Wrinted in Great Britain
EVIDENCE FOR A SYNAPTICALLY MEDIATED DECREASE
IN CONDUCTANCE IN A CRUSTACEAN MYOCARDIUM
BY J. C. DELALEU
Laboratoire d'Electrophysiologie, University Claude Bernard 69621 Villeurbcmne, France
{Received 22 January 1976)
SUMMARY
1. In the neurogenic heart of the isopod Porcellio dilatatus, electrical stimulation of the cardio-regulatory nerves at rates greater than 20-25 pulses/s elicited inhibitory junctional potentials (IJPs) in the myocardium. It9 cessation was followed by a long lasting hyperpolarization of the myocar-dial membrane (post-stimulus hyperpolarization = PSH).
2. During the PSH the membrane resistance of the heart muscle increased. The PSH was enhanced by myocardium hyperpolarization, decreased by depolarization and reversed around — 50 mV.
3. Picrotoxin inhibited the summated IJP9 elicited by the stimulation and thus caused the membrane to maximally hyperpolarize during the inhibitory train, thus suggesting a composite nature of the inhibitory processes.
4. The PSH was reversibly reduced in K+-free saline or in ouabain con-taining saline but partial restoration was obtained by injection of inward cur-rent to the myocardium.
5. The PSH was abolished in lithium saline and reduced in Na+-deficient (choline) solution. Cl~-deficient solution that markedly affected the summated IJPs shortly after its introduction did not affect the PSH.
6. It is proposed that the PSH results from a decrease in conductance, presumably to both Na+ and K+. The implication of such a mechanism as a component of the inhibitory regulation of this crustacean heart is discussed.
INTRODUCTION
In a preceding paper, the existence of peripheral inhibition was demonstrated in the neurogenic heart of the terrestrial isopod Porcellio dilatatus (Delaleu & Holley, 1976).
It was also found that following suitable repetitive electrical stimulations of the cardio-inhibitory fibres, the myocardial membrane hyperpolarized.
1973) or for both Na+ and K+ (snail neurone; Gerschenfeld & Paupardin-Tritsch
1974)-The present study is an investigation of the mechanisms responsible for the post-stimulation hyperpolarization (PSH) observed in the heart of Porcellio, employing treatments known to alter passive conductances or active transport mechanisms. Preliminary results have been reported elsewhere (Delaleu, 1974; Delaleu & Holley,
METHODS
Methods were generally similar to those described earlier (Holley & Delaleu, 1972; Delaleu & Holley, 1976). The heart tube was isolated but remained connected to the 'aorta' attached to the anterior tergites. The cardio-regulatory nerves, which run along the dorsal wall of the aorta, were stimulated by means of a pair of Ag-AgCl wires embedded in polyethylene tubes and connected to a stimulator through an isolation unit. Spontaneous electrical activity of the heart was recorded with glass micro-electrodes filled with 3M-KCI or o-6 M-K2SO4. A second microelectrode inserted
close to the recording electrode was used to inject current for estimating the value of membrane resistance. The heart-aorta preparation was bathed in a continuously flow-ing physiological saline containflow-ing (mill): Na+, 306-6; K+, 6; Ca+a, 13.5; Cl~, 337-2; CO3H~, 2-4. Changes in K+ content were made by removing KC1 without
compensa-tion for the changes in osmolarity. Low Na+ solucompensa-tions were prepared by substituting LiCl or choline chloride for NaCl. Chloride deficient solutions were prepared by substituting 90% of the Ringer chloride with impermeant methylsulphate anions.To test the action of low temperature, cool solutions were introduced into the experi-mental chamber, the temperature of which was monitored regularly. The mechanical tension of the heart was recorded with a RCA 5734 transducer.'
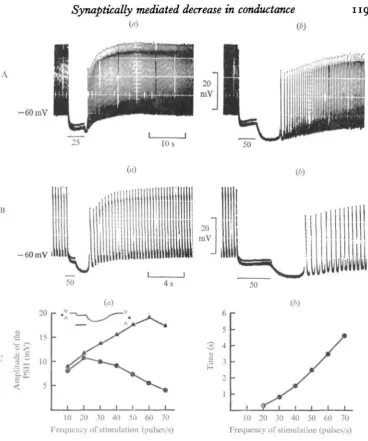
In preliminary experiments it was found that a 4-53 period of stimulation at a rate of 50 pulses/s produced post-stimulation hyperpolarization that was nearly maximal and did not fatigue the heart; this was therefore adopted as the standard stimulation regime in the present experiments. The amplitude of the hyperpolarization following application of the inhibitory trains was measured with respect to the value of normal diastolic potential (EM); its duration was the time for EM to return to its normal value.
