Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Molecular Analysis of
Mycobacterium avium
Isolates by Using
Pulsed-Field Gel Electrophoresis and PCR
MARTINE PESTEL-CARON,1* GABRIEL GRAFF,1GILLES BERTHELOT,2JEAN-LOUIS PONS,1
ANDJEAN-FRANC¸OIS LEMELAND1
Groupe de Recherche sur les Antimicrobiens et les Micro-organismes (GRAM, EA 2656), 76000 Rouen,1
and Laboratoire de Bacte´riologie, Centre Hospitalier Ge´ne´ral, 76202 Dieppe Cedex,2France
Received 11 January 1999/Returned for modification 12 March 1999/Accepted 7 May 1999
Genetic relationships among 46 isolates ofMycobacterium aviumrecovered from 37 patients in a 2,500-bed hospital from 1993 to 1998 were assessed by pulsed-field gel electrophoresis (PFGE) and PCR amplification of genomic sequences located between the repetitive elements IS1245and IS1311. Each technique enabled the identification of 27 to 32 different patterns among the 46 isolates, confirming that the genetic heterogeneity of M. aviumstrains is high in a given community. Furthermore, this retrospective analysis of sporadic isolates allowed us (i) to suggest the existence of two remanent strains in our region, (ii) to raise the question of the possibility of nosocomial acquisition ofM. aviumstrains, and (iii) to document laboratory contamination. The methods applied in the present study were found to be useful for the typing ofM. aviumisolates. In general, both methods yielded similar results for both related and unrelated isolates. However, the isolates in five of the six PCR clusters were distributed among two to three PFGE patterns, suggesting that this PCR-based method may have limitations for the analysis of strains with low insertion sequence copy numbers or for resolution of extended epidemiologic relationships.
Despite the common occurrence of disseminated
Myco-bacterium aviumcomplex disease in patients with AIDS, the
epidemiology of this infection is incompletely understood. No-tably, the predominant source of infection and whether dis-seminatedM. aviumcomplex infection results from reactiva-tion or recent acquisireactiva-tion of infecreactiva-tion in human AIDS patients remain unclear. It has recently been demonstrated that water distribution systems may be colonized withM. avium(18) and may subsequently serve as a potential source of infection for AIDS patients (43). However, in contrast to these findings, some epidemiologic and clinical studies have failed to find an association between specific environmental sources and human infection (16, 44). These conflicting results may, in particular, illustrate the need for suitable epidemiologic markers for in-vestigation of the sources ofM. aviuminfections as well as the routes of transmission, especially because of the large numbers of potential sources for human exposure.
Different laboratory methods, including serotyping (41), multilocus enzyme electrophoresis (45), restriction fragment length polymorphism (RFLP) analysis and hybridization to specific probes (4, 10, 11, 14, 15, 19, 29, 32, 33), and pulsed-field gel electrophoresis (PFGE) (2, 4, 7, 23, 24, 36), have been applied for these purposes. The last two methods mentioned are DNA-based methods and typically use agarose gel electro-phoresis of restriction enzyme-digested genomic DNA which is stained directly with ethidium bromide (PFGE) or which is transferred to membranes and probed with labeled DNA (RFLP analysis). Both techniques are relatively slow and la-bor-intensive (especially forM. avium, whose slow growth can delay the time to retrieval of results), requiring DNA of high integrity and at high concentrations. More recently, PCR-based typing methods have been described (22, 25, 31). The application of PCR to the molecular typing of bacterial species
offers the potential for a relatively simple and inexpensive means of typing bacterial isolates for epidemiologic purposes. One of them, described by Picardeau and Vincent (31), used primers that bound to the ends of insertion elements specific
forM. avium(IS1245and IS1311), thus amplifying the DNA
between closely spaced copies of these elements.
We investigated the genetic relationships of all M. avium
isolates consecutively recovered from patients in the Rouen university hospital from 1993 to 1998 and a few isolates from patients in two smaller hospitals in the neighboring area by PFGE and PCR typing as described by Picardeau and Vincent (31). The aim of the study was (i) to characterize the genetic diversity of theM. aviumstrains from the Rouen hospital, (ii) to investigate whether nosocomial acquisition of M. avium
infection either by cross-contamination or by exposure to a common source occurred in our large urban teaching hospital, and (iii) to evaluate whether this PCR-based method is reliable for typing and longitudinal analysis of large numbers of iso-lates.
