TDO was codon-optimized for E. coli, and synthesized by Genewiz, and ligated into vectors pET28a and pCold I between NdeI and EcoRI sites.
- S4 -
Plasmids encoding KshA, FDH, and various reductases including KshB, TDO, DMR1, DMR2, and DMR3, were transformed into E. coli BL21(DE3) cells, which were grown in lysogeny broth (LB) with antibiotics (100 μg/mL ampicillin or 50 μg/mL kanamycin) at 37 °C. When the optical density at 600 nm (OD600) reached 0.6-0.8, isopropyl β-D-1-thiogalactopyranoside (IPTG) was
added to a final concentration of 500 μM and the cells were then incubated at 16 °C overnight. The cells were harvested by centrifugation at 5,000 × g and resuspended in lysis buffer [20 mM Tris-HCl pH 8.0, 5 mM imidazole, 0.5 M NaCl, and 1 mM dithiothreitol (DTT)], and 1 mM phenylmethanesulfonyl fluoride (PMSF) and 0.5 mg/mL lysozyme were added. The cells were disrupted on ice by sonication using an ultrasonicator (SCIENTZ-950E, Ningbo Scientz Biotechnology Co. Ltd., Ningbo, China), followed by centrifugation at 40,000 × g at 4 °C for 30 min. The enzymes in cleared supernatants were trapped on 1 mL Ni-NTA Superflow resin (Qiagen, Hilden, Germany) and loaded onto a column. The resin was washed twice with 10 mL wash buffer (20 mM Tris-HCl, pH 8.0, 10 mM imidazole, 0.5 M NaCl, 1 mM DTT) and then target proteins were eluted with 10 mL elution buffer (20 mM Tris-HCl, pH 8.0, 300 mM imidazole, 0.5 M NaCl, 1 mM DTT).2,3 The target enzymes in elution buffer were exchanged into
phosphate-buffered saline (except for KshA, which was unstable).4,5 The protein concentration of each was then determined by BCA assay. The amount and purity of the target proteins was analyzed by SDS-PAGE followed by Coomassie Brilliant Blue staining.
Construction of modified KshB and TDO
The plant-type [2Fe–2S] cluster domain (pFeS, residues 298-358) of KshB was deleted from
kshB by PCR with the primers ∆KshB_F and ∆KshB_R (Table S2). The PCR product was digested with NdeI and EcoRI and cloned into the pCold I to generate plasmid ∆KshB. The
- S5 -
Rieske [2Fe–2S] cluster domain of TDO (residues 417-523) was removed using PCR with primers ∆TDO_F and ∆TDO_R. The product was digested with NdeI and EcoRI and cloned into the pCold I, which resulted in plasmid ∆TDO. A fragment encoding pFeS from KshB was amplified with primers of pFeS-KshB_F and pFeS-KshB_R. This was then inserted into vector pCold-KshB linearized with NdeI using the NEBuilder high-fidelity DNA assembly cloning kit following the manufacturer’s protocol to produce pFeS-KshB. A fragment encoding the Rieske [2Fe–2S] cluster domain (residues 417-523) of TDO was amplified with primers of TDO-F-KshB_F and TDO-F-KshB_R. It was then inserted into the pCold-KshB vector linearized with NdeI using the above cloning kit, to produce plasmid TDO-F-KshB. The DNA encoding pFeS was amplified with primers of pFeS-TDO_F and pFeS-TDO_R and inserted into the pCold-TDO linearized with NdeI, which created pFeS-TDO. The gene encoding TDO-F (i.e., residues 417–523 of TDO) was amplified with primers of TDO-F-TDO_F and TDO-F-TDO_R and inserted into pCold-TDO vector linearized with NdeI, which created plasmid TDO-F-TDO. All the constructed plasmids were confirmed by sequencing (performed by Genewiz). The plasmids were transformed into E. coli BL21 (DE3) for protein expression, as described above.
