J. Exp. Biol. (1966), 45, 449-464 4 4 9 With 10 text-figures
Printed in Great Britain
OLFACTION AND VISION IN FISH SCHOOLING
BY C. C. HEMMINGS*
Department of Zoology, University of Cambridge, and Stazione Zoologica, Naples
(Received 17 June 1966)
INTRODUCTION
A fish school must be regarded as a group of individuals in dynamic equilibrium, with the sensory input of each fish providing a feedback of information affecting the maintenance and behaviour of the school.
Many previous studies of schooling have concentrated upon analysis of the reactions of individual fish to stimuli involved in the formation and maintenance of school structure, as distinct from the group behaviour of the whole school. The important sensory modalities involved in both school structure and group behaviour are vision, the acoustico-lateralis sense and olfaction, which all give fish information about stimuli at some distance from their source.
Most work on the sensory basis of schooling indicates that vision is the most im-portant modality (Spooner, 1931; Shlaifer, 1942; Morrow, 1948; Keenleyside, 1955; Breder, 1959; Shaw, 1962; and many others). It does not appear from any of this work that either shape, colour, movement or any other particular aspect of the visual stimulus is specific for any species. Moulton (i960) suggested that sounds produced by swim-ming fish are stimuli used in the maintenance of school structure. Harris & van Bergeijk (1962) conclude from research on the lateral line that it could be used for the main-tenance of school structure, and this view is supported by Dijkgraaf (1963).
Less attention has been paid to the possible role in schooling of the chemical senses, although the repellent odour of' Schreckstoff is well known (von Frisch, 1938, 1941 a,
b; review by Pfeiffer, 1962). Olfaction is to be functionally distinguished from gustation
and the common chemical sense in fish largely on grounds of its higher sensitivity, allowing perception of odours at some distance from their source. Wrede (1932) was the first to show a positive response in untrained fish to a species-specific odour, using minnows in a two-compartment choice situation. Goz (1941) followed up this work by a further study of minnows using a training technique. He showed that not only can minnows distinguish between a variety offish and amphibian species, but even between individuals of their own species. Keenleyside (1955) showed that temporarily blinded untrained rudd preferred that end of a tank where rudd odour was present.
In the present study the behaviour of roach in an odour gradient, and of roach, grey mullet and Chromis chromis, & pomacentrid species, to visual stimuli involved in schooling were investigated. Incidental observations were made on other aspects of schooling behaviour related to the main problem of contact between individuals. Nearly all workers on schooling have coined their own definitions of 'school' or
• Present address: Marine Laboratory, Aberdeen.
'shoal', but that of Keenleyside (1955) is followed in the present study. A school is defined by him as ' an aggregation formed when one fish reacts to one or more fish by staying near them'. Excluded from such aggregations are cases where the groups so formed are clearly associated for fighting or courtship. It follows that two fish can constitute a school provided that they are aware of each other. The degree of parallel orientation is not thought to be important.
MATERIAL
Roach, Rutihis rutilus L. (Cyprinidae), of 9-12 cm. length, were caught in Coe Fen Brook in Cambridge. Those kept socially were put in large holding aquaria filled with tap water in the same room as the experimental apparatus. Some fish were kept in isolation in conditions described in the relevant section. The fish were fed on ' Bemax' with occasional chopped earthworm and Tubifex, but the fish also browsed occasionally on the algal film inside the aquaria. All aquaria were permanently aerated with a jet of tap water. Kept in this way the fish appeared healthy and none died during the experi-mental period.
Chromis ckromis L. (Pomacentridae), of 8-10 cm. length, and grey mullet, Mugil chelo Cuvier (Mugilidae), of 10-14 cm- length, were caught in the Bay of Naples by
fishermen from the Stazione Zoologica, and kept separately after capture in large holding aquaria. Both species were fed on macerated fresh sardine flesh. Aeration was provided by a jet of sea water from the laboratory circulation. Initial mortality was restricted to those with fins damaged or scales removed during capture, the remainder stayed healthy for the duration of the experimental work.
