MALE-SPECIFIC LETHAL MUTATIONS
OF
DROSOPHILA
MELANOGASTER.
11.
PARAMETERS OF GENE ACTION
DURING MALE DEVELOPMENT
JOHN M. BELOTE
Department of Biology, University of Calijornia, San Diego, La Jolla, Calijornia 92093
Manuscript received May 25, 1983 Revised copy accepted August 1 , 1983
ABSTRACT
The male-specific lethal mutations (msl's) identify loci whose wild-type gene products are essential for male, but not female, viability. Earlier studies in which X-linked gene activities were monitored in msllmsl male larvae demon- strated that these genes are responsible for setting and/or maintaining the level of X chromosome transcription in males (i.e., they are necessary for proper dosage compensation). The present study examines several important questions concerning their mode of action during development-The results of an examination of the effects of an msl-1 deficiency on male-lethal phase and female viability suggest that this mutation is an amorph, or a severe hypo- morph. The effects of rendering a fly mutant for more than one male-lethal mutation were also examined. Multiply mutant flies were no more severely affected than singly mutant ones. A gynandromorph analysis revealed that the male-limited lethality associated with msl-2 has no single lethal focus. Somatic clones of homozygous msl-2 cells were initiated at various times during devel- opment by X-ray-induced mitotic recombination. An examination of the via- bility, growth patterns and morphology of marked clones demonstrated that:
(1) msl-2+ acts in a cell autonomous manner, (2) msl-2+ function is required not only in larval (polytene) cells as was shown in previous work but is also needed in the diploid cells that give rise to adult structures, (3) the msl-2' gene is needed fairly late in development and perhaps continuously, (4) the msl-2 mutation does not affect sexual differentiation.
HE male-specific lethal mutations of Drosophila melanogaster (e.g.
,
mle, msl-T
1 , msl-2, mEe(3)132, msl-3) identify loci whose wild-type products are es- sential for male viability (FUKUNAGA, TANAKA and OISHI 1975; BELOTE and LUCCHESI 1980a; UCHIDA, UENOYAMA and OISHI 198 1; LUCCHESI, SKRIPSKY and TAX 1982). Males (=,AA; in which A signifies a haploid set of autosomes) homozygous for any of these autosomal recessive mutations exhibit prolonged posthatching development and eventually die during the late larval or early pupal stages. BELOTE and LUCCHESI (1 980b) demonstrated that the physiolog- ical basis for this male-limited lethality is a reduced level of X chromsome gene activity in affected males. Our interpretation of this finding was that the male- specific lethal loci are involved in the regulatory hierarchy controlling dosage compensation (for reviews see LUCCHESI 1977, 1983, STEWART and MERRIAM1980), and that their wild-type gene products function in males to set and/or
a82 J. M. BELOTE
maintain the transcription of the X chromosome at a level approximately twice that which occurs in females. T h e absence of phenotypic effects of these mu- tants in females (XXAA) suggests that these genes are not normally expressed in the female soma, and, consequently, the
X
chromosome is transcribed at the lower level characteristic of females.During development the X A ratio provides a signal to influence both the fly’s dosage compensation mechanism and its pathway of sexual differentiation. One gene, Sex-lethal (Sxl), has been identified that plays a pivotal role in both processes (CLINE 1978). Thus, mutations at this locus not only alter the level of X chromosome gene activity in affected individuals (LUCCHESI and SKRIPSKY
1981; CLINE 1983) but also affect their sexual development (CLINE, 1979a, 1983; SANCHEZ and NOTHIGER 1982). A maternally acting gene, daughterless
(da), encodes a product that is thought to function in the zygote to control the expression of the Sxl locus and, thus, is also implicated in the control of both dosage compensation and sex determination (CLINE 1976, 1978, 1980, 1983). T h e existence of mutations at other loci that affect only sex determi- nation (e.g., dsx, tra) or dosage compensation (e.g., mle) but not both, implies that the control of these two pathways diverges subsequent to the functioning of Sxl+
(BAKER
and RIDCE 1980; BELOTE and LUCCHESI 1980a).T h e finding that males homozygous for any of the male-specific lethals ex- hibit a level of X-linked gene activity that is 50-65% of that seen in wild type might mean that this level of X activity represents the basal level of transcrip- tion that is seen for the autosomes of both sexes and for the Xs in females. According to this view, the msl’ gene products act in males in conjunction with the cells’ general transcriptional machinery to approximately double the transcription of X chromosomal loci. An alternative view, however, is that the
msE+ genes are necessary for normal X chromosome activity in both sexes and this low, but nonzero, level of X activity observed in mutant males is the consequence of “leakiness” of the particular male-specific lethal mutants that have beenstudied. One important question, then, is whether any of the extant mutants represent a complete loss of function at the locus involved (i.e., are they amorphs?). T h e identification of amorphic alleles is also important for understanding the domain of function of the wild-type gene. In particular, one would like to know whether more severe phenotypic effects (e.g., embryonic or early larval lethality; reduced viability in mutant females; abnormal sexual differentiation) can be associated with male-specific lethal mutations, other than those thus far observed with existing alleles.
In addition to these questions, other properties of these loci need to be examined before one can reasonably determine their mode of action during male development. (1) Do these genes act in a cell autonomous manner?
(2)
Are these genes required in all tissues? (3) At what time(s) during development must these genes function?MATERIALS AND METHODS
Lethal phase determinations: The effective lethal phase (ELP) for each of the mutant combinations was determined as described by BELOTE and LUCCHESI (1980a).