Construction of whole-cell catalysts for the transformation of AD
Whole-cell catalysis systems were constructed in the E. coli BL21(DE3) cells for stable and sustainable production of 9OHAD (Table S1). To construct strain BLKA-KB, pET28a harboring
kshA and pCold I carrying kshB were co-transformed into E. coli BL21(DE3). For strain BLKA-T, pET28a harboring kshA and pCold I carrying TDO were co-transformed into E. coli BL21(DE3). To construct strain BLKA-KB-F, the kshB gene was amplified, digested, and inserted into MCS1 of pETDuet-1 between EcoRI and HindIII sites. The gene encoding FDH was inserted into MCS2,
- S6 -
between NdeI and XhoI, which created pETDuet-1-KshB-FDH. pETDuet-1-KshB-FDH and pET28a harboring kshA were co-transformed into E. coli BL21(DE3) cells. For the construction of strain BLKA-T-F, the TDO-encoding gene was amplified, digested, and inserted into MCS1 of pETDuet-1 between EcoRI and HindIII sites. The gene encoding FDH was inserted into the same position in MCS2 as in pETDuet-1-TDO-FDH, which created pETDuet-1-TDO-FDH. pETDuet-1-TDO-FDHand pET28a harboring kshA were co-transformed into E. coli BL21(DE3) cells. For the construction of strain BLKA-TM-F, a gene encoding TDO mutant MT9 replaced the gene encoding TDO in pETDuet-1-TDO-FDH to generate pETDuet-1-TDOMT9-FDH. The latter plasmid was co-transformed with pET28a harboring kshA into E. coli BL21(DE3) cells. For the construction of strain BLKA-RTM-F, the gene encoding TDO-F-TDOMT9 was constructed based on strain BLKA-T-F above, which created pETDuet-1-TDO-F-TDOMT9-FDH. This plasmid and pET28a harboring kshA were co-transformed into E. coli BL21(DE3) cells. For the construction of strain BLKA-KBF, the FDH-encoding gene was inserted into the pCold-KshB linearized with EcoRI, which created pCold-KshB-FDH. pCold-KshB-FDH plasmid and pET28a harboring kshA were co-transformed into E. coli BL21(DE3) cells. To construct strain BLKA-TF, the gene encoding FDH was inserted into the pCold-TDO plasmid as above, which created pCold-TDO-FDH, which was co-transformed with pET28a harboring kshA into E. coli
BL21(DE3). For the construction of strain BLKB-KA-F, kshA was amplified, the fragment was digested, and it was inserted into MCS2 of pETDuet-1-KshB between NdeI and XhoI sites, which created pETDuet-1-KshB-KshA. pETDuet-1-KshB-KshA and pET28a harboring FDH were co-transformed into E. coli BL21(DE3) cells. To construct strain BLT-KA-F, kshA was amplified, digested, and inserted into MCS2 of pETDuet-1-TDO between NdeI and XhoI sites, which created pETDuet-1-TDO-KshA. The latter plasmid and pET28a harboring FDH were
- S7 -
co-transformed into E. coli BL21(DE3) cells. All the transformants mentioned above were selected using 100 μg/mL ampicillin and 50 μg/mL kanamycin.
Bioconversion of AD to produce 9OHAD
The constructed recombinant strains listed in Table S1 were grown in lysogeny broth (LB) at 37 °C containing 100 μg/mL ampicillin or 50 μg/mL kanamycin. IPTG was added to a final concentration of 0.5 mM when the OD600 reached 0.6–0.8, followed by incubation at 16 °C
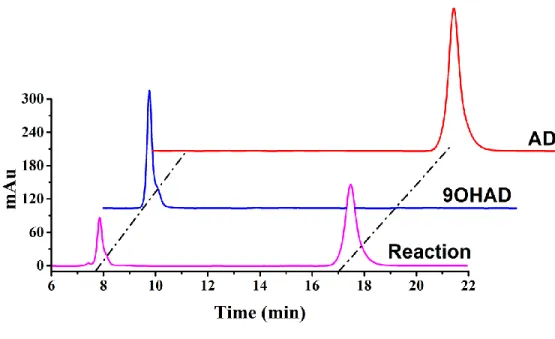
overnight. The cells were harvested by centrifugation at 8,000 × g for 15 min and resuspended in 30 mL 50 mM PBS (pH 7.4) in 250-mL Erlenmeyer flasks. The production of 9OHAD was conducted in reaction mixture containing 22.8 g/L methyl-β-cyclodextrin, 50 g/L wet cells, 5 g/L AD, and 0.12 g/L sodium formate at 35 °C. Products were determined by high-performance liquid chromatography.