METHODS
(a) Apparatus
The behaviour of Rutilus was studied at Cambridge using a horizontal linear gradient tank similar to that devised by Shelford & Allee (1913), and subsequently used by Shelford & Powers (1915), and Bowen (1931). The apparatus used is shown diagram-matically in Fig. 1. Mains tap water at a controlled rate of 2 l./min. was equally divided into the two inlet tanks, and water from these then entered the experimental tank through perforated glass pipes in each corner. Flow out of the experimental tank from the middle was via a large siphon made of Alkathene (black polyethylene) expanded into a funnel made of Perspex (clear acrylic plastic) filling the whole width of the ex-perimental tank. All other piping was of glass and Polythene (polyethylene).
By equalizing the rates of mains inflow and the final outflow from the apparatus the water levels were kept constant. The experimental tank was much larger than those used by the above-mentioned authors, being 180x28 cm. with water to a depth of 22 cm. This fact, plus the relatively slow flow rate, reduced the possibility of the fish receiving rheotactic cues due to water movement through the tank.
01]-action and vision in fish schooling 451
tank in order to prevent the fish seeing its own reflexion. The experimental tank and the mirror were surrounded by a black screen forming part of a light-tight canopy extending in front of the apparatus. The observer sat at a table outside the canopy and observed the fish through a small peephole.Inlet siphon Outlet siphon Inlet siphon
Front elevation
Jtl
Mains supplyInlet tank
Screen Outlet siphon
Inlet tank
Inlet siphons ,Perspex funnel
Plan
-Mirror
Inlet tank
End section
Outlet *. tank
-Mirror
Outlet siphon
t 1
1
Perspex "funnel
Centre section
Fig. 1. Experimental apparatus used for study of reactions of Rutilus to olfactory and visual stimuli.
The experimental tank used in Naples for the study of Mugil and Chromis was 100 x 30 cm. with water to a depth of 20 cm. The bottom of the tank was marked with a rectangular grid of 16-6 x 15 cm. rectangles. Observation was again by a mirror at 450 above the tank, the observer being seated behind a screen with a peephole. Illumination of the tank was by north daylight.
(b) Procedure
assumed that the water mass was still. All visual experiments were commenced at least 15 min. after filling when slight water movements still occurred. These were not considered to be significant.
(i) Olfactory and control experiments. At the start of olfactory experiments using attractant odour four Rutilus were transfered to one of the inlet tanks with no flow through the system. After 1 hr. flow of water was started, and after a further 5 min. to establish an odour gradient in the experimental tank, a single fish caught at random in the holding aquarium was transferred to the odour end of the experimental tank, i.e. the end with the inlet from the tank containing four Rutilus. Observation was begun within a few seconds. No attempt was made to analyse the odour which it was assumed the fish secreted or excreted at a constant rate. The technique was standardized to result in a concentration of four ' fish-hour units' of odour in the inlet tank at the start of an experiment. Control experiments differed only in that tap water flowed from each end, with no fish in either inlet tank.
A short series of negative olfactory experiments involving the ' Schreckreaktion' was carried out. The skin from one flank of a freshly killed Rutilus (an area of 8-10 cm.2) was ground in a mortar. The resulting paste was shaken up with water and filtered. Half the filtrate was added to one of the inlet tanks and the other half was set to run slowly out of a burette (into the same inlet tank) during the course of an experiment. Fish were transfered between the holding aquaria and the experimental tank in a special net comprising a short tube of soft Nylon netting terminating in a cornerless Polythene bag. The fish were thus contained in 200-400 c.c. of water after the method of Verheijen (1953). The use of a normal net in which the fish flapped about when being transfered, was shown to cause a ' Schreckreaktion' on some occasions. This was also reported by Kiilzer (1954) in work on tadpoles.
(ii) Visual experiments. Two sources of visual stimulus were used in experiments on
Rutilus, Mugil and Ckromis, either ' mirror' or ' caged fish'. In the former, the ground
glass screen at one end of the experimental tank was replaced by a mirror, and in the latter a watertight transparent box made of Perspex and measuring 28 x 15 cm. was placed at one end of the experimental tank. This box contained a single fish.
RECORDING
Olfaction and vision in fish schooling 453
of that predicted were its behaviour entirely random. The response strength is thus defined numerically as the sum of the deviations from random of the frequencies of recordings in the three regions of the tank nearest the stimulus.