Cytogenetic mapping ofmsl-2: Translocation stocks of LINDSLEY et al. (1972) were used to generate homozygous msl-2 males carrying terminal duplications of chromosome arm 2L. The cytological location of msl-2+ was inferred from the observation that homozygous msl-2 males carrying the distal 2 L fragment of T(Y;Z)RI36 or T(y;2)GI46 fail to survive, whereas such males carrying the distal 2L piece of T(Y;Z)GZ20, L I 2 6 , P8, H I 1 6 or R I 2 7 are recovered with normal frequencies. This places the msl-2 locus between the breakpoints of T(y;2)GZ46 and T(y;2)GZ20 which have been determined by ASHBURNER and VELISSARIOU (1980) to be in salivary chromosome regions 23El-2 and 23F6, respectively.
Gynandromorph analysis: Gynandromorphs were generated in the followirig cross: R(I)2,w"/u+Y;
msl-2 cn/+
+
males X y f96"/yy6";
msl-2 cnlmsl-2 cn females. Ring-X/rod-X flies (females and gynandromorphs) homozygous (cn in phenotype) and heterozygous (m') for msl-2 were preserved in three parts 70% ethanob1 part glycerol, then eviscerated and mounted in Euparol between coverslips according to the method of SZABAD (1978). Male tissue, phenotypically yellow with forked bristles, was scored as described by BRYANT and ZORNETZER (1973).Clonal analyses: For blastoderm irradiations, y/y; msl-2 pun/In(2LR)CyO (experimental) or y/y; pwn/In(2LR)CyO (control) females were crossed to y/y; M(2)z Dp(I;2)Sc",y+ pr/In(2LR)CyO males in population cages, and eggs were collected at 2-hr intervals on 14-cm diameter Petri plates con- taining standard medium sprinkled with dried yeast. Two hours later, plates were irradiated (Torrex 150 X-ray Inspection System, 120 kV, 5 mA, 0.5-mm aluminum filter) at a rate of 350 r/min to a final dose of 500 r. Larvae were allowed to develop on the Petri plates and were transferred as third instars to quarter-pint bottles containing fly food to complete development. Adults of the appropriate genotype were collected, aged 1 day and preserved. Whole bodies were eviscerated and mounted in Euparol between coverslips and were examined with a Zeiss Universal microscope.
For the larval irradiations, eggs were collected as described earlier and then transferred to split quarter-pint bottles. At the appropriate time, cultures were irradiated in the lower part of the bottle (1000 r, 1-mm Plexiglas filtration), and the adults of interest were prepared for microscopy. Only patches with more than one bristle were scored, since single-bristle clones are sometimes ambiguous and do not always represent a clone generated by mitotic recombination at the time of irradiation (GARCIA-BELLIW 1972).
For the analysis of clones induced in the abdominal histoblasts during larval development, asynchronous larval cultures of genotype y ; Dp(Z;2)sc'g,y+/msl-2 p u n (experimental) or y Dp(I;Z)s~'~,y+/@n (controls) were irradiated in the lower portions of split bottles (1000 r, 1-mm Plexiglas filtration), and pupae were collected every 24 h. After eclosion, flies were aged 2 days. Their abdomens were then removed, eviscerated and mounted on microscope slides. Tergites 11- VI were scored for mosaic spots ( y or pun); only clones encompassing more than one bristle were counted. Since bristle-forming cells can undergo extensive cell movement during formation of the tergite, all similarly marked bristles in the same hemitergite were considered to belong to the same clone (GARCIA-BELLIDO and MERRIAM 1971).
RESULTS
The nature
of
extant alleles and mutant interactions: T h e utility of mutational analysis to infer the normal function of wild-type genes depends on an under- standing of the nature of the mutations used. To examine the nature of a typical male-specific lethal (to be referred to collectively as msl's), comparisons were made of female viability and male ELP in individuals homozygous fora84 J . M. BELOTE
2 + locus (Df(2L)203, WRIGHT et al. 198 1) have been compared. No differences
in either male lethal phase or female viability and morphology were observed (Table 1). These results suggest that msl-2 is functionally either a severe hy- pomorph or a null allele.
Double and triple mutant chromosomes were constructed via meiotic recom- bination, and the effects of rendering an individual mutant for more than one msl were observed. Lethal phase determinations for flies carrying msl-2 msl-1, msl-1 mle, msl-2 mle and msl-2 msl-1 mle are shown in Table 1. Although the frequency of hatchiwg is less in the multiply mutant flies than in single mutant strains [-80% us. -go%, see TANAKA, FUKUNAGA and OISHI (1976) and BE-
LOTE and LUCCHESI (1980a) for single mutant data], the fact that there is no difference in the fertility of homozygous and heterozygous mothers suggests that this decrease is not the result of the msl’s
per
se but perhaps is due to background effects. In all cases the number of surviving adults and their gen- otypic segregation are close to what one would expect if only homozygous males are dying posthatching, and that the failure to hatch is random. This implies that there is litttle, if any, effect on mutant female, or on heterozygous male, viability. Although the attrition rate of young mutant male larvae ap- peared to be greater in the triple-mutant crosses than in the single- or double- mutant ones, some males homozygous for msl-2 msl-1 mle were, nevertheless, capable of surviving to the late third instar stage.Gynandromorph analysis: X chromosome transcription is regulated autono- mously in gynandromorphs (LAKHOTIA and MUKHERJEE 1969): in XX//XO mo-
saics, the XO cells exhibit the high level of transcription characteristic of males, whereas X X cells have X chromosome activity typical of females. If the function of the msl+’s is to regulate dosage compensation in males, then this observation predicts that the male-specific lethality associated with msl’s be cell autonomous and have no particular site of lethal action ( i . e . , no single lethal focus).