Structural modeling of KshB
Modeller 9.9 software was used to generate a homology model of KshB.6 The crystal structure
of phthalate dioxygenase reductase from Pseudomonas cepacia (PDB ID: 3VYL), which has 30.0% sequence identity to KshB, was used as the template.7 The model was evaluated by
Ramachandran analysis; the favoured and allowed zones represented 93.1% and 2.9%,8
respectively, which suggested that the homology model was reasonable. The generated model structure was visualized and analyzed using the PyMol Molecular Graphics System (http://www.pymol.org).9
- S8 -
Differential scanning fluorimetry experiments were performed using a CFX96 Real-Time PCR system (Bio-Rad, Hercules, CA, USA) with an excitation wavelength of 492 nm and emission wavelength of 610 nm.10 SYPRO Orange (Ex/Em: 470/570 nm; Invitrogen, Carlsbad, CA, USA)
was used as the reporter dye. Samples were prepared in microplate wells containing 22.5 μL of 5 μM purified protein, 1mM DTT, and 2.5 μL of a 25-fold dilution of SYPRO Orange. Then, the samples were sealed with Optical-Quality Sealing Tape and heated in the Real-Time PCR system. The samples were denatured using a linear gradient from 20 to 95 °C at 0.5 °C per step (30 s). Protein unfolding was monitored by detecting changes in SYPRO Orange fluorescence. The inflection point of curves of fluorescence vs temperature was identified by plotting the first derivative against temperature in CFX Manager Software (Bio-Rad), and the minimum value was referred to as the melting temperature.
- S9 - Table S1 Bacterial strains used in this study
Strains Description Source
E. coli JM109 Host of plasmid for cloning Lab stock
E. coli BL21 (DE3) Host of plasmid for expression Lab stock
BLKA-KB pET28a carries KshA, pCold carries KshB; BL21 (DE3) This study
BLKA-T pET28a carries KshA, pCold carries TDO; BL21 (DE3) This study
BLKA-KB-F pET28a carries KshA, pETDuet carries KshB and FDH; BL21 (DE3) This study BLKA-T-F pET28a carries KshA, pETDuet carries TDO and FDH; BL21 (DE3) This study BLKA-TM-F pET28a carries KshA, pETDuet carries TDOMT9 and FDH; BL21 (DE3) This study BLKA-RTM-F pET28a carries KshA, pETDuet carries TDO-F-TDOMT9 and FDH;
BL21 (DE3)
This study BLKA-KBF pET28a carries KshA, pCold carries KshB and FDH; BL21 (DE3) This study BLKA-TF pET28a carries KshA, pCold carries TDO and FDH; BL21 (DE3) This study BLKB-KA-F pET28a carries FDH, pETDuet carries KshB and KshA; BL21 (DE3) This study BLT-KA-F pET28a carries FDH, pETDuet carries TDO and KshA; BL21 (DE3) This study
- S10 - Table S2. Primers and plasmids used in this study.