Records of the position of the fish, in a numbered succession at fixed intervals of space and time, allowed the calculation of speed and activity. Activity is defined as the total recorded distance -r- total recorded time, i.e. cruising speed including stops. The, calculation of speed and activity is more fully discussed in another paper (Hemmings, 1966).
EXPERIMENTS WITH SOCIALLY LIVING FISH
(a) Results of olfactory experiments on Rutilus
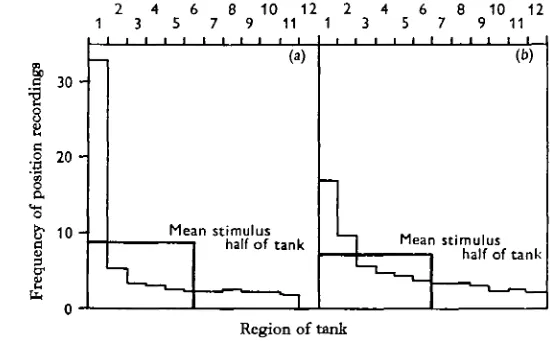
A total of forty experiments on the responses to Rutilus attractant odour was per-formed, and the mean result of these is given in Fig. 2 a. There was a clear preference on the part of individual fish for that end of the tank where Rutilus odour was present.
- 2 5
- ' 2 0
- 15
- 10
Region of tank
Fig. 2. Results from olfactory experiments using Rutilus. (a) Attractant odour;
(b) repellent Schreckstoff.
Also shown in Fig. zb is the mean result from seven experiments in which the repellent ' Schreckstoff' was used as the olfactory stimulus. Comparison of the two histograms shows that the positive reaction to the odour from undamaged fish was rather less strong than the ' Schreckreaktion' or negative response to the odour of damaged fish. This difference occurred because all the fish when tested showed a strong response to the ' Schreckstoff', but some fish showed very little response to the attractant odour. It might be expected that the 'Schreckreaktion' would be a more fixed behaviour pattern than the attraction to species odour. In the case of the attractant odour experi-ments, an inverse correlation occurred between the response strength and the activity of the fish, that is, those showing a lower preference for the odour were more active. A Spearman rank correlation coefficient (see Siegel, 1956) gave rs = 0-53. Although
this is not a high degree of correlation, it is highly significant, P < 0-0005.
454
of the gradient. If the result of the positive olfactory experiments could in some degree be accounted for by a tendency for fish to prefer the end into which they were first placed, then the results from control experiments arranged with the ' introduction end' to the left should indicate this tendency. A x2 test on the data from fifty control experi-ments gave a probability of being random of P = 0-95.
(i) Changes in reaction to odour with time. Two classes of decline in response strength occurred: first, the proportion of time spent at the stimulus end of the odour gradient decreased during the experiment; and secondly, the longer the fish had been in
Region of tank 1 2 3 4 5 6 7 8 9 10 1112
I I I I I I I I I I I
S 10 15 Minutes of experiment
Fig. 3 Fig. 4
Fig. 3. Short-term change in response strength and activity during experiments. • • , Response strength in repellent experiments; • • , response strength in attractant experi-ments ; O O, activity in attractant experiexperi-ments.
Fig. 4. Long-term decline in strength of response to attractant odour, (a) Testing begun after 3 days in captivity; (b) testing begun after 74 days in captivity; (c) testing begun after 182 days in captivity.
captivity in the holding tank the lower was their response when tested. These two changes are referred to below as the short-term and long-term declines in response strength.
Olfaction and vision in fish schooling 455
the experiment, the response remained greatest at the odour end of the gradient. The 'Schreckreaktion' also shown in Fig. 3 does not show the same decline in response strength, but increases during the first three recording periods. A x2 test on the frequency data gave a probability of being from the same population of P = o-oi. This suggests that the slow rise and sharp fall are significant. It may be that the break in response strength of both attractant and repellent at about the same time, is associated with a change in the equilibrium of the odour gradient. The curve for activity of fish during the positive olfactory experiments in Fig. 3 clearly shows the inverse relation between response strength and activity.