UENOYAMA et al. (1982) examined this question by producing gynandro- morphs homozygous for one of three msl’s (mle, mle(3)132 and m~l-2‘~). In all cases a very low frequency of gynandromorphs was observed, and those that were recovered had small, abnormal XO patches. I have obtained similar results using the original msl-2 allele. In a cross that was expected to yield 36 msl-2/ msl-2 gynandromorphs, none survived to adulthood. It is unlikely that X X / / XO;msl-2/msl-2 mosaic embryos grew up with the X X cells completely competing out the XO cells to yield normal females since there was no increase in the number of XX;msl-2/msl-2 females recovered (six expected, five observed).
Although these results indicate that the male lethality associated with msl-2
has no single lethal focus, they d o not provide a test of the cell autonomy of
msl-2+ gene function, since all cells are msl-2/msl-2. Nor does such an experi- ment demonstrate whether the msl-2+ gene is required in male imaginal cells for their viability, since the X is lost so early that large regions of larval tissue are necessarily rendered XO. T o examine these questions, and to determine the time($ at which the msl-2+ gene must function, a clonal analysis was carried out.
W .3 0 CO n Q, e4
s
P-*
P- v m5.
P- m t-s
2
5% sf: W 0 0 011 m n 0 P- t- W P-a
-
E Q, 32
E 0 0 e4 h2
s
n n 3 0 0*
P- 011 3 0 CO2
m2
hz
s
m P- nF
3 f-, m t- na
3 Q, 01 e4 In v 0 Q) m 0 I- O W W In W2
W 0 0.I v 3 h e mj E 011 0 e4 h m! E P-n I
0 Q, n 00 n 0 0 m 0 m
s.
2
m$
s
2
a
m*
CC n n v m 0 0*
2 I 0 P-2
P- Q, h-
Ir; f-, m n n2
f-, -I- -I- n h 9e
In0 m -I- 0 0 W 9,
-
E1 m W 0 0 In
-
2
-
h 3 L 6 m 011 n
s
W 0*
n f-, h2
e
m m 0 0 t- 9,-
E1
3%
2
3
886 J. M. BELOTE
STERN 1936; GARCIA-BELLIDO 1972; BECKER 1976; RIPOLL 1977 for discus- sions of this technique). T h e viability, growth patterns and morphology of the resulting marked clones lacking msl-2+ function provide information not only about the question of cell autonomy but also indicate in which tissues msl-2+ must act. Furthermore, by inducing such clones at different stages of devel- opment, one can demonstrate the latest time that msl-2' must be present in particular cell lineages for normal growth and differentiation to occur.
Figure 1 shows the genetic constitution of irradiated individuals. T h e posi- tion of the msl-2 locus was determined as described in MATERIALS AND METHODS to be in salivary chromosome region 23E1-2 to 23F6. This is just distal to the M(2)z locus at 24E-25A, and the y+ insertion of Dp(1;2)~c'~,y+ in region 25-
26. Any single crossover occurring proximal to the y+ marker will render the msl-2 mutant homozygous in daughter cells that exhibit loss of the
y+
pheno- type. Such a crossover, followed by the appropriate segregation, will result in the M(2)z mutant being concomitantly lost. If the msl-2 mutation has no effect on the cells' growth, these clones will have a substantial growth advantage over background cells, which are still Minute (MORATA and RIPOLL 1975). As an internal control, the right arm of chromosome 2 is heterozygous for the bristle and trichome marker pawn ( p w n , GARCIA-BELLIDO and DAPENA 1974). T h e frequency and size of pwnlpwn clones provide a relative standard against which the frequency and size of yellow clones (i.e., msl-2/msl-2) can be measured. An independent control is provided by comparisons of clone frequency and size in males and their female sibs, where loss of msl-2+ has no effect. In some experiments males not carrying msl-2, but otherwise genotypically similar to\
x.ray\
"7.9-l + + . .
msl-2 t v*n
I
PHENOTYPE
Minute+ y e l l o w m s l - 2
M/ M
cell lethal
Minute pawn
Minute
the experimentals, afford a third standard of comparison for determining the effect of msl-2 on viability and growth of clones.
Tables
2
and 3 present the results of blastoderm irradiations (3.0 & 1 hr postfertilization). Since the viability and penetrance of these markers in somatic clones is high (GARCIA-BELLIDO 1972; GARCIA-BELLIDO and DAPENA 1974), the frequency of yellow clones in the experimental classes can be assumed to equal the frequency of msl-2/msl-2 patches.In females and control males the difference between the mean size of y us.