Primers and
plasmids Sequence of primer
Primers
∆KshB_F GGAATTCCATATGATGTTGACCGAGGCAATTGGAGACG
∆KshB_R GGAATTCTTAGCCCGCGGCCAGCAGCACG pFeS-KshB_F CATCATATCGAAGGTAGGCATATGGGCCTGGACGCGCCG pFeS-KshB_R TCCAATTGCCTCGGTCAACATTGGGCCAGGATTCTCCTCCACGTC ACCGCATGTTAGGAGACTTCCTCTGCCCTCCTCGTCGTAGGTCAC TTCCACCGAATCAG pFeS-TDO_R GATGGCAACATGGGTGGCCATTGGGCCAGGATTCTCCTCCACGTC ACCGCATGTTAGGAGACTTCCTCTGCCCTCCTCGTCGTAGGTCAC TTCCACCGAATCAG KshB_F CGGAATTCGATGTTGACCGAGGCAATTGG KshB_R CCCAAGCTTTTACTCGTCGTAGGTCACTTCCAC
KshA_F GGAATTCCATATGAGCACCGATACGAGCGG
KshA_R CCGCTCGAGTTAGTGCTGCTCGGCCGG
FDH_F GGAATTCCATATGAGATCGTGCTGGTGCTGTATGATGC
FDH_R CCGCTCGAGTTATTTTTTATCATGCTTGCGTATGCCTTGGTC
∆TDO_F GGAATTCCATATGGCCACCCATGTTGCCATCATC
∆TDO_R GGAATTCTTATGTCAGATCACCTTCATTTGCGCGC
TDO-F-KshB_F CATCATATCGAAGGTAGGCATATGACTTGGACCTATATTTTAC TDO-F-KshB_R TCCAATTGCCTCGGTCAACATTGGGCCAGGATTCTCCTCCACGTC ACCGCATGTTAGGAGACTTCCTCTGCCCTCTTTCAGTTCACCGTTA TC TDO-F-TDO_R GATGGCAACATGGGTGGCCATTGGGCCAGGATTCTCCTCCACGTC ACCGCATGTTAGGAGACTTCCTCTGCCCTCTTTCAGTTCACCGTTA TC
TDO_F GGAATTCGATGGCCACCCATGTTGCCATCATC
TDO_R CCCAAGCTTTTATTTCAGTTCACCGTTATCTAAATC
TDOMT_F CATCATATCGAAGGTAGGCAATGGCCACCCATGTTGCC
TDOMT_R CTCGAGGGTACCGAGCTCCATTATTTCAGTTCACCGTTATCTAAAT
CAACG
RTDOMT_F ATCACCACAGCCAGGATCCGATGACTTGGACCTATATTTTAC
- S11 - FDH-KshB_F AGATAACGGTGAACTGAAAGAGGGCAGAGGAAGTCTCCTAACAT GCGGTGACGTGGAGGAGAATCCTGGCCCAATGAAGATCGTGCTG GTG FDH-KshB_R ACTGCAGGTCGACAAGCTTGTTATTTTTTATCATGCTTGCCG FDH-TDO_F AGTGACCTACGACGAGGAGGGCAGAGGAAGTCTCCTAACATGCG GTGACGTGGAGGAGAATCCTGGCCCAATGAAGATCGTGCTGGTG Plasmids
pET28a-KshA pET28a, contain KshA gene (codon-optimized), KanR