The long-term decline is illustrated in Fig. 4, showing the mean result from three series of ten experiments performed after the fish had been different periods in holding aquaria. A x% test on the original frequency data gave a probability of their being from the same population of P <^ o-ooi. A fourth series of experiments using freshly caught fish on the third day after capture failed to confirm the expected high response strength, but this was probably due to the high level of sexual activity shown by these fish in the holding tank. There is no evidence that olfactory stimuli play any part in the courtship behaviour of Rutilus. The 'Schreckreaktion' also shows a similar decline in response strength (Pfeiffer, 1962). These results may be evidence for a general lowering of responsiveness of fish kept in captivity.
(ii) Olfactory control experiments. Learning of odours plays some part in the social behaviour of fish (Kuhme, 1962), and latent learning of possibly significant odours could occur in the holding aquaria. To investigate this possibility a series often control experiments was carried out using recently caught fish as experimental animals. The odour was derived for half the experiments from fish that had been in the same holding aquarium as the experimental fish for the 3 days after capture, and half from fish that had been in a different holding aquarium for over a year. The results showed no suggestion of a difference in response, a x* test giving a value of P = 0-98.
The only inter-specific controls comprised a series of ten experiments using small perch, Perca fluviatilis in the inlet tank. The result of these experiments showed no significant difference from random, a x2 test giving a value of P = 0-85. This result indicates either that there is no response by Rutilus to odour from Perca, or alternatively that Perca does not normally produce any odorous substance.
(b) Results of visual experiments
the two classes of experiment with mean frequencies grouped into stimulus and non-stimulus halves of the experimental tank:
Stimulus Non-stimulus
Olfactory
Visual 4447-95
16 \ 12-05/
A X* t681 o n original data gave P = 0-05
It is thus clear that in the conditions of the experiment Rutilus odour and the visual image of Rutilus were both effective attractants to individual fish.
The visual results using mirror and caged fish as stimuli are given separately in Fig. 6 for Rutilus, Mugil and Chromis, together with the mean result of control experi-ments for the same species. A ;\;2 test was used to compare adjacent results, i.e. caged fish with mirror and mirror with control. The results of these tests are given as the values of P in the figure. Rutilus and Mugil showed strong reactions towards both
2 4 6 8 1 0 1 2 2 4 6 8 10 12
1 3 5 7 9 11 1 3 5 7 9 11
I 1 1 1 1 I 1 I 1 r 1 I r 1 1 1 1 I 1 1 1 1 1
8
I
20fr 10
-a
(a)
Mean stimulus half of tank
(b)
Mean stimulus half of tank
Region of tank
Fig. 5. Response of Rutihu to: (a) visual stimulus (directional), and
(b) olfactory stimulus (non-directional).
types of visual stimulus, but in both species the result from the mirror experiments was less marked than that from the caged-fish experiments. This was probably due to the optical difference of the stimulus situations; an experimental fish at the opposite end of the tank sees the caged fish through a water mass equal to the length of the tank, but its virtual image 'behind the mirror' through a water mass equal to twice that length. Thus not only the clarity of the image but the angular size at the eye will differ.
Chromis showed quite a strong reaction to the caged fish but its response to the mirror
did not significantly differ from the control case. However, underwater observations in the field show that the response of Chromis to mirrors is then rather greater. The change in response strength during visual experiments is shown in Fig. 7. It is clear that the marked decline in response that occurs with the olfactory stimulus does not occur in this case. Mugil does in fact show a slight rise in response strength during the experimental period.
[image:8.451.80.355.249.421.2]Olf action and vision in fish schooling 457
and forwards along the tank, but not showing any marked preference for the ends.Mugil and Ckromis spent much time swimming at the end walls of the experimental
tank, as if in an effort to escape. Ckromis often spent some time at the beginning of experiments backed tail first in a corner, a typical territorial defense attitude for this species.