RESULTS
General aspects of post-stimulation hyperpolarization
As observed by Delaleu & Holley (1976), cardio-inhibitory stimulation of the heart of Porcellio markedly affected both the time course of spontaneous electrical responses and EM. Heartbeat stopped with stimulation frequencies around 10 pulses/s, and beyond 20-25 pulses/s the post-stimulation hyperpolarization was observed at the cessation of the inhibitory train. The PSH could be recorded with both KC1 or K2SO4
- 6 0 mV
mV
(a)
- 6 0 m V ' l L,
50 4 s
o > 20 r •
15 10
(b) 6
5
^ 4
s 3 H
10 20 30 40 50 60 70 Frequency of stimulation (pulses/s)
[image:3.451.41.410.31.473.2]10 20 30 40 50 60 70 Frequency of stimulation (pulscs/s)
Fig. 1. The PSH recorded in Porcellio myocardium. (A). The duration of stimulation of the cardiac nerves is kept constant (5 s) and the frequency raised: 25 pulses/s in (a); 50 pulses/s in (6). (B) The frequency in the train is kept constant (50 pulses/s) and its duration varied: 800 ms in (a); 3400 ms in (b). (C) Changes in the PSH versus the frequency recorded from a heart without spontaneous activity. Train duration: 5 8. (a) Changes in the amplitude of the PSH (triangles) and of the level of EM during stimulation (circles). See inset diagram. (6) Changes in the time from onset to peak. Resting EM = — 60 mV. In this figure and in the following, the horizontal bar indicates the duration of the train, and the numbers indicate the stimulation frequency (pulses/s).
)
Control After 5 min with picrotoxin
- 6 5 m V
CO
After 5 mia with picrotoxin
I i ! II t
i i m u t u i
10 s
20 mV
Fig. 2. Effect of picrotoxin and changes in membrane resistance. (A) (<j) PSH control in normal solution. (6) Maximal hyperpolarization after 5 min in the presence of picrotoxin (16 x io"1 ITIM). (B) Changes in Ru in normal saline (a) and in picrotoxin saline (6) recorded on another heart. Pulse intensity: — 6 x io~* A. (C) Changes in RM observed in another heart. In B and C, the upper part of the spontaneous electrical responses was intentionally suppressed.
Effect of picrotoxin
- 5 8 mV. 20 raV
[image:5.451.55.414.47.173.2]10 s Fig. 3. Behaviour of the P S H during experimental changes in EM- (a) Control in normal solution; (A) Increase in P S H upon injecting — 5 x 1 o " ' A current; (c) Reversal of P S H during the injection of 5 x i o "8 A current. In this experiment EM was —58 mV and £peH —50 mV.
A effect would be represented by the summated IJPs and the B effect would induce the PSH. As previous experiments suggested that GABA could be the transmitter of the A effect (Delaleu & Holley, 1976), we used its antagonist, picrotoxin (i-6 x icr1 HIM) in an attempt to selectively suppress the A effect.
Picrotoxin (PTX) was able to dissociate the A and B effects, as it reversibly abolished the A effect. The myocardial membrane maximally hyperpolarized during stimulation, which could signify that the B effect was unmasked right at the beginning of stimula-tion. This is clearly illustrated by the traces in Fig. 2, A and B. Similar experiments with 10 hearts provided identical results. It should be noted that PTX could not block the inhibitory effect of stimulation on the cardiac ganglion, for the heartbeat stopped even though the A effect, in the myocardium, was strongly reduced. How-ever, trains at moderate frequencies (25 pulses/s) that suppressed the spontaneous rhythmicity of some hearts in normal saline were seen only to reduce it during pro-longed exposure to PTX.