MATERIALS AND METHODS
M. aviumisolates.Forty-six isolates, including 35 isolates recovered from 26 patients with AIDS and 11 isolates from 11 human immunodeficiency virus (HIV)-uninfected patients, were studied. These isolates consisted of allM. avium
isolates collected in the Rouen university hospital from March 1993 to March 1998 (40) and of other isolates initially cultured in two different hospitals in the area (a hospital in Dieppe, France, five isolates; a hospital in Evreux, France, one isolate). Among these 46 isolates were 13 sequential isolates cultured from identical or nonidentical sites from four patients (at intervals ranging from 8 to 670 days). The 46M. aviumisolates were cultured from sterile sites (blood [n⫽
25 isolates], bone marrow [n⫽2], lymph node [n⫽2], bladder [n⫽1], and cutaneous biopsy specimen [n⫽1]) and from nonsterile sites (gastric aspirates [n⫽2], bronchopulmonary specimens [n⫽11], and cutaneous specimens [n⫽
2]). In this study, isolates recovered from different patients were considered to be unrelated, and sequential isolates obtained from a single patient over weeks were considered to be related.
Isolates were identified asM. aviumon the basis of conventional biochemical tests and by PCR-restriction enzyme pattern analysis of thehsp65gene (37).
PFGE.M. aviumisolates were grown in 10 ml of Middlebrook 7H9 broth
supplemented with 0.5 M sucrose–0.05% Tween 80–10% oleic acid-albumin-dextrose until they reached an optical density of 0.250 at 650 nm. Plugs were prepared and digested as described previously (21, 36) with 25 U ofAseI (New
* Corresponding author. Mailing address: Laboratoire de Bacte´ri-ologie, Centre Hospitalier Universitaire Charles Nicolle, 76031 Rouen Cedex, France. Phone: 33. 2.32.88.80.52. Fax: 33. 2.32.88.80.24. E-mail: [email protected].
2450
on May 15, 2020 by guest
http://jcm.asm.org/
England BioLabs, Beverly, Mass.). Large restriction fragments were separated in a 1% agarose gel (SeaKem GTG; FMC BioProducts, Rockland, Maine) at 14°C for 19.7 h by using the Gene Path system (Bio-Rad Laboratories, Ivry/Seine, France). The patterns were visualized under UV light and were digitized with the Gel Doc 1000 documentation system (Bio-Rad Laboratories). PFGE fingerprints were analyzed by applying the Dice coefficient to peaks. For clustering, the unweighted pair group method with arithmetic means was used. A tolerance in the band positions of 1.2% was applied for comparison of the fingerprint pat-terns. Fingerprint analysis and the methods and algorithms used in this study were performed according to the instructions of the manufacturer. Bacterio-phage lambda DNA concatemers (New England BioLabs) were included as molecular weight standards with each run.
PCR.The PCR typing method used in the study was a variation of a previously reported procedure (31), with specific modifications made to simplify the extrac-tion of DNA from mycobacteria. Briefly, one colony ofM. aviumwas taken from Middlebrook 7H10 plates and was suspended in 20l of TE buffer (10 mM Tris, 0.1 mM EDTA [pH 7.6]) containing 1% Triton X-100. Five microliters of this suspension was submitted to a lytic cycle directly in the amplification tube of a GeneAmp PCR system 2400 (Perkin-Elmer Cetus, Norwalk, Conn.) as described previously (3). Subsequently, 45l of the PCR reagent mixture was added to the PCR tube to initiate amplification. The PCR mixture and the amplification reactions were performed as described by Picardeau and Vincent (31). All experiments included negative controls which were processed with the samples. Amplification products were electrophoresed on a 1.5% agarose gel (SeaKem LE; FMC BioProducts) and were detected by ethidium bromide staining. Gels were photographed with UV illumination, and band patterns were compared visually.
Reproducibility and discriminatory power of PFGE and PCR.A total of 13
and 20 isolates were studied in duplicate by PFGE and PCR, respectively, to assess the reproducibilities of the PFGE and PCR patterns in our hands. Re-producibility was defined as the percentage of pairs with identical patterns. The discriminatory power was calculated as described by Hunter and Gaston (17) on the basis of the patterns obtained with the 37 unrelated isolates.
RESULTS
PFGE. PFGE after AseI digestion of chromosomal DNA revealed 32 distinct banding patterns, according to the inter-pretive criteria of Tenover et al. (39), among the 46 isolates collected over 5 years from 37 patients. Of these 32 PFGE patterns, 25 were unique. The patterns observed were poly-morphic and complex, including 10 to 18 fragments ranging from 35 to 900 kb (Fig. 1). The reproducibility rate of PFGE was 100%.