pET28a-FDH pET28a, contain FDH gene (codon-optimized), KanR
pET28a-DMR1 pET28a, contain DMR1 gene (codon-optimized), KanR
pET28a-DMR2 pET28a, contain DMR2 gene (codon-optimized), KanR
pET28a-DMR3 pET28a, contain DMR3 gene (codon-optimized), KanR
pCold-KshB pCold I, contain KshB gene (codon-optimized), AmpR
pET28a-TDO pET28a, contain TDO gene (codon-optimized), KanR
- S12 -
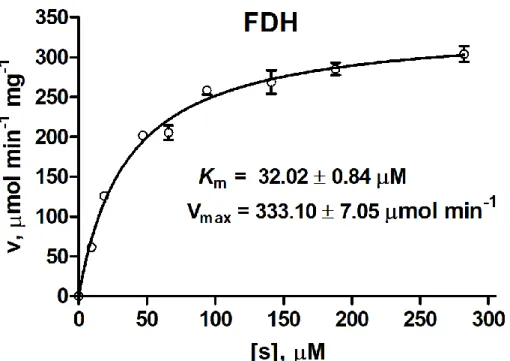
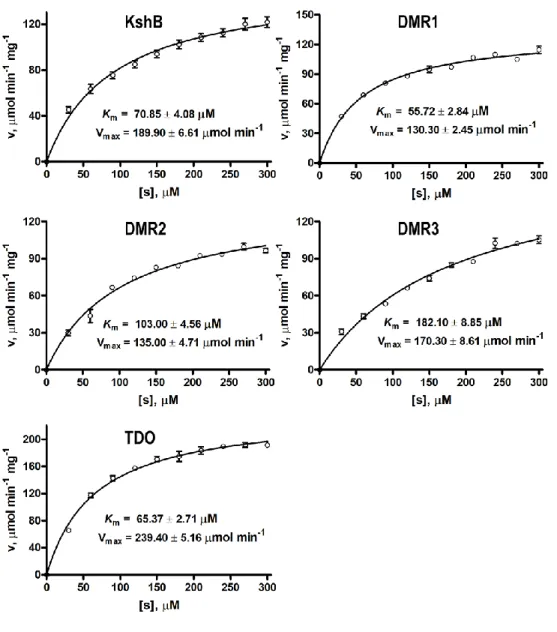
Figure S1 TheMichaelis-Menten plots of FDH toward NAD+. Data are shown as the mean ± SD
- S13 -
Figure S2 The Michaelis-Menten plots of reductases toward NADH. Data are shown as the mean ± SD from three independent experiments.
- S14 -
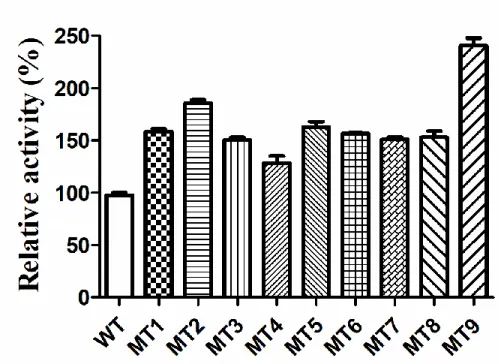
Figure S3 The relative activity of TDO mutants by error-prone PCR. Data are shown as the mean ± SD from three independent experiments.
- S15 -
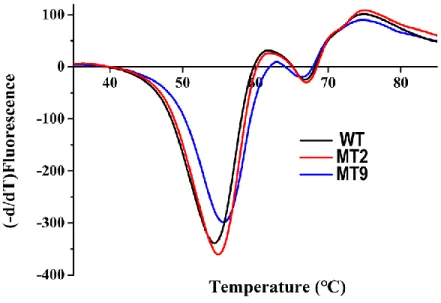
Fig. S4 Melting curves of wild-type and mutant TDO measured using differential scanning fluorimetry.
- S16 -
Fig. S5 The relative activity of modified [2Fe-2S] cluster in KshB and TDO toward NADH. The relative activity of KshB was set as 100%. Data are shown as the mean ± SD from three independent experiments.
- S17 -
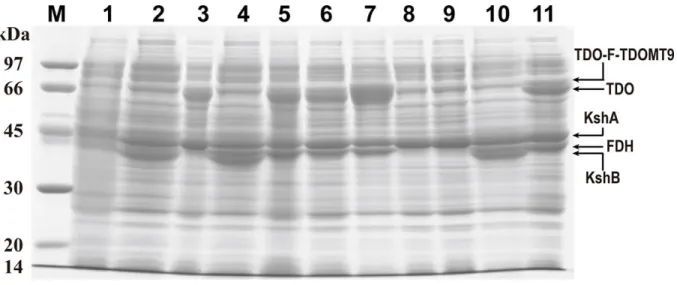
Fig. S6 SDS-PAGE analysis of the enzymes expressed in E. coli BL21(DE3) cells. M: protein marker; lane 1, control: E. coli; lane 2, BLKA-KB; lane 3, BLKA-T; lane 4, BLKA-KB-F; lane 5, BLKA-T-F; lane 6, BLKA-TM-F; lane 7, BLKA-RTM-F; lane 8, BLKA-KBF; lane 9, BLKA-TF; lane 10, BLKB-KA-F; lane 11, BLT-KA-F.