Changes in membrane resistance
In a preceding paper (Delaleu & Holley, 1976), it was reported that changes in membrane resistance (RM) of the myocardial membrane were associated with the stimu-lation of the cardiac nerves. In the present study special attention has been paid to the changes in RM occurring during the PSH. A large increase in RM was always recorded and the magnitude of the resistance change appeared to parallel the amplitude of the PSH. In some instances this increase reached 100% as illustrated by the traces in Fig. 2 B, a. In the presence of PTX, the increase in RM occurred immediately at the onset of stimulation and was contemporaneous with a large hyperpolarization (Fig. 2B, b). This effect contrasted with the decrease in RM frequently observed during stimula-tion in normal saline (Delaleu & Holley, 1976, and Fig. 2C).
Effect of changes of membrane potential on the PSH
cur-Control [K+]o = 6 mM [K+]o = 0 after 5 min [K+]o =6mM
A -65 mV 40 niV
[K+]o = 0 after 15 min [K+], = 0 after 17 min 10 s
B -53 mV
10 s
20 mV
(a) CO
Control TEA = 10 mM after 2 min
40 mV
10 s
Fig. 4. Changes in the PSH in K+-free solution and in presence of TEA. (A) (a) Control in normal saline; EM = — 65 mV; (6) after 5 min in K+-free saline; (c) restoration of the normal [K+]o. (B) Another heart. Suppression of the PSH after 15 min in K
+
-free saline, (a) Partial restoration of the PSH by injection of — 5 x io~8 A current (A). (C) Another heart, (a) PSH in normal saline; (6) the PSH after 2 min in TEA solution (10 mM). The horizontal line represents electrical zero.
rents. Similar observations were made with a total of ten hearts, examples being given in Fig. 3 and Fig. 7B. The PSH reversed around —50 mV, i.e. at a value of 10 mV less negative than the normal diastolic polarization. In most of the hearts tested, the level of EM during inhibitory stimulation (the A effect) was little changed by the con-ditioning current, as illustrated in Fig. 3. However, in some preparations, EM during stimulation shifted in the expected positive direction during the passage of inward currents.
Effect of potassium-free solution and tetraethylammonium chloride
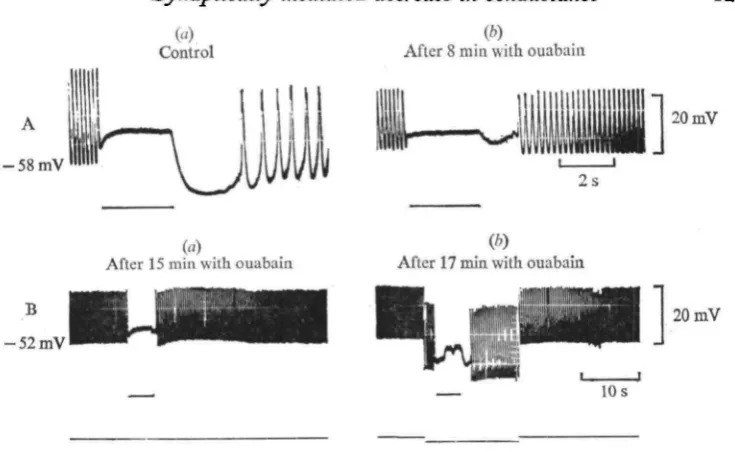
K+-free saline reversibly reduced or abolished the PSH within a 5-10 min period (6 experiments). As shown in Fig. 4A, c, the usual marked hyperpolarization appeared upon restoration of the K+ level, presumably due to the activity of an electrogenic Na+ pump (see Delaleu & Holley, 1975). Inhibitory stimulation of this hyperpolarized myocardium elicited a PSH whose amplitude was somewhat increased, compared to the control. Additional experiments provided evidence that a PSH suppressed by prolonged washing in K+-free saline could be partially restored by injecting an inward current in the myocardium (Fig. 4B).
Control After 8 min with ouabain
20 mV
()
After 15 min with ouabuin After 17 min with ouabain
B
-52mV
20 mV
[image:7.451.43.411.38.264.2]10 s
Fig. 5. Effect of ouabain on the PSH. (A) (a) PSH in normal saline; (6) the PSH after 8 min in an ouabain containing saline (1-7 X io-'mM). (B) Another heart, (a) Suppression of a PSH by a
15 min washing in ouabain solution, and (6) partial restoration by application of — 5 x io~* A current.
The spike generating effect of TEA was thought to be mediated by a decrease in K+ conductance and it was thus of interest to test its action on the PSH. PSHs could be recorded when trains were applied during TEA induced activity (Fig. 4C, b). Atten-tion must be drawn to the similarity of the time course of the responses immediately following the cessation of stimulation in TEA saline with those usually observed in normal medium (C, a): the overshooting spikes originating from the plateau phase before stimulation were temporarily suppressed.