pwn clones can be attributed to the fact that
y,
but not pori, clones are Minute+ and, thus, exhibit an increased mitotic rate resulting in larger clones (MORATA and RIPOLL 1975). The differences between the frequency ofy
vs. pwn clones in these flies is due to both the larger size (and, hence, the increased probability of encompassing a bristle) ofy
clones and the fact that thepon
marker is nearer to the centromere than they+
insertion is, and, therefore, crossover events proximal to pwn are less likely to occur.The frequency and size of pwn clones are similar in males and females, experimentals and controls, indicating that the efficiency of X-ray-induced mitotic recombination is approximately the same in these genotypes, and that the growth and scorability of mosiac spots in general do not differ substantially in males and females. Moreover, in control males (not carrying msl-2) and in females, the frequency and mean size of
y
spots are similar. The striking decrease in both frequency and size ofy
clones in experimental males indicates that homozygous msl-2 cells, although not completely lethal, are at a severe growth disadvantage, even when they are M + in an M background. This ob- servation proves that the msl-2+ gene acts in a cell autonomous manner, and that it functions in imaginal discs and abdominal histoblasts at some point after blastoderm formation. The lack of any difference between control and exper- imental females (Tables2
and 3) iny
clone frequencies and sizes greatly strengthens the assumption that the wild-type msl-2' gene carries out no essen- tial function in females. Thus, not only does the msl-2 mutation have no dis- cernible effect on the viability and development of female flies (BELOTE and LUCCHESI 1980a) but it also has no apparent effect on the viability and growth parameters of individual cell lineages in females.Clones were also examined in the imaginal disc derivatives of irradiated third instar larvae (Figure 1; Tables 4 and 5). Again, the frequency and size of pwn
888
E
0
._ U
._
i
.f: k 9 2 s 3 -Q._
2
2
-
s
o " ? J
-
5 3
2 % -
ii ,'1 &
s s
-
g D-
U b U d 92 U
-
u n
E:
J. M. BELOTE
L
os, iijz U
m "e"
8 $ij m
-
i
8
+I ' w * * t . *
8
P
C C E . S I Cz
pc?::
2
0
&j
ao s 0
-
W W W ~ ~-
3 --
P-
s
$
.- 8
$2
-
2
s
t l
P
w * * w *s
$
yE E c m c
5
. g 4
9 ? ? O r - ? e
T
z
g
.i
E L q
-
m h c P - m m 3C I U i 2
-
0
4
:@
0 0 - o o m m m m m o o V Yu
E
c
5
.o
$ $ Z
m e o o o - * - . + m+I $1 +I +I +I +I +I 9
a 8 'j; 6
g
E de
m 0
d
c r ."
U %
o h ; v1
$ E
z u
c +
a
rAS U
-
0 0 0 - 0 - - - w m ~ 0
O h
0 - 0
+I t I +I t l +I +I +I 5
2
c mf-
z u h O
o s , w o o o o w i Q i w u l m w
iij3 U A A A
. ^ Y
z
3
.G
:
9 k
+I ":
2
9 c ? ? 4 ' 4z : ;
t u 0+* 8 w - o o ~ w w e w o -..
3
j
q 5 A A A m m m - P - o? 0
5:
*5
g c
m
2
+ #i
2 2
2h;
q
. .2o C f i m w m w a w m m n ,
-
es
X &
0 s
.&
.^ a:g
fg
*L,
3
g
$ A T A A A 528 L,
o n
Z
z
m C E k $
S
a
u
z
o o o o% C ? & ! . . . t * * W m ~ ~ ~ K8
C i%*z 0 .5
" 0 t l ~ " ~ + I + I + I t I + I + I t I +I 5 4 8 0 c
z
g
mc v j t . A A A A Y ? l . O O O Q1 $ % b j 3s
z
2 r - m d . w w n 0.I2
.ll;3gs- e . - -
? o n - G
g m c 0.500
$ A : h P P
0 m c
&
k.?
j i . 2
c
52
2
2
T o g $ -M M 3 * 5 0
2.&$
2 & 0 2 y y z $ r m T O &
x 2 .&G.i;o.&.Gm
$ 2
%-&,TI 2 g3:
g 2 Z T j 7 J c C c e c
gE t 1
= = c-
- - -
c:
2 ' Z Q c v1 t l +I m hc
*
+I $1 $1 tr +I ti +I8;
v1r d d r d r - o j 0
2%T
E gc
z uCI o s , w o o o o m w o w o ~ m
W O
u & s
m u ) o o o m - m . + - e4/ \ - - -
I
+I p-
-
B .N.b U $ J
3
E
'Z e.c
2%
i i v ,
0 8 a e u l l n w a m u J o - c w -
z u
E
0 x nL, *.z c U m e ,
Y
.- .-
'cl
2
-0 x E 2" 9 -
U - c y h y y y h L- h 2.2 L ' - 0 && z y F
h J E = = 0*c' .s % 2
E
% a" C Y ._ E-
9*
P
5
.g
$z/"
t.. y
-
B
kj.
3
'II
.- c' C v B U U (r
m w t - m o o e m
o l e ,
m.5 U
.-
p & j2
mor, mt i
.
U * ? ? , . . I C a Xp z E E m - m o -
9
g
G " +Iz
z
;
2
z
2
z
2
t-
ti
kj
0.z u
$
2
a o n Q z m K -
2
m .E
.s
-
2q
B I E c 3 m 3 E
8
8
2;
$1 +I +I +I +I $1 +I B
=
.YE
2
9 "",2C?$Z
a8
aj2
.g+
Q J a* G E
co rm
E
zz
ti
ti
o o o o o m m m t - m ~ o4
:E
h
2 8
5
-2
.s
p
&2
or, .r! 2m
8
I m
8
+I +I +I +I +I +I +I +I
-
B 'j; 6i
Q. r .-
W
0 " v)
-
- 0 0 3 m m * * o r , ~ 0
3
-
2
c
- -
o l e ,
m.5 U
2
92
.-
+I u v ~ q o *
-
m 'Z n!+
z u
o d u -0 c - 0
(r
m m w O " o r , w * .-
m.5 U
g
3%
g
+I * o ~ ~ w * o * -
8
-
3g p
2
C Oj
9
~ ' " 6 c
"
+I +I +I ti +I +I +I $1 +I % 0 5 .ri
8
q I lz i
2
p C O - 0 E c m E
?=?X?4X$$
t- !??% W M;+3-c3 8s
-
2.1
- u E
E X$
ti5
o - o r , m e m m m L o - - 3 03 %
c e
.&
5 i z
z z
g
c o d e , t - o o o o 3 3 a 3 a a m
u g $ m m o o o m m e - -
-
A A A
g
- O . O ir2;8
$ 5
O+I
2
? d + Q O U - + m$ 2
1
o o o o m a - m m c c
8
k2
. z u= T k 2
2
2
&as
oE
o o - * m b " l?