- S18 -
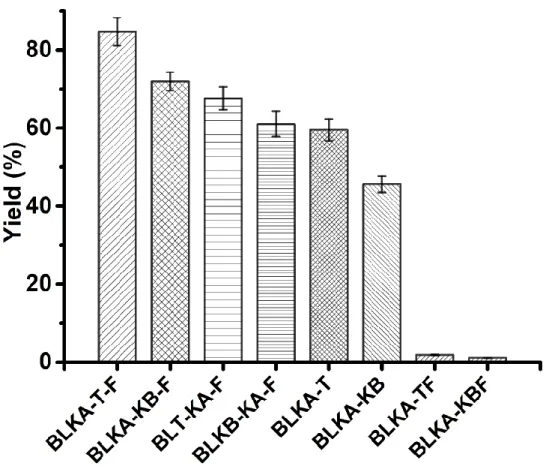
Figure S7 The yields of 9OHAD using different strains containing different plasmids. Data are shown as the mean ± SD from three independent experiments.
- S19 -
- S20 -
References
(1) Friemann, R.; Lee, K.; Brown, E. N.; Gibson, D. T.; Eklund, H.; Ramaswamy, S. Structures of the multicomponent Rieske non-heme iron toluene 2,3-dioxygenase enzyme system.
Acta Crystallogr. D: Biol. Crystallogr. 2009, 65, 24-33.
(2) Qin, H. M.; Miyakawa, T.; Inoue, A.; Nishiyama, R.; Nakamura, A.; Asano, A.; Ojima, T.; Tanokura, M. Structural basis for controlling the enzymatic properties of polymannuronate preferred alginate lyase FlAlyA from the PL-7 family. Chem. Commun.2018, 54, 555-558. (3) Qin, H. M.; Miyakawa, T.; Inoue, A.; Nishiyama, R.; Nakamura, A.; Asano, A.; Ojima, T.;
Tanokura, M. Structure and polymannuronate specificity of a eukaryotic member of polysaccharide lyase family 14. J. Biol. Chem.2017, 292, 2182-2190.
(4) Liu, H. H.; Xu, L. Q.; Yao, K.; Xiong, L. B.; Tao, X. Y.; Liu, M.; Wang. F. Q.; Wei, D. Z. Engineered 3-ketosteroid 9α-hydroxylases in Mycobacterium neoaurum: an efficient platform for production of steroid drugs. Appl. Environ. Microbiol. 2018, 84, e02777-02717.
(5) Petrusma, M.; van der Geize, R.; Dijkhuizen, L. 3-Ketosteroid 9α-hydroxylase enzymes: Rieske non-heme monooxygenases essential for bacterial steroid degradation. Antonie van Leeuwenhoek2014, 106, 157-172.
(6) Šali, A.; Blundell, T. L.; Comparative protein modelling by satisfaction of spatial restraints.
J. Mol. Biol.1993, 234, 779-815.
(7) Correll, C. C.; Batie, C. J.; Ballou, D. P.; Ludwig, M. L. Phthalate dioxygenase reductase: a modular structure for electron transfer from pyridine nucleotides to [2Fe-2S]. Science1992, 258, 1604-1610.
(8) Laskowski, R. A.; MacArthur, M. W.; Moss, D. S.; Thornton, J. M. ProCheck: a program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283-291.
(9) Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33-38.
(10) Niesen, F. H.; Berglund, H.; Vedadi, M., The use of differential scanning fluorimetry to detect ligand interactions that promote protein stability. Nat Protoc.2007, 2, 2212-2221.
![Fig. S5 The relative activity of modified [2Fe-2S] cluster in KshB and TDO toward NADH](https://thumb-us.123doks.com/thumbv2/123dok_us/1872923.2773402/16.918.246.676.104.461/fig-relative-activity-modified-cluster-kshb-tdo-nadh.webp)