Effect of ouabain and cooling
The PSH was generally decreased or abolished when ouabain (1-7 x io~2 HIM) was added to the bathing medium (Fig. 5 A). As with K+-free saline, the PSH could be partially restored by passing inward current across the myocardial membrane (B).
The effects of cooling were somewhat unclear but it appeared that lowering the temperature to 5-6 °C had a depressing effect on the PSH. In some hearts it was abolished whereas in others its amplitude was not noticeably reduced although its time from onset to peak was markedly increased.
Effect of lowering the external Na+ concentration
Control After 2 min with Ii+ QNa() +]0 =0)
n. i.u.t.i.i.i.:. u
V/v
Control. [Na+]0 = 306-6 nut
20 mV
[Na+] 0= 7 6 nut (choline)
- 6 5 m V
Fig. 6. Effect on the PSH of varying the external Na+ concentration. (A) (a) PSH in normal solution; (6) after a min in Na+-free (Li+) saline. (B) (a) Control in normal solution; (6) PSH after i min in Na+-deficient medium (75% of external Na+ are replaced by choline ions).
Effect of lotoering the external chloride content
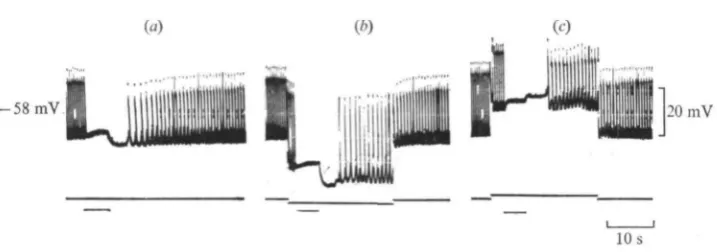
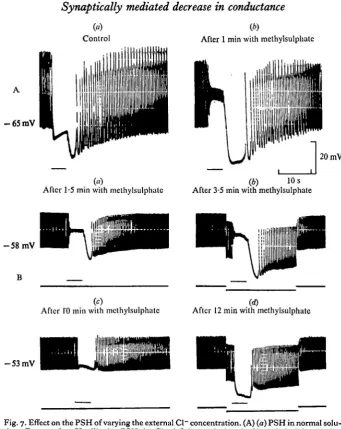
Within 60 s in Cl~ deficient solution (methylsulphate) the summated IJPs of the A effect always shifted in the positive direction (by 20 mV in Fig. 7A, b)\ this agrees with previous results suggesting the implication of Cl~ in their electrogenesis (Delaleu & Holley, 1976). By contrast the PSH was usually unmodified or sometimes slightly increased as shown in Fig. 7 A. Three experiments made on different hearts where it was possible to record continuously from the same cell indicated that the time course of the PSH varied with the duration of the perfusion (Fig. 7B). After 1-2 min, the changes in EM analogous to those reported above were recorded (Fig. 7B, a) and the injection of a —io~7A inward current to the myocardium (Fig. 7B, b) increased the PSH, as in the normal medium (see Fig. 3). After 10 min, the level of the sum-mated IJPs tended gradually to approach the resting membrane potential and the PSH was markedly decreased (Fig. 7B, c). Application of the same inward current then caused the membrane to maximally hyperpolarize as soon as the beginning of the train (Fig. 7B, d). This last finding is comparable to some extent to that observed in the presence of picrotoxin (cf. Fig. 2).
The PSH and contraction
((4
Control
-
(4
Aficr 1.5 min with mcthylsulphnte
(b)
After 1 min with methylsulphatc
-
"U'
J 2 0 m V(4 10 s After 3.5 inin with methylsulphate
( c )
After I'O mi11 with methylsulphate After 12 min with methylsulphate (4
Fig. 7. Effect on the PSH of varying the external Cl- concentration. (A) (a) PSH in normal solu- tion; EM = -65 mV; (b) the PSH in CI--deficient solution (90% rnethylsulphate) after
I min. Note the shift of the A effect. (B) Another heart. (a) PSH after 1.5 min in methyl-
sulphate saline; (b) application of - I. x 10-' A current; (c) after 10 min; (d) application of the
same current; the myocardial membrane maximally hyperpolarized at the beginning of the train.