& 2 % & 0 d 3
3 - 3 o 0
mn.Ezs
2 x gz.5 :g.E 2
;g
* d . t - a m a o m m m m m, 6 B
m c3 - 3 1
p m w :
z =
g
p %'&l4$4*l. 2 "
0 m c .&.E a.%
+I
-
€ 2 VI y Uh 3 2 3 C m o S
e, .z E nir z o g " o o g m . 2 3 C a
b u y u z z $
r
. z ~ ~ ~ T ~ ~Y
c o w U -z u
* O * % C
-2 s.2
O M g
A - - - . -Cr C L j
L *
N L -
ii;> U
*B
-
,& MB
v, d t i 10 m m m +I +I ti +I +I +I +I +I Eg ic v j t 9 - A A A A , . . I q w , 5 q o t-
W i1 2 E U E
c
4
-
%
E .- M m:
G b o 2 q &2
X M .a .&.&l.&.s.&2 Lt:?g$;5;g
0
a2
22azz 2$ s . r a n s . 5 - c s
5 5
i
2.2
g$
& ; z Q p g G8 90 J. M. BELOTE
c
o l w
iijs U
$E5
- f i n - L o 3*
;
22;
2 3
8
4
m o m f i nZgj
* n m o * me
.I d-
c d $1 +I +I +I +I +IC99q 5'4
2
2
0
L x
z u
o l o
;ij U
L
$53
*
* m 2 : =Q p
z
C m w C 0p.
a
' Z O c y1 +I +I +I +I +I +Ic y 5 0 o y q
2
h Lr,
4
4
r o ( 0 c o c o 0._
z
2
w,m Un
-+I
.
*..?:a
0s
U U
i r r
O c Q
s
ti
$2
% z u
p @ j * a n f i c o m
w b o w
+I * y ( s q c o *
e
' Z 6 +I $1 +I +I t l +I t Is
--
E n v m n * m._ U
m o - * o m
L
-
O B 0- 3
el .E
2
8
Qw
C m o m m C8
y
2
8 "
0 y c ? ? 0 0-
2
2
a
F: z '
-
o d oB
z
k
;g
3
tis
w c o i o n m -CI
.-
m m
pf+2
U 2 s s g % w+I 9 ? * n o 9
-
m;$
+ I + I t I + I + I t I +IB
!+
2
w m o c o a vm - -
2
q q 5 . 9
q
e
.Ea 0n Q - - . - P
0'?9'1C9"9
-
'i
c x
Z U
0 0
t i j m - ( 0 * ( 0 - m m - 3 -
+I
m 5
.-
6
M ' --
5
2
2
g
m 7 1 2 ~ ~ $
$2
. ~$ 2
p . 5s
2
2
8"
X M M SZ C t - . r * E Z : i l U
p:
$
1
a3 E
w
:
=
2
;
s
v)g
25 %
E
B r
$ C
.o
g
'E.$
h $ 2
71 - & d
9
Ea.?
2 - $ %
cy 0 m q 3
3 2
g.gE
.I1
. s z
&
4 5 %
-5: +I
:
&..&
B E %
.L- 7 1
-
U Y C.^ 61
8
: E ,zg
g z s
C u e+ " C h
-U> w e ,
Y m 2 = $
2
E.p
.E
$8%
3
p
S s g % < z
M.3
x
k$ 3
~ 1 1 s ~ a ~
z2.p
2
g
%u g n - g ~ k
";
:.g
3 s A l $
2
e, m 2 'GO .:
m E . 2 u u
E o 3 8
5
. ~ z m . r j g .
% o g " . s &
z p & &
- Z O > b y
-=
- u : o i .2 u u
& g + E $ $
U *
o h ,
e *
o m 0
z & i
m e e Q ) * n.-
rnS U
n
+I + T o T q q 0
.D
c 4 s +I +I +I +I +I tis
m o w L O y q e 02
2
"mv";a
" * - "
@ e o - - 0 0 0
+I
-
8c
3
z u
e *
o m e r
$E3
" " m "+I * q u - ) q s . ? *
'i. 7
'=
4 +I +I +I +I +I $1 +Ig
m m m * m m ?c\ U
r( .-
v,-m U
n
-8 3 C @ 4 - 0 0 \ I K
8
-e
v)
{
4 " O q ? o q o 0c
;g
m 2 w r - m -s
%
z
u
P
-
o x e rrnS U
E
.e U
c
.-
g & B o . ( * ' c r ) p - - 3
cl
c9 .fi +I * * U ? q y b
9
2 ' = n c ~ +I +I +I +I +I.-
" S
w b q
Ge
4
3
+- g C K O + *8
+I T
s
m 0 0 " 1 0 ? ? n-
-
5
m v m * * 0B
.2
;g
%?
b
c:
z u
B
02 ~ - i c l o a o
e r & g ( 0 o . t v f S m m
e r 0 - e r
-
- c v - o v
N i-
.- CASU
w
U
Cr,
+I 1 1 9 Y ?