Isolates obtained from different specimens collected from the same patient were compared. For two of the four
HIV-infected patients studied, all sequential isolates had identical PFGE profile (patterns P1 and P3); for the third patient, one isolate differed from the two others by only a single band consistent with a single genetic event (Dice coefficient, 96%) (Fig. 2). Such minor variation was considered consistent with variation within a strain (pattern P2) (Fig. 2). For the fourth HIV-infected patient studied, the patterns of the two isolates cultured 668 and 670 days, respectively, after culture of the initialM. aviumisolate presented up to six band differences compared with the original profile. Thus, they were considered possibly related to the first isolate and their profiles were designated subtypes of the initial profile (pattern P6) (39). Such variations have been reported among isolates collected over a long period of time (ⱖ6 months) (39). Thus, for all four patients, we explicitly documented clonally disseminatedM.
aviuminfections, with three patients infected with a strain at
multiple sites.
[image:2.612.55.290.71.261.2]Among the patterns observed, one (pattern P4) was strictly identical for three isolates obtained from one urine and two bronchopulmonary samples from three HIV-negative patients monitored in three different units of the hospital in Dieppe. Their place of residence was not the same, but their clinical specimens were received in the laboratory on the same day,
FIG. 1. Restriction patterns fromAseI digests ofM. aviumisolates resolved by PFGE. Lanes: 1 and 6, bacteriophage lambda DNA concatemers (sizes [in kilobases] are indicated on the left); 2, isolate 100A8; 3 and 4, pattern P7 (isolates 100A28 and 100A32, respectively); 5, isolate 100A25; 7 to 11, five sequential isolates from one patient, respectively (pattern P1); 12 to 15, four isolates from two patients, respectively (pattern P2).
FIG. 2. Dendrogram of PFGE fingerprints of 46M. aviumisolates as deter-mined by the Dice method. Brackets indicate identical or closely related pat-terns.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:2.612.320.548.326.698.2]suggesting either laboratory contamination or nosocomial ac-quisition of the isolates. On the other hand, three particular clusters (patterns P2, P5, and P7) of two to four isolates each were defined according to the interpretive criteria of Tenover et al. (39). These clusters comprised isolates that were consid-ered to be closely related because they exhibited very close PFGE patterns which differed by only one or two DNA frag-ments (Dice coefficients, 92 to 96%) (Fig. 2). Each of these clusters consisted of clinical isolates cultured from two patients who either attended the same hospital but at different times (from 3 to 21 months apart) or were monitored at different study sites (hospitals in Rouen and Dieppe) at different peri-ods of time (18 months apart).
The patterns exhibited by the 11 isolates collected from HIV-negative patients were distributed throughout the den-drogram.
PCR.The same 46 isolates were also analyzed by a PCR-based typing technique with primers directed at the conserved inverted repeats of IS1245and IS1311. This PCR was designed to amplify DNA segments between multiple copies of these elements, resulting in a mycobacterial strain-specific banding profile (31). These two primers generated PCR banding pat-terns with DNAs from all M. aviumisolates included in the study. Twenty-seven profiles were observed among the 46 iso-lates. The PCR profiles were relatively diverse (Fig. 3). Band-ing patterns consisted of fewer than 10 bands rangBand-ing from 350 to 2,900 bp, with some corresponding to intense bands and others corresponding to weaker bands (Fig. 3). The PCR pro-files were identical for strains isolated from the same patient (including isolates from different body sites) and were different for the majority of the strains from different patients. Twenty-one isolates had unique PCR patterns, and six profiles (labeled profiles A to F) with up to six bands were observed for two or more isolates. Most of the isolates included in the same PCR cluster had the same minor bands; the exception was for cluster D, the two isolates of which differed by one reproducible minor band, suggesting that these isolates were more likely closely related than identical (data not shown).
The reproducibility rate for this PCR based-typing method in our hands was 90% on the basis of different PCR tests with the same bacterial extracts of 20 isolates. Of note, for one isolate, an extra major band was apparent compared to the bands for other sequential isolates from the same patient (Fig. 4, lanes 7 and 9), but this band was no longer present when the
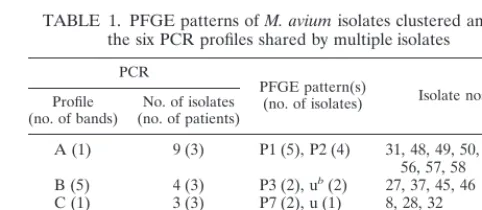
PCR was repeated on two separate occasions (data not shown). Comparison of typing methods.To evaluate the epidemio-logic value of PCR typing for isolates collected over a 5-year period, related and unrelated isolates were studied by both PFGE and PCR techniques. Analysis of the same 46 isolates by PCR yielded results similar to those of PFGE because of the similar banding patterns for isolates within most clusters de-tected by PFGE. One exception was for two isolates (isolates 100A13 and 100A26) that belonged to PFGE cluster P5 but that exhibited clearly different PCR profiles (Fig. 4, lanes 15 and 16). On the other hand, most of the unrelated isolates with distinct PFGE patterns had distinct PCR profiles. However, even if the numerical index of discriminatory power of both methods was 0.98, the isolates clustered in five of the six PCR profiles common to multiple isolates were distributed among two or three distinct PFGE patterns (Dice coefficients, 40 to 67%) (Fig. 2; Table 1). The DNAs of isolates with these five PCR profiles exhibited one to five bands (Table 1).