§
•a
Caged fish Mirror Control
2 4 6 8 1 0 1 2 2 4 6 8 1 0 1 2 2 4 6 8 1 0 1 2
1 3 5 7 9 1 1 1 3 5 7 9 1 1 1 3 5 7 9 1 1
4 0 •
30
20
10
-0
40 •
30
-O- 20
c 10 H
I
-4 0 •
30
20
10
-I -I -I -I -I -I -I -I -I -I -I Caged fish/mirror
P=003
(a)
Caged fish/ mirror P<0 001
(b)
Caged fish/ mirror P-C0 001
I I I I I I I Mirror/control
Mirror/control
Mirror/control
P = 0 25
I I I I I I I I I I I
I I I I I I I I I I I I I I I I I 1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
Region of tank
Fig. 6. Results of visual experiments using caged-fish and mirror stimuli compared with control results: (a) Eutilus, (b) Mugil, (c) Chromit.
two fish swimming free were never more than 75 cm. apart in 2400 recordings (ten experiments), whereas the caged-fish result in Fig. 6 a shows eighty-four recordings out of the same total when the experimental fish was more than 75 cm. from the caged fish. Two individuals of Mugil and Ckromis were observed to behave in very much the same way, that is, they stayed very much closer than the results from visual experiments with mirror and caged fish would suggest.
e
3 0"0
c
B 2 5
-S
§ J 20
-a 5
2 £ l | i S
-%%
strer
i
o
f e
r
0
2
I
s
"
0 -*
P /
I 1 I 1
•
1 1 1 0 5 10 15 20
Minutes of experiment
Fig. 7. Short-term change of response strength during visual experiments. • , Rutilus; O, Mugil; x, Ckromis.
50
40
30
-§
2010
-(3) Social (b) Isolate
\ I I I 1
0 15 30 45 60 75 0 15 30 45 60 75 90 105 120135150165 cm. Separation between fish
Fig. 8. Distance of separation between two fish: (a) both fish living socially and
(b) one fish living socially and the second living in isolation.
Olf action and vision in fish schooling 459
patterns which it is suggested are of basic significance in the cohesion of a fish school. The three motor patterns are as follows:
(1) Exploratory tendency—either fish would initiate swimming in any direction away from the other.
(2) Returning tendency—if the fish swimming away was not followed, it would turn and swim back to the other.
(3) Following tendency—usually when one fish swam away it would be followed by the second fish.
The first motor pattern appeared to be quite spontaneous and is referred to as an 'action' in this context, although it is probably caused by some unknown internal or external stimulus. The other two motor patterns clearly depended upon the perception by either fish of the increase in separation brought about by the original action, and are therefore termed 'reactions'. The dynamic relationship between two individuals seems entirely explicable by these three motor patterns. Such interspecific differences as occur can be explained by differences in the relative strength of the three com-ponents, and the delay between the occurrence of the exploratory action and one of the reactions.
EXPERIMENTS WITH R UTIL US KEPT IN ISOLATION
A study was made of the changes in reaction to visual and olfactory stimuli of four
Rutilus kept in isolation instead of socially in a holding aquarium. A number of the
resulting changes in behaviour give indications of possible sensory mechanisms that operate in typical schooling behaviour.
(i) Conditions of isolation. Two fish (Ai and A 2) were isolated after 214 days living socially in a holding aquarium and two (B1 and B 2) were isolated immediately after capture. Each isolation aquarium measured 28 x 28 cm. with water to a depth of 32 cm. Conditions of feeding and aeration were exactly as for the social holding aquaria. The isolates thus experienced the following deprivations:
(1) Visual contact with other fish. (The tanks were screened from each other with black paper and after a few days the growth of an algal film prevented the fish seeing their own reflexions.)
(2) Olfactory stimuli from other fish.
(3) Sounds, water movements and pressure changes associated with the presence of other fish.
In isolation all four fish were much less active than those living socially. Evidence for this comes not only from direct observation but also from the distribution of faecal pellets which collect in the corners of an aquarium when fish are active. The pellets in the four isolate aquaria were always scattered over the whole area of the bottom.
(ii) Sequence of experiments. The same four fish were used for two groups of experi-ments : the first group comprised three trials per fish of each experiment type, control, visual and olfactory arranged in a randomized order; the second group comprised one trial per fish of control, olfactory and visual type, in that order. The fish had been in isolation for 171 days prior to the first group of experiments, and an interval of 132 days elapsed before the second group was started. An interval of 61 days then elapsed before a two-fish separation experiment and return to social life.