8
-
6 +I +I +I +I +I +I +I" I ? " ? c 9 9 Q1
-G ,gw L m m r D -
2
2
4
m m c 0 ( 0 o ? m-
z
c v j2 k
0 0 * e n o ? m m z u
0 J * m l o . ( - c . I
+,
.- M .-
-
02
m
E cd
$
$
g
$
3
g m bo9Z h k , E T O 5
u
c . 2
pp-" 5 $2cd 0
e q ' g
0 6o s 0''
5
9
ei
'
P
.N2
;
o g ,
5 $
9 - 2 .
g
6
3
Y3
,o.g
U
$7
,"
a b2
p z
s
. g > *
"Id Z U B
+ Q = % E
2 ;
5 3 ;
372
CJgs E = -5 2
. L Q+.,g
:&
-,w , C L
r z
> z g2-
E.E
3
4
b % E O
-
d
--
evj
&
% E b m om m
8%
=DL..%a S o k
o E
$ a - d E o E
$zi
2
&. 0 L.. w'
%fozgh%-
3 2 2
~ 2 . z
3 EO i h 4
, g , o . l c c
b m m ;
",g
% zs"GJ+g C a 3 C w E m & m . N C J
2 0 2 ; 5 r
.- m B z _ o 0 ? & , x U o E
A z o > y F
L. O U K
2
119
E au3 J A I g g Z
U b 0 K L . . &
h a c = 3 0
892 J. M. BELOTE
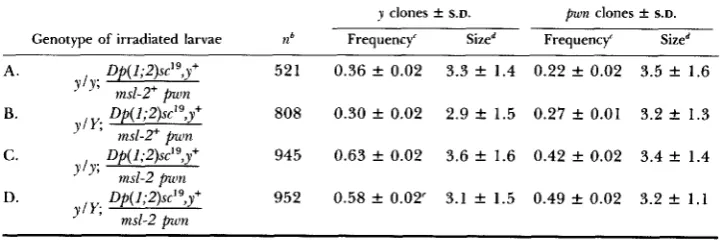
that msl-Z/msl-2 clones encompassing sexually dimorphic regions of the adult exhibit normal sexual differentiation supports the notion that msl's do not normally affect sex determination (see footnotes to Tables 3-6).
Although msl-Z+ function is needed in imaginal disc cells late in development, the results shown in Table 6 show that one can remove the msl-2' gene from abdominal histoblast cells at a time corresponding to the resumption of histo- blast cell division at the early pupal stage (see GARCIA-BELLIDO and MERRIAM
1971 for the parameters of abdominal tergite development and clonal analysis) with little, if any, deleterious effect. A likely explanation of this result is that the abdominal histoblast cells have accumulated enough ms1-P product in their cytoplasm for them to complete development in the absence of continued transcription from the msl-2+ gene during the final burst of cell division (i.e.,
there is a perdurance effect). An alternative interpretation of the data would be that there is no cell competition occurring in the abdominal histoblast cells during formation of the tergites. According to this view each progenitor cell would contribute the same fraction of cells to the adult's tergite regardless of the mitotic rate of daughter cells. T h e existing data do not allow one to distinguish between these alternatives. In any event, the results using the msl-
2 mutant resemble the effect of Minute mutations, which have slighter effects in abdominal tergite clones than in imaginal disc cells (FERRUS 1975).
DISCUSSION
All of the autosomal recessive ins1 mutations that have been extensively studied are characterized by relatively late ELPs and the absence of any overt effects in females. All of these alleles were picked up solely on the basis of their male-differential viability. It is possible that they all represent hypo-
TABLE 6
Clonal analysis of msl-2 in abdominal tergites follming larval irradiation"
y clones & S.D. pwn clones & S.D. Genotype of irradiated larvae ?Zb Frequency' Sized Frequency' Sized
A . Of( 1;2)sc'9,y+ 521 0.36 f 0.02 3.3 f 1.4 0.22 f 0.02 3.5 f 1.6 B. Of( 1;2)sc'9,y+ 808 0.30 f 0.02 2.9 f 1.5 0.27 k 0.01 3.2 k 1.3 C. LIP( 1;2)sc'9,y+ 945 0.63 f 0.02 3.6 f 1.6 0.42 f 0.02 3.4 f 1.4 D. Op( 1;2)sc'qy+ 952 0.58 f 0.02' 3.1 f 1.5 0.49 rt 0.02 3.2 f 1 . 1
y l g ;
msl-2+ pwn
J" ms1-T pwn
J'Y; msl-2 pion
msl-2 p u n
ylv;
" Flies irradiated 0 to 96 hr before puparium formation.
n equals the number of abdomens scored.
Frequency is number of clones per number of abdomens scored.
Size is mean number of bristles per clone; in experimental classes (C and D) y clones are msl- The 129 clones that were in the sexually dimorphic fifth and sixth tergites showed normal
morphic alleles of genes that are essential for both male and female develop- ment, and that the “leakiness” of the mutants allows a threshold
of
function to be reached permitting females to survive. According to this view, the late lethal phases of the various mutant alleles in males may also be a consequence of a low level of residual activity of the msl gene products. As a test of this possibility, mutants of msl-1 were made heterozygous with a chromosomal de- ficiency for the locus, and the ELP and female viability were compared with those of homozygous msl-1 flies. T h e lack of any significant difference between the two mutant conditions suggests that msl-1 is a functionally null allele. A related observation concerns the effects of rendering flies homozygous for multiple msl mutants. T h e finding that individuals simultaneously mutant for three different msl’s are no worse off than singly mutant flies supports the earlier point that the most severe phenotypes associated with lesions in themsl’s are those already seen with extant mutant alleles.