DISCUSSION
[image:3.612.55.290.73.227.2]Precise definition of the epidemiology and mode of trans-mission of infectious bacterial diseases requires both detailed
[image:3.612.312.548.74.229.2]FIG. 3. PCR typing of clinicalM. aviumisolates. Lanes: 1 to 5, 7 to 11, and 13 to 17, patterns of isolates obtained from 15 unrelated patients; lanes 6 and 18, bacteriophage lambda DNA-BstEII digest molecular weight marker; and lane 12, pBR322 DNA-MspI digest (New England Biolabs).
FIG. 4. Electrophoretic PCR patterns forM. aviumisolates. Lanes 6 and 18, bacteriophage lambda DNA-BstEII digest; lane 12, pBR322 DNA-MspI digest (New England Biolabs); lanes 1 to 5, five sequential isolates from one patient, respectively (PCR profile A, PFGE pattern P1); lanes 7, 8, and 9, three sequen-tial isolates from one patient, respectively (PCR profile A, PFGE pattern P2) (no amplification product was detected in lane 8 in that experiment); lane 10, isolate 100A31 from another patient (PCR profile A, PFGE pattern P2); lanes 11 and 13, two sequential isolates from one patient, respectively; lane 14, isolate 100A7 from a different patient (PCR profile E, PFGE pattern P6 or a unique pattern); lanes 15, 16, and 17, isolates 100A13, 100A26, and 100A30, respectively.
TABLE 1. PFGE patterns ofM. aviumisolates clustered among the six PCR profiles shared by multiple isolates
PCR
PFGE pattern(s)
(no. of isolates) Isolate nos.a Profile
(no. of bands) (no. of patients)No. of isolates
A (1) 9 (3) P1 (5), P2 (4) 31, 48, 49, 50, 51, 52, 56, 57, 58 B (5) 4 (3) P3 (2), ub(2) 27, 37, 45, 46
C (1) 3 (3) P7 (2), u (1) 8, 28, 32 D (4) 2 (2) u (2) 34, 38 E (4) 4 (2) P6 (3), u (1) 7, 17, 18, 19 F (6) 3 (3) P4 (3) 41, 42, 43
aThe prefix for all isolate designations is 100A. bu, unique PFGE pattern.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:3.612.311.552.606.711.2]clinical and epidemiologic data and an effective method (which meets the criteria of typeability, reproducibility, and discrimi-natory power [1, 20]) for the differentiation of different strains of the organism concerned. The analysis of multiple isolates from each infected patient can also provide insights into the pathogenesis of infection (2, 36).
The aim of the present study was to analyze retrospectively related and unrelatedM. aviumisolates collected over a 5-year period from patients in the same hospital, to define the local epidemiology ofM. avium infections, and to expand our un-derstanding of the local chain of transmission ofM. avium.
Among all the various approaches previously attempted for the typing ofM. aviumisolates, PFGE has been proposed as the “gold standard” (1, 2, 20, 24, 38) because of its highly discriminatory and reproducible results. However, because this method remains laborious and time-consuming, especially when it is applied to mycobacteria, the evaluation of other techniques is appreciated. For this reason, we analyzed our collection of 46 isolates ofM. avium recovered from 1993 through 1998 by two techniques that rely on independent mo-lecular markers: PFGE afterAseI digestion of the chromo-somal DNA and a PCR-based technique with oligonucleotide primers against the inverted repeats of IS1245and IS1311(31). Picardeau and colleagues (30, 31) have previously shown that this PCR typing technique is rapid and simple and is as discriminatory as RFLP analysis. In agreement with their find-ings, in our hands, this typing system provided reproducible and easy-to-analyze patterns comprising fewer than 10 bands. Faint bands were taken into account only when they were reproducible in different PCR tests. Indeed, there were varia-tions in some of the minor and/or major PCR products, which may make the comparison of large number of strains difficult. One of the most critical limitations, which was associated with a low insertion sequence copy number, is that the correspond-ing patterns, which consisted of only one band, are poorly discriminatory for epidemiologically unrelated isolates that represent distinct strains as resolved by PFGE (Dice coeffi-cients, 40 to 62%). These observations are analogous to those obtained by IS1245Southern blot analysis (8, 10, 11, 29, 32) and to those reported by Ross and Dwyer (34) from their analysis of two strains with one IS6110copy by a similar PCR-based method that relied on the amplification of DNA frag-ments between IS6110copies. On the other hand, this PCR typing method seems to have a lower discriminatory power than PFGE (despite a good numerical index that was calcu-lated according to the recommendations of Hunter and Gaston [17]) because five sets of isolates clustered in one PCR pattern were distributed among two or three PFGE patterns, and the two isolates in one PFGE cluster pattern (pattern P5) had two distinct PCR patterns.