P S H caused a large relaxation of the myocardium (Fig. 8A). I n other preparations, the heart muscle maximally relaxed at the onset of stimulation and the P S H did not cause additional relaxation (Fig. 8B).
DISCUSSION
[image:9.451.52.394.29.459.2]- 6 2 m V
MAMA/
1
Is 2s
Fig. 8. The PSH and the mechanogram. (a) Relaxation of the heart caused by the PSH; (6) in another heart, the PSH provided no additional relaxation. The spontaneous electrical activity is in the upper part of the figure, the mechanogram in the lower.
cean heart. However, in the heart ganglion of Squilla, Watanabe, Obara & Akiyama (i968)]have reported that the membrane of the pacemaker neurones is sometimes hyper-polarized after stimulation of the a nerves (inhibitory). These authors noted that this effect sometimes lasted for several seconds and that interburst intervals were prolonged. In Porcellio, cessation of heartbeat (up to 5-10 s) might possibly be attributed to a hypothetical PSH in the neurones of the cardiac ganglion. However, other hypotheses may be proposed to account for these prolonged arrests: a relatively slow decay of synaptic action of the inhibitory transmitter in the ganglion and/or changes in mechanical tension induced by the PSH and communicated to the ganglion that modified or blocked its activity.
The most significant features that characterize the PSH in the woodlouse's myo-cardium are (i) an increased RM (ii) an ' atypical' behaviour with changes in EM (iii) a reversal potential around — 50 mV, i.e. lying close to the resting membrane potential. The results obtained in picrotoxin salines are also of importance since this substance was shown to dissociate the A and the B effect. This finding allows a reasonable understanding of the changes in EM and RM that were recorded. Since the A and B effects have opposite actions on membrane resistance and may combine in different proportions, the increase in RM that was recorded during stimulation can be explained as predominance of the B effect. Conversely, the predominance of the A effect, imply-ing an increased permeability to Cl~, resulted in a decrease in RM during the stimula-tion.
kowever, our findings indicate that the PSH, reduced or abolished in K+-free saline or in presence of ouabain, was partially restored by inward current injection in the myo-cardium. In any case the pump hypothesis does not explain the conductance changes and the voltage dependence of the PSH, since the electrogenic Na+-pump in most tissues does not have these properties (see the review by Thomas, 1972). It should be also stated that, in Porcellio,the myocardial membrane behaves like an ohmic resistance during the passage of inward currents, which rules out the possibility of a rectification being involved in the PSH.
depriva-tion eliminates the A effect, possibly by preventing the synaptically mediated increasj in Cl~ conductance.
It remains necessary to determine the functional significance of the PSH in the physiology of this crustacean heart. The A and B effect could be mediated by a single class of regulatory fibres or by two distinct classes. Following the 'single-class' assumption, a unique neurotransmitter would subserve the two effects. The differen-tial action of PTX on the A and B effect does not argue against this hypothesis because picrotoxin, in antagonization of GABA, is known to interfere with conductance changes rather than with transmitter-receptor interaction (Takeuchi & Takeuchi, 1969). The 'two-class' assumption would be that a category of fibres mediate the post-synaptic change in Cl~ conductance (the A effect) while another category of fibres, releasing a different transmitter, induce the Na+-K+ conductance decrease (the B effect). Bearing in mind that cardiac nerve stimulation could excite both acceleratory and inhibitory fibres (Delaleu & Holley, 1976), the excitatory component could be res-ponsible for the B effect. However, it is not at all clear how the decrease in conductance could subserve a reinforcement of the heart activity. On the contrary, this effect could be better connected with inhibitory processes. Indeed, given that the spon-taneous electrical responses of the myocardium to the heart ganglion output imply an increase in both Na+ and K+ conductances (Delaleu et al. 1972), it becomes con-ceivable that the opposite action of the B effect inhibits these responses. Cl~ conduc-tance increase and Na+-K+ conductance decrease would co-operate. The mediation of this unusual decrease in conductance is unknown. It is possible that a synaptic trans-mitter directly operated the change in conductance by turning off ionic channels in the heart membrane. Alternatively, it may result from the suppression of a continuous release of transmitter by the cardiac ganglion, a release which normally maintains Na+ and K+ conductances at relativly high values. The stimulation of the cardiac nerves would then inhibit the permeabilizing system, either in the cardiac ganglion or in the implicated presynaptic fibres. We have no experimental data supporting this hypothe-sis, which has previously been proposed in the vertebrate retina (Dowling & Ripps, 1973). In Porcellio, the finding that the PSH could be recorded in a heart without spontaneous activity, i.e. when the rhythmic activity of the cardiac ganglion was blocked, does not seem to favour the proposal, but this does not necessarily implicate the blockade of the postulated permeabilizing system.