T h e relatively late lethal phases associated with the msl mutants imply that male larvae can survive and grow, albeit slowly, with gross reductions in their levels of X-linked gene products. Although whole-arm hypoploids for auto- somes have been reported to die during the embryonic stage (HERSKOWITZ and SCHALET 1957), it is not valid to assume that msllmsl males, which are essentially hypoploid for a whole chromosome arm (due to their X chromo- somes not being dosage compensated), should die at a similar stage. For ex- ample, msllmsl males should begin to be affected only after the time during development that the msl+ gene normally begins to function. This may be later than the time that autosomal hypoploidy begins to have a detrimental effect. Furthermore, it is not necessarily expected that hypoploidy for each of the five major chromosome arms should exhibit similar lethal phases.
T h e observations that no combination of msl mutants thus far examined
(e.g., msl-llmsl-1, msl-l/Dj(2L)203, msl-2 msl-1 mlelmsl-2 msl-1 mle) have shown any effect on the viability of female individuals (Table l), taken together with the results on the viability and growth of msl-2/msl-2 cell lineages in hetero- zygous females (Tables 2-6), strongly suggest that the wild-type products of these genes are not essentially for any vital process in females.
T h e results of the gynandromorph analysis of msl-2 are reminiscent of what was seen in similar analyses of the mutants da, a female-specific maternal effect lethal (CLINE 1976), and Sxl’#’, a male-specific lethal that also transforms sex (CLINE 1979a). These mutants, like msl-2, have been implicated in the process of dosage compensation (LUCCHESI and SKRIPSKY 1981; CLINE 1983). In those studies no single lethal focus was found, and the patches corresponding to the affected sex were smaller than in controls and were often abnormal. Similar results were seen with gynandromorphs infected with male-killing Sex Ratio (SR) organisms (TSUCHIYAMA, SAKACUCHI and OISHI 1978). Such gynandro- morphs either died before eclosion or exhibited small XO patches. T h e basis of SR infection male-specific lethality, however, is not known.
8 94 J. M. BELOTE
tiating into adult cuticular structures. However, since their experiment did not account for differences that may occur between mutant and wild type with regard to such parameters as growth rate and individual cell viability, their results do not necessarily indicate that mle+ is nonautonomous. The experi- ments described here have the advantage that the mutant male cells can be directly compared with their female or wild-type male counterparts. In addi- tion, one is able to monitor viability, growth and differentiation of the mutant cells in an intact fly.
That dosage compensation is controlled autonomously in gynandromorphs
(LAKHOTIA and MUKHERJEE 1 9 6 9 ) leads to the expectation that the msl+ loci act in a cell autonomous manner to control X chromosome transcription and, hence, viability of male cells. However, there are two possible ways in which these genes may act and still give this result. The ms1+ genes may be required at a discrete (early) stage of development to irreversibly set the activity of the
X chromosome at the high level characteristic of males, this decision being inherited by daughter cells. Alternatively, the wild-type gene products may be required continuously throughout development for the occurrence of male X
hypertranscription. According to the first possibility, if one removes the msl+
gene from a male cell after it has performed its required function, then that cell and its descendents should show no ill effects. Under the second hypoth- esis, removal of msl' function at any time during development should lead to a decrease in X transcription, followed by slow growth and/or death of the daugher cells.
T h e results of the clonal analysis of msl-2 presented here clearly indicate that the msl-2+ gene functions autonomously and that it acts in imaginal disc cells at some point beyond the mid-third instar stage to influence the viability and/or growth of male cells. This finding, together with previous observations
(BELOTE and LUCCHESI 1980a,b) showing that the msl+ genes act during early development and in larval (polytene) cells, is consistent with the notion that this gene acts continuously and in most, if not all, tissues of the male. In this regard, the msl's may be analogous to other major regulatory genes (i.e., home- otic loci), such as those controlling sex determination, which act continuously and in many different tissues to control sexually dimorphic gene expression
(BAKER and RIDGE 1 9 8 0 ; BELOTE and BAKER 1 9 8 2 ) .
I wish to thank BRUCE BAKER and JOHN LUCCHESI, for their valuable comments during the preparation of this manuscript, and ROB DENELL, for suggestions on the mapping of msl-2. Re-
search supported by Public Health Service grants GM-07642 and GM-23345.
LITERATURE CITED
ASHBURNER, M. and V. VELISSARIOU, 1980 BAKER, B. S. and K . RIDGE, 1980 BECKER, H. J., 1976
BELOIE, J . M. and B. S. BAKER, 1982
New mutants. Drosophila Inform. Serv. 55: 196. Sex and the single cell: on the action of major loci affecting Mitotic recombination. pp. 1020-1084. In The Genetics and Biology ofDroso-
Sex determination in Drosophila melanogaster: analysis of sex determination in Drosophila melanogaster. Genetics 94: 383-423.
phila, Edited by M. ASHBURNER and E. NOVITSKI, Vol. IC. Academic Press, New York
BELOTE, J. M. and J. C. LUCCHESI, 1980a Male-specific lethal mutations of Drosophila melanogaster.
BELOTE, J. M. and J. C. LUCCHESI, 1980b Control of X chromosome transcription by the maleless
BRYANT, P. J. and M. ZORNETZER, 1973 Mosaic analysis of lethal mutations in Drosophila. Ge-
CLINE, T. W., 1976 A sex-specific temperature-sensitive maternal effect of the daughterless mu-
CLINE, T. W., 1978 Two closely linked mutations in Drosophila melanogaster that are lethal to
CLINE, T. W., 1979a A male-specific mutation in Drosophila melanogaster that transforms sex. Dev.
CLINE, T. W., 197913 A product of the maternally-influenced sex-lethal gene determines sex in
CLINE, T. W., 1980 Maternal and zygotic sex-specific gene interactions in Drosophila melanogaster.
CLINE, T. W., 1983 The interaction between daughterless and Sex-lethal in triploids: a lethal sex-transforming maternal effect linking sex determination and dosage compensation in Dro- sophila melanagaster. Dev. Biol. 95: 260-274.