460 C. C. HEMMINGS
giving P = 0-99. One interesting point was that all fish were much more active in the experimental tank than in their isolate aquaria.
Shown in Fig. 9 are the curves for the short-term changes in response strength of the two isolate series of olfactory experiments and their mean, and the mean social result for comparison. The results indicated that the total response of isolates was greater than the mean result for fish living socially, and that this was due to a less
Pi
u
I I I I [
0 5 10 15 Minutes of experiment
Fig. 9. Change in strength of response to attractant odour during experiments. • • , First series of twelve isolate experiments; • • , second series of foui isolate experiments; 0 * , mean of first and second isolate series; O O, mean of thirty social experiments.
marked short-term decline in response strength. There was no significant difference between the results of the two A fish that had been a long period in captivity and the two B fish that were isolated immediately on capture. The long-term decline in response strength thus occurs in the isolates to a much lesser extent than in the socially living fish. The results of the second series of isolate experiments defy logical explanation but the two series together show that experiments must be conducted with larger numbers of isolate fish and with more intermediate testing.
The visual reaction of isolate fish was tested using only the caged-fish stimulus. The normally clear-cut visual response was completely upset by periods of isolation, so that the mean frequency distribution of position recordings would give no real in-formation about the behaviour of the fish. The results could be simply categorized as follows: (1) positive reaction ( + )—attraction towards caged fish; (2) neutral reaction (N)—apparent indifference to the other fish; (3) negative reaction (-)—avoidance of the vicinity of the other fish. The following table shows the results for each fish from each visual experiment, with the time interval in days between successive experiments:
Interval between
isolation 1st and 3rd 4th and testing experiment Interval experiment Interval experiment Interval experiment
I ? s N 12 N N N + 7 +N + + 140 + + + +
174 N N + + 11 9 J4° N - + +
Fish
Ai A2 Bi
B2 173
N N
12 11
17
12
N 140
140
Olfaction and vision in fish schooling 461
The four symbols shown for each experiment represent four 5 min. periods into which each experiment was divided. Although the normally high response to visual stimuli was completely altered by isolation, there is some evidence for a return to the positive reaction occurring either within or between experiments.
After an interval of 61 days a short 'two-fish separation' experiment was carried out on three of the isolate fish which were then returned to the social holding aquarium. The experimental technique was exactly as for the socially living fish but here using one isolate fish and one social fish, and recording for only 5 min. The mean result is shown in Fig. 8b. The reason for the rather greater separation shown in this series of experiments can be described by using the 'action and reaction' system of motor patterns described above. On first being placed in the experimental tank both fish were very active, showing little response to each other, but quite quickly the social fish began to follow the isolate. This behaviour persisted until some time later when the isolate fish showed returning and following reactions also; the returning reaction of the isolate fish was the last motor pattern to appear.
At the conclusion of each experiment the two fish were returned to a holding aquarium. After about 5 min., when co-ordinated swimming with the school was poor, the isolates became indistinguishable from other members of the school.
DISCUSSION AND CONCLUSIONS
The results from these experiments agree with those from previous work on schooling behaviour that vision is the sense primarily involved. Little attempt has been made to analyse or even describe school cohesion with the exception of the work of Breder (1959), of Horstmann (i960) and of Shaw & Tucker (1965) who investigated the opto-motor responses of schooling fish. It is suggested that the system of action and reaction tendencies described above is of value in describing school cohesion, and may even be of underlying significance. The graph in Fig. 10 shows non-quantitatively how the tendencies are related in the simple but representative case of a two-fish school. All schooling fish seem to show a strong exploratory tendency which is normally maximal when they are at the optimum separation. If fish ' 1' swims away from fish ' 2 ' , the most likely occurrence is that fish ' 2' follows it (i?2), because the following tendency is
observed to be greater than the returning tendency. Alternatively the fish ' 1' may stop exploring and show a returning tendency (i?x). The observations by Keenleyside (1955)
and Steven (1959) could very easily be described by this system which might help in relating the optomotor responses to school cohesion. Many hours of underwater observation in the Mediterranean and in British waters suggests that this system of tendencies does have real significance. Characteristic changes in school shape, inter-individual distance and orientation associated with, for example, the onset and cessa-tion of feeding in a wide variety of species can be explained by changes in the relative strength of the different tendencies.