Except for these limitations, the comparison of the two fin-gerprinting methods revealed that the banding patterns were similar within each cluster and were distinct from those for strains from different clusters.
Both typing techniques performed in the present study dem-onstrated the heterogeneity of theM. aviumspecies by the high number (32 PFGE patterns and 27 PCR profiles) and high degree of diversity of the patterns observed for the 37 unre-lated isolates. This is consistent with the results of previous studies based on PFGE, RFLP analysis with repetitive inser-tion sequences as DNA probes, or PCR (2, 6, 14, 24, 30, 31, 33, 36). No prevalent strain was identified among HIV-infected patients, and the patterns of the isolates from HIV-negative patients were diverse, too. This marked polymorphism con-trasts with the similarity between isolates obtained from the same patient over time, which indicates monoclonal infections.
Among the 37 unrelatedM. aviumisolates included in our study, PFGE and/or PCR analyses defined four clusters (clus-ters P2, P4, P5, and P7) of identical or closely genetically related isolates recovered from two patients. The information collected for patients infected with isolates in clusters P2 and P5 indicated that there was no epidemiologic link between the two patients. Under these conditions, the identification of two strains collected a few years apart and/or in different cities could suggest that someM. aviumstrains could be maintained in a population and/or in the same geographic area for several years, as reported previously for twoM. aviumstrains collected for up to almost 4 years in the recirculating hot water systems of two hospitals (43). This has also been described for other bacteria such as isolates of Staphylococcus aureus that were genotypically identical and that were recovered over a long period of time from unrelated patients (28, 35). With respect to cluster P7, because the two patients concerned attended the same facility at the same hospital over a 3-month period but also lived in cities that were close to each other, many hypoth-eses can be evoked, including exposure to an unidentified com-mon (nosocomial [43] or not nosocomial) environmental source or direct transmission from patient to patient, even if the latter has never been reported (6, 18, 26). Cluster P4 included clinical isolates collected on the same day from three HIV-negative patients (patients 1, 2, and 3) attending three different medical units of the same hospital during the same period. A nosocomial outbreak or laboratory contamination, as reported previously (5), could therefore be suspected. The retrospective review of the bacteriological data for the three patients revealed that three urine samples collected 1 day apart from patient 1 yielded multipleM. aviumcolonies 21 to 32 days after the inoculation of the solid media, consistent with a high likelihood of trueM. aviuminfection. In contrast, retrospective assessment of the bacteriological and clinical significance of the isolation of M. avium from patients 2 and 3 failed to suggest a role for these isolates as pathogens. No contamina-tion at the time of sample colleccontamina-tion can be suspected because this step was performed by different persons in each unit. In contrast, a laboratory contamination event remains a likely explanation for cluster P4. Indeed, the two identicalM. avium
isolates from patients 2 and 3 cultured over a long time (51 to 75 days) may be the result of cross-contamination from the positive urine sample from patient 1 since the samples from patients 2 and 3 were received and sequentially processed at the laboratory on the same day as the sample from patient 1. The two other M. avium strains (strains 100A26 and 100A39) isolated in this laboratory from 1993 to 1998 had clearly distinct PCR and PFGE profiles (Dice coefficient, 43%), suggesting that this contamination was self-limited. The impact of this contamination was much lower than that re-ported previously (5, 9, 12, 13, 27, 40, 42) as the result of a dysfunction of the BACTEC system or a low inoculum ofM.
aviumin medium additives from an exogenous source or other
hospital or laboratory sources, which generated large pseudo-outbreaks.
In summary, the one-band patterns and the variations ob-served in some of the minor and/or major bands could make the comparison of large numbers of isolates by the PCR-based technique used in this study difficult. We therefore recommend that this rapid technique, which does not need a tedious DNA preparation step in particular, could be used to investigate small numbers of isolates collected over a short period of time or for preliminary screening (especially for investigation of several colonies from a single strain), whereas PFGE remains the reference technique for strain characterization and seems more suitable for large-scale studies.
on May 15, 2020 by guest
http://jcm.asm.org/
The application of molecular techniques such as PFGE and PCR enabled us (i) to investigate the genetic diversity of theM.
aviumstrains present in a given community, (ii) to identify the
existence of possible remanent strains in our particular given region, (iii) to raise the question of the nosocomial acquisition of anM. aviumstrain, and (iv) to document laboratory con-tamination. Thereby, this study confirms how useful molecular strain typing can be in investigations of the genetic relation-ships ofM. aviumisolates collected in a given community.