FERRUS, A., 1975 Parameters of mitotic recombination in Minute mutants of Drosophila melano- gaster. Genetics 7 9 589-599.
FUKUNAGA, A., A. TANAKA and K. OISHI, 1975 Maleless, a recessive autosomal mutant of Dro- sophila melanagaster that specifically kill male zygotes. Genetics 81: 135-141.
GARCIA-BELLIDO, A., 1972 Some parameters of mitotic recombination in Drosophila melanogaster.
Mol. Gen. Genet. 115: 54-72.
GARCIA-BELLIW, A. and J. DAPENA, 1974 Induction, detection, and characterization of cell dif- ferentiation mutants in Drosophila. Mol. Gen. Genet. 128: 117-130.
GARCIA-BELLIW, A. and J. R. MERRIAM, 1971 Clonal parameters of tergite development in
Drosophila. Dev. Biol. 2 6 264-276.
HERSKOWITZ, I. H. and A. SCHALET, 1957 Induced changes in female germ cells of Drosophila.
V. The contribution of half-translocation and nondisjunction to the dominant lethality induced by x-raying oocytes. Genetics 42: 649-660.
LAKHOTIA, S. C. and A. S. MUKHERJEE, 1969 Chromosomal basis of dosage compensation in
Drosophila. I. Cellular autonomy of hyperactivity of the male X chromosome in salivary gland and sex differentiation. Genet. Res. 1 4 137-1 50.
LINDSLEY, D. L. and E. H. GRELL, 1968 Genetic variations of Drosophzla melanagaster. Carnegie Inst. Wash. Publ. 627.
LINDSLEY, D. L., L. SANDLER, B. S. BAKER, A. T. C. CARPENTER, R. F. DENELL, J. C. HALL, P.
A. JACOBS, G. L. GABOR MIKLOS, B. K. DAVIS, R. C. GETHMANN, R. W. HARDY, A. HESSLER, S. M. MILLER, H. NOZAWA, D. M. PARRY and M. COULD-SOMERO, 1972 Segmental aneuplo- idy and the genetic gross structure of the Drosophila genome. Genetics 71: 157-184.
Dosage compensation: transcription level regulation of X-linked genes in
Drosophila. Am. Zool. 17: 685-693.
The relationship between gene dosage, gene expression, and sex in Dro- sophila. Dev. Genet. In press.
The link between dosage compensation and sex differ- entiation in Drosophila melanogaster. Chromosoma 82: 21 7-227.
Genetics 9 6 165-186.
gene in Drosophila. Nature 285: 573-575.
netics 75: 623-637.
tation of Drosophila melanogaster. Genetics 8 4 723-742.
opposite sexes and interact with daughterless. Genetics 9 0 683-698.
Biol. 72: 266-275.
Drosophila melanogaster. Genetics 91 (Suppl.): s22.
Genetics 9 6 903-926.
LUCCHESI, J. C., 1977
LUCCHESI, J. C., 1983
896 J. M. BELOTE
LUCCHESI, J. C., T. SKRIPSKY, and F. E. TAX, 1982
MORATA, G. and P. RIPOLL, 1975
A new male-specific lethal mutation in
Minutes: mutants of Drosophila autonomously affecting cell
Behavior of somatic cells homozygous for zygotic lethals in Drosophila melano- gaster. Genetics 8 6 357-376.
Clonal analysis of Sex-lethal, a gene needed for female sexual development in Drosophila melanogaster. Wilhelm Roux' Arch. 191: 21 1-214.
Dosage compensation. pp. 107-140. In The Genetics and
Biology of Drosophila, Edited by M. ASHBURNER and T . R. F. WRIGHT, Vol. 2d. New York, Academic Press.
Somatic crossing over and segregation in Drosophila melanogaster. Genetics 21:
Quick preparation of Drosophila for microscopic analysis. Drosophila Inform.
Studies on the sex-specific lethals of Drosophila melanogrrster: further studies on a male-specific lethal gene, maleless. Genetics 84: 257-266.
Analysis of gynandromorph survivals in
Drosophila melanogaster infected with the male-killing SR organisms. Genetics 89: '7 1 1-721. Studies on the sex-specific lethals of Drosophila melamgaster. 111. A third chromosome male-specific lethal mutant. Jpn. J. Genet. 56: 523- 527.
UENOYAMA, T., S. UCHIDA, A. FUKUNAGA and K. OISHI, 1982 Studies on the sex-specific lethals of Drosophila melanogaster. 1V. Gynandromorph analysis of three male-specific lethals, mle, msl- 2", and mle(3)132. Genetics 102: 223-231.
WRIGHT, T . R. F., W. BEERMANN, J. L. MARSH, C. P. BISHOP, R. STEWARD, B. C. BLACK, A. D. T h e genetics of dopa decarboxylase in Drosophila melan-
Corresponding editor: A. CHOVNICK
Drosophila melanogaster. Genetics lOO(Suppl.): s42.
division rate. Dev. Biol. 42: 21 1-221. RIPOLL, P., 1977
SANCHEZ, L. and R. NOTHIGER, 1982
STEWART, B. and J. MERRIAM, 1980
STERN, C., 1936 625-730. SZABAD, J., 1978
TANAKA, A., A. FUKUNAGA and K. OISHI, 1976
TSUCHIYAMA, S., B. SAKACUCHI and K. OISHI, 1978 Serv. 53: 2 15.
UCHIDA, S., T . UENOYAMA and K. OISHI, 1981
TOMSETT and E. Y. WRIGHT, 1981