462 C. C. HEMMINGS
plane surface or try to nose their way through, but if one slides the glass sheet away they take up a normal inter-individual distance. It seems unlikely that there is any optical information which does not pass through the glass sheet, suggesting that the repulsive force could be the water movements caused by the swimming of each fish, perceived by the lateral-line sense of the other. The work of Harris & van Bergeijk (1962) shows the lateral line to be sensitive to near-field displacements at a distance not greater than would fit the requirement of a repulsive force.
R1+R2
S1 S2 Separation between two fish
Fig. 10. Proposed basis of visual contact between schooling fish. Ai (action), exploratory tendency of first fish; Ri (reaction), returning tendency of first fish; R2 (reaction), following tendency of second fish; So, optimum or preferred separation between fish; Si, separation when^4i = R 1 +.R2, with two fish swimming freely ; 5 z , separation when^4i = i ? i , w i t h o n e fish free and the other caged but visible.
Olfaction and vision in fish schooling 463
passive drift of fish in a water mass under these conditions does occur in the open sea (Jones, 1962). It is useful to consider the curves in Fig. 10 extended beyond the limit of visual contact between fish. The following tendency depends upon intervisibility and must fall to zero, but the exploratory and returning tendencies may remain constant as a pattern of activity and turning that are a part of the mechanism of orientation to odour gradients described in another paper (Hemmings, 1966). It was shown that the response strength in an odour-gradient situation was inversely related to activity. The experimental method was not designed to show the cause and effect relationship in this correlation, but two alternatives are possible: either the fish be-comes less active when it senses an odour of a particular concentration, or alternatively when some other factor lowers activity a greater response is then shown to odours. If the latter were the case in those species that show a diurnal activity cycle with lower activity at night, then it might be expected that the response to odours increases and so the school does not break up. The role of the lateral-line sense in conditions of low visibility would be an obstacle-avoidance sense preventing fish bumping into each other. This is still in effect a 'repulsive force' and quite within the known limits of the lateral-line sensitivity. Thus the change-over from vision plus lateral-line by day to olfaction plus lateral-line at night does not require any great change in the role of the lateral-line sense.
SUMMARY
1. The behaviour of individual roach was observed in a horizontal linear gradient of species odour. No attempt was made to analyse the odour involved.
2. The behaviour of roach, grey mullet and Chromis was studied in the presence of (a) a mirror, or (b) a second fish behind a clear plastic sheet.
3. Roach showed nearly as strong a response to attractant species odour as to the repellent 'SchreckstofF. The relative strengths of responses to directional visual images and non-directional odour gradients were compared.
4. Short-term and long-term declines in the strength of the response to odour occurred.
5. Observation of a 'two-fish school' suggested that a system of exploratory, returning and following tendencies co-ordinated schooling behaviour.
6. Isolation of roach for a period of 20-25 weeks caused enhancement of the olfactory reaction but completely disrupted the visual response.
7. It is suggested that school structure is maintained by balanced attractive and repulsive ' forces'; the attraction modalities involved are vision by day and olfaction by night, and the repulsion modality is the lateral line sense.
Financial support for this work which forms part of a Ph.D. thesis submitted to the University of Cambridge, was received from the Development Commission in the form of a Fishery Research Training Grant, and from the Royal Society and the Philosophical Society of Cambridge for table space at Naples. I am indebted to Prof. C. F. A. Pantin for accommodating me in the Department of Zoology and to Dr P. Dohrn and the staff of the Stazione Zoologica for their assistance. I would like to thank Dr H. W. Lissmann for his help and supervision during the work, and Dr Liss-mann, Dr F. R. Harden Jones and Mr B. B. Parrish for suggestions at the manuscript stage.
REFERENCES
BOWEN, E. S. (1931). The role of the sense organs in the aggregation of Ameiurus melas. Ecol. Monogr. I.