ACKNOWLEDGMENT
We gratefully thank R. D. Arbeit for interest and helpful discus-sions.
REFERENCES
1.Arbeit, R. D.1999. Laboratory procedures for the epidemiologic analysis of
microorganisms, p. 116–137.InP. R. Murray, E. J. Baron, M. A. Pfaller, F. C. Tenover, and R. H. Yolken (ed.). Manual of clinical microbiology, 7th ed. American Society for Microbiology, Washington, D.C.
2.Arbeit, R. D., A. Slutsky, T. W. Barber, J. N. Maslow, S. Niemczyk, J. O.
Falkinham III, G. T. O’Connor, and C. F. Von Reyn.1993. Genetic diversity
among strains ofMycobacterium aviumcausing monoclonal and polyclonal bacteremia in patients with AIDS. J. Infect. Dis.167:1384–1390.
3.Barbier-Frebourg, N., D. Nouet, L. Lemee, E. Martin, and J.-F. Lemeland.
1998. Comparison of ATB Staph, Rapid ATB Staph, Vitek, and E-test methods for detection of oxacillin heteroresistance in staphylococci possess-ingmecA. J. Clin. Microbiol.36:52–57.
4.Bono, M., T. Jemmi, C. Bernasconi, D. Burki, A. Telenti, and T. Bodmer.
1995. Genotypic characterization ofMycobacterium aviumstrains recovered from animals and their comparison to human strains. Appl. Environ. Micro-biol.61:371–373.
5.Burki, D. R., C. Bernasconi, T. Bodmer, and A. Telenti.1995. Evaluation of
the relatedness of strains ofMycobacterium aviumusing pulsed-field gel electrophoresis. Eur. J. Clin. Microbiol. Infect. Dis.14:212–217.
6.Carbonne, A., N. Lemaitre, M. Bochet, C. Truffot-Pernot, C. Katlama, J.
Grosset, F. Bricaire, and V. Jarlier.1998.Mycobacterium aviumcomplex
common-source or cross-infection in AIDS patients attending the same day-care facility. Infect. Control Hosp. Epidemiol.19:784–786.
7.Coffin, J. W., C. Condon, C. A. Compston, K. N. Potter, L. R. Lamontagne,
J. Shafiq, and D. Y. Kunimoto.1992. Use of restriction fragment length
polymorphisms resolved by pulsed-field gel electrophoresis for subspecies identification of mycobacteria in theMycobacterium aviumcomplex and for isolation of DNA probes. J. Clin. Microbiol.30:1829–1836.
8.Collins, D. M., S. Cavaignac, and G. W. de Lisle.1997. Use of four DNA
insertion sequences to characterize strains of theMycobacterium avium com-plex isolated from animals. Mol. Cell. Probes11:373–380.
9.Conville, P. S., J. F. Keiser, and F. G. Witebsky.1989. Mycobacteremia
caused by simultaneous infection withMycobacterium aviumand Mycobac-terium intracellularedetected by analysis of a BACTEC 13A bottle with the Gen-Probe kit. Diagn. Microbiol. Infect. Dis.12:217–219.
10. Devallois, A., and N. Rastogi.1997. Computer-assisted analysis of
Mycobac-terium aviumfingerprints using insertion elements IS1245and IS1311in a Caribbean setting. Res. Microbiol.148:703–713.
11. Garzelli, C., N. Lari, B. Nguon, M. Cavallini, M. Pistello, and G. Falcone.
1997. Comparison of three restriction endonucleases in IS1245-based RFLP typing ofMycobacterium avium. J. Med. Microbiol.46:933–939.
12. Graham, L., Jr., N. G. Warren, A. Y. Tsang, and H. P. Dalton.1988.
Myco-bacterium aviumcomplex pseudobacteriuria from a hospital water supply. J. Clin. Microbiol.26:1034–1036.
13. Gubler, J. G. H., M. Salfinger, and A. Von Graevenitz.1992. Pseudoepidemic
of nontuberculous mycobacteria due to a contaminated bronchoscope clean-ing machine. Chest101:1245–1249.
14. Guerrero, C., C. Bernasconi, D. Burki, T. Bodmer, and A. Telenti.1995. A
novel insertion element fromMycobacterium avium, IS1245, is a specific target for analysis of strain relatedness. J. Clin. Microbiol.33:304–307.