I am indebted to Dr H. M. Gerschenfeld for making valuable suggestions and to Dr A. Holley for helpful discussion during the preparation of the manuscript. This work constitutes partial fulfilment for the requirements of a Doctorat es Sciences,
CNRS19785.
REFERENCES
BRODWICK, M. S. & JUNCE, D. (1972). Post-stimulus hyperpolarization and slow potassium conduc-tance increase in Aplysia giant neurone. J. Phytiol., Lond. 333, 549—70.
DELALEU, J. C. (1974). Structure et physiologie d'un coeur a automatisme neurogene (Crustace Isopode): myocarde, relations neuro-musculaires, facteurs de la regulation. These de Doctorat es Sciences. A.O 9785 Lyon.
DELALEU, J. C , BLONDKAU, A. & HOLLBY, A. (1972). Electrophysiology of the heart of an Isopod
Crustacean: Porcellio dilatatus. II. Effects of ions and membrane permeability inhibitors. J. exp. Biol. 57, 609-31.
DELALEU, J. C. & HOLLEY, A. (1973). Modalites et mecanismes de l'inhibition peripherique dans le coeur d'un Crustace Isopode. J. Phytiol., Parii 67, 184A.
DELALEU, J. C. & HOLLEY, A. (1976). Contribution of an electrogenic pump to the resting membrane polarization in a crustacean heart. J. exp. Biol. 64, 59-74.
DELALEU, J. C. & HOLLEY, A. (1976). Neural regulation of the heart muscle in an isopod crustacean: acceleration and peripheral inhibition. ,7. exp. Biol. 64, 345-356.
DOWLINO, J. E. & RIPPS, H. (1973). Effect of magnesium on horizontal cell activity in skate retina. Nature, Lond. 342, 101-3.
GERSCHENFELD, H. M. & PAUPARDIN-TRITSCH, D. (1974). Ionic mechanisms and receptor properties underlying the responses of molluscan neurones to 5-hydroxytryptamine. J. Phytiol. Lond. 343, 427-S6.
Hoi,LEY, A. & DBLALEU, J. C. (1972). Electrophysiology of the heart of an isopod Crustacea: Porcellio dilatatus. I. General properties. J. exp. Biol. 57, 589-608.
KUFFLER, S. W. (i960). Excitation and inhibition in single nerve cells. Harvey Led. 54, 176—218. NAKAJIMA, S. & TAKAHASHI, K. (1966). Post-tetanic hyperpolarization and electrogenic Na pump in
stretch receptory neurone of crayfish. J. Phytiol., Lond. 187, 105-27.
RANG, H. P. & RITCHIE, J. M. (1968), On the electrogenic sodium pump in mammalian non-myelinated fibres and its activation by various external cations. J. Phytiol., Lond. 196, 183-221.
TAKEUCHI, A. & TAKEUCHI, N. (1969). A study of the action of picrotoxin on the inhibitory neuromuscular junction of the crayfish. J. Phytiol., Lond. 305, 377-91.
THOMAS, R. C. (1972). Electrogenic sodium pump in nerve and muscle cells. Phytiol. Rev. 53, 563-94. WATANABE, A. OBARA, S. & AKIYAMA, T. (1968). Inhibitory synapses on pacemaker neurons in the
heart ganglion of a Stomatopod. Squilla oratorio. J. gen. Phytiol. 53, 908-24.
WEIGHT, F. F. & PADJEN, A. (1973a). Slow synaptic inhibition: evidence for synaptic inactivation of sodium conductance in sympathetic ganglion cells. Brain Ret. 55, 219—24.
WEIGHT, F. F. &PADJBN, A. (19736). Acetylcholine and slow synaptic inhibition in frog sympathetic ganglion. Brain Ret. 55, 225-8.
WEIGHT, F. F. & VOTAWA, J. (1970). Slow synaptic excitation in sympathetic ganglion cells: evidence for synaptic inactivation of potassium conductance. Science, N.Y. 170, 755—8.