I-35-BRBDER, C. M. (1959). Studies on social groupings in fish. Bull. Am. Mus. not. Hist. 117, 397-481. CROOK, J. H. (1961). The basis of flock organisation in birds, Current Problems in Animal Behaviour.
Ed. W. H. Thorpe and O. L. Zangwill. Cambridge University Press.
DIJKGRAAF, S. (1963). The functioning and significance of the lateral-line organs. Biol. Rev. 38, 51-105. EMLEN, J. T. J. (1952). Flocking behaviour in birds. Auk 69, 160—70.
FRISCH, K. VON (1938). Zur Physiologie des Fischschwarmes. Naturwissenschaften 36, 601-6.
FRISCH, K. VON (1941a). Die bedeutung des Gerunchsinnes im Leben der Fische. Naturwissenschaften 29,
321-33-FRISCH, K. VON (1941J). Ober einen Schreckstoff der Fischhaut und seine biologische Bedeutung.
Z. vergl. Physiol. 39, 46-145.
Goz, H. (1941). Ober den Art—und Individual Geruch bei Fischen. Z. vergl. Pkysiol. 39, 1—45. HARRIS, G. C. & BERGIJK, W. A. VAN (1962). Evidence that the lateral-line organ responds to near-field
displacements of sound sources in water. J. acoust. Soc. Am. 34, 1831-41.
HEMMINGS, C. C. (1966). The mechanism of orientation of roach, Rutilus rutilus L. in an odour gradient.
J. Exp. Biol. 45, 465-74.
HORSTMANN, E. (i 960). Schwarmstudien unter Ausnutzung einer optomotorischen Reaktion bei Mugil
cephalut. Pubbl. Stax. xool. Napoli 31, 25-35.
JONES, F. R. H. (1962). Further observations on the movement of herring {Clupea harengus) in relation to the tidal current. J. Cons. perm. int. Explor. Mer 37, 52-76.
KEENLEYSIDE, M. H. S. (1955). Some aspects in the schooling behaviour of fish. Behaviour 8, 183-248. KCHME, W. VON (1962). Das Schwarmverhalten elterngefuhrter Jungcichliden (Pisces). Z. Tierpsychol. 19,
5!3-538.
KCLZER, E. (1954). Untersuchungen liber die Schreckreation der Erdkrotkaulquappen (Bufo bufo).
Z. vergl. Physiol. 36, 443-63.
MORROW, J. E. (1948). Schooling behaviour in fishes. Q. Rev. Biol. 33, 27-38.
MOULTON, J. M. (i960). Swimming sounds and the schooling of fishes. Biol. Bull. Mar. Biol. Lab.,
Woods Hole 119, 210-23.
PARR, A. E. (1927). A contribution to the theoretical analysis of the schooling behaviour offish. Oec. Pap.
Bingham Oceanogr. Colin, no. 1, 32 pp.
PFEIFFER, W. (1962). The fright reactions offish. Biol. Rev. 37, 495-512. SHAW, E. (1962). The schooling of fishes. Scient. Am. (June 1962), 10 pp.
SHAW, E. & TUCKER, A. (1965). The optomotor reaction of schooling carangid fish. Anim. Behav. 13, 330-336.
SHELFORD, V. E. & AT.T.KB, W. C. (1913). Reactions of fish to gradients of dissolved atmospheric gases.
J. Exp. Biol. 14, 207-26.
SHELFORD, V. E. & Powers, E. B. (1915). An experimental study of the movement of herring and other marine fishes. Biol. Bull. Mar. Biol. Lab., Woods Hole 28, 315-34.
SHLAIFER, A. (1942). The schooling behaviour of mackerel, a preliminary experimental study. Zoologica,
N. Y. 27, 75-80.
SEEGEL, S. (1956). Non-parametric Statistics for the Behavioural Sciences. New York: McGraw-Hill. SPOONER, G. M. (1931). Some observations in schooling in fish. J. Mar. biol. Ass. U.K. 17, 431-48. STEVEN, D. M. (1959). Studies on the shoaling behaviour of fish. I. Responses of two species to changes
of illumination and to olfactory stimuli. J. Exp. Biol. 36, 261—80.