15. Hampson, S. J., J. Thompson, M. T. Moss, F. Portaels, E. P. Green, J.
Her-mon-Taylor, and J. J. McFadden.1989. DNA probes demonstrate a single
highly conserved strain ofMycobacterium aviuminfecting AIDS patients. Lanceti:65–69.
16. Horsburgh, C. R., Jr., D. P. Chin, D. M. Yajko, P. C. Hopewell, P. S. Nassos,
E. P. Elkin, W. K. Hadley, E. N. Stone, E. M. Simon, P. Gonzalez, S. Ostroff,
and A. L. Reingold.1994. Environmental risk factors for acquisition of
Mycobacterium aviumcomplex in persons with human immunodeficiency virus infection. J. Infect. Dis.170:362–367.
17. Hunter, P. R., and M. A. Gaston.1988. Numerical index of the
discrimina-tory ability of typing systems: an application of Simpson’s index of diversity. J. Clin. Microbiol.26:2465–2466.
18. Inderlied, C. B., C. A. Kemper, and L. E. M. Bermudez.1993. The
Myco-bacterium aviumcomplex. Clin. Microbiol. Rev.6:266–310.
19. Lari, N., M. Cavallini, L. Rindi, E. Iona, L. Fattorini, and C. Garzelli.1998.
Typing of humanMycobacterium aviumisolates in Italy by IS1245-based restriction fragment length polymorphism analysis. J. Clin. Microbiol.36:
3694–3697.
20. Maslow, J. N., M. E. Mulligan, and R. D. Arbeit.1993. Molecular
epidemi-ology: application of contemporary techniques to the typing for microorgan-isms. Clin. Infect. Dis.17:153–164.
21. Maslow, J. N., A. M. Slutsky, and R. D. Arbeit.1993. Application of
pulsed-field gel electrophoresis to molecular epidemiology, p. 563–572.InD. H. Persing, T. F. Smith, F. C. Tenover, and T. J. White (ed.). Diagnostic molecular microbiology: principles and applications. American Society for Microbiology, Washington, D.C.
22. Matsiota-Bernard, P., S. Waser, P. T. Tassios, A. Kyriakopoulos, and N. J.
Legakis.1997. Rapid discrimination ofMycobacterium aviumstrains from
AIDS patients by randomly amplified polymorphic DNA analysis. J. Clin. Microbiol.35:1585–1588.
23. Mazurek, G. H., D. P. Chin, S. Hartman, V. Reddy, C. R. Horsburgh, Jr.,
T. A. Green, D. M. Yajko, P. C. Hopewell, A. L. Reingold, and J. T. Crawford.
1997. Genetic similarity amongMycobacterium aviumisolates from blood, stool, and sputum of persons with AIDS. J. Infect. Dis.176:976–983.
24. Mazurek, G. H., S. Hartman, Y. Zhang, B. A. Brown, J. S. R. Hector, D.
Murphy, and R. J. Wallace, Jr.1993. Large DNA restriction fragment
polymorphism in theMycobacterium avium-M. intracellularecomplex: a po-tential epidemiologic tool. J. Clin. Microbiol.31:390–394.
25. Mazurek, G. H., V. Reddy, B. J. Marston, W. H. Haas, and J. T. Crawford.
1996. DNA fingerprinting by infrequent-restriction-site amplification. J. Clin. Microbiol.34:2386–2390.
26. McFadden, J. J., Z. M. Kunze, F. Portaels, V. Labrousse, and N. Rastogi.
1992. Epidemiological and genetic markers, virulence factors and intracel-lular growth ofMycobacterium aviumin AIDS. Res. Microbiol.143:423–430.
27. Murray, P. R.1991. Mycobacterial cross-contamination with the modified
Bactec 460 TB system. Diagn. Microbiol. Infect. Dis.14:33–35.
28. Pestel, M., J.-L. Pons, R. Goodman, E. Aronson, J. Maslow, and R. D. Arbeit.
1996. Fifteen year review of the genetic diversity of methicillin-sensitive
Staphylococcus aureusbloodstream isolates at a VA Medical Center, abstr. P297.InProgram and abstracts of the 8th International Symposium on Staphylococci and Staphylococcal Infections.
29. Pestel-Caron, M., and R. D. Arbeit.1998. Characterization of IS1245for
strain typing ofMycobacterium avium. J. Clin. Microbiol.36:1859–1863.
30. Picardeau, M., A. Varnerot, T. Lecompte, F. Brel, T. May, and V. Vincent.
1997. Use of different molecular typing techniques for bacteriological fol-low-up in a clinical trial with AIDS patients withMycobacterium avium
bacteremia. J. Clin. Microbiol.35:2503–2510.
31. Picardeau, M., and V. Vincent.1996. Typing ofMycobacterium aviumisolates
by PCR. J. Clin. Microbiol.34:389–392.
32. Ritacco, V., K. Kremer, T. Van Der Laan, J. E. M. Pijnenburg, P. E. W. de
Hass, and D. Van Soolingen.1998. Use of IS901and IS1245in RFLP typing
of Mycobacterium avium complex: relatedness among serovar reference strains, human and animal isolates. Int. J. Tuberc. Lung Dis.2:242–251.
33. Roiz, M. P., E. Palenque, C. Guerrero, and M. J. Garcia.1995. Use of
restriction fragment length polymorphism as a genetic marker for typing
Mycobacterium aviumstrains. J. Clin. Microbiol.33:1389–1391.
34. Ross, B. C., and B. Dwyer.1993. Rapid, simple method for typing isolates of
Mycobacterium tuberculosisby using the polymerase chain reaction. J. Clin. Microbiol.31:329–334.
35. Schlichting, C., C. Branger, J.-M. Fournier, W. Witte, A. Boutonnier, C.
Wolz, P. Goullet, and G. Doring.1993. Typing ofStaphylococcus aureusby
pulsed-field gel electrophoresis, zymotyping, capsular typing, and phage typ-ing: resolution of clonal relationships. J. Clin. Microbiol.31:277–232.
36. Slutsky, A. M., R. D. Arbeit, T. W. Barber, J. Rich, C. F. Von Reyn, W.
Pieciak, M. A. Barlow, and J. N. Maslow.1994. Polyclonal infections due to
Mycobacterium aviumcomplex in patients with AIDS detected by pulsed-field gel electrophoresis of sequential clinical isolates. J. Clin. Microbiol.
32:1773–1778.
37. Telenti, A., F. Marchesi, M. Balz, F. Bally, E. C. Bottger, and T. Bodmer.
1993. Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis. J. Clin. Microbiol.31:175– 178.
38. Tenover, F. C., R. D. Arbeit, and R. V. Goering.1997. How to select and
interpret molecular strain typing methods for epidemiological studies of bacterial infections: a review for healthcare epidemiologists. Infect. Control Hosp. Epidemiol.18:426–439.
39. Tenover, F. C., R. D. Arbeit, R. V. Goering, P. A. Mickelsen, B. E. Murray,
D. H. Persing, and B. Swaminathan.1995. Interpreting chromosomal DNA
restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol.33:2233–2239.
40. Tokars, J. I., M. M. McNeil, O. C. Tablan, K. Chapin-Robertson, J. E.
Patterson, S. C. Edberg, and W. R. Jarvis.1990.Mycobacterium gordonae
pseudoinfection associated with a contaminated antimicrobial solution. J. Clin. Microbiol.28:2765–2769.
41. Tsang, A. Y., J. C. Denner, P. J. Brennan, and J. K. McClatchy.1992.
on May 15, 2020 by guest
http://jcm.asm.org/
Clinical and epidemiological importance of typing ofMycobacterium avium
complex isolates. J. Clin. Microbiol.30:479–484.
42.Vannier, A. M., J. J. Tarrand, and P. R. Murray.1988. Mycobacterial cross
contamination during radiometric culturing. J. Clin. Microbiol.26:1867– 1868.
43. Von Reyn, C. F., J. N. Maslow, T. W. Barber, J. O. Falkinham III, and R. D.
Arbeit.1994. Persistent colonisation of potable water as a source of
Myco-bacterium aviuminfection in AIDS. Lancet343:1137–1141.
44. Yajko, D. M., D. P. Chin, P. C. Gonzalez, P. S. Nassos, P. C. Hopewell, A. L.
Reingold, C. R. Horsburgh, Jr., M. A. Yakrus, S. M. Ostroff, and W. K.
Hadley.1995.Mycobacterium aviumcomplex in water, food, and soil samples
collected from the environment of HIV-infected individuals. J. Acquired Immune Defic. Syndr. Hum. Retrovirol.9:176–182.
45. Yakrus, M. A., M. W. Reeves, and S. B. Hunter.1992. Characterization of
isolates ofMycobacterium aviumserotypes 4 and 8 from patients with AIDS by multilocus enzyme electrophoresis. J. Clin. Microbiol.30:1474–1478.
![FIG. 1. Restriction patterns from Aseby PFGE. Lanes: 1 and 6, bacteriophage lambda DNA concatemers (sizes [inkilobases] are indicated on the left); 2, isolate 100A8; 3 and 4, pattern P7(isolates 100A28 and 100A32, respectively); 5, isolate 100A25; 7 to 11,](https://thumb-us.123doks.com/thumbv2/123dok_us/8229585.825775/2.612.320.548.326.698/restriction-patterns-bacteriophage-concatemers-inkilobases-indicated-isolates-respectively.webp)