INTERACTIONS BETWEEN FUSED AND ENGRAILED, TWO MUTATIONS AFFECTING PATTERN FORMATION IN
DROSOPHILA M E L A N O G A S T E R
ANNE FAUSTO-STERLING A N D HEIDI SMITH-SCHIESSI
Brown University, Division of Biology and Medicine, Prouidence, R.I. 02912
Manuscript received September 3, 1981 Revised copy accepted February 5,1982
ABSTRACT
In this paper we demonstrate that the severity of the engrailed phenotype is greatly increased when engruiled is combined with the X-linked mutation fused. The secondary sex combs of fu;en flies contain from two to four times more setae than do those of en siblings. The number of transverse rows of bristles on the metatarsus of the metathoracic leg is reduced by a factor of one and a half to two in f q e n flies when compared to their e.n siblings, while the frequency of reversed bract polarity, bristle abnormalities and misshapen metatarsi increases greatly. In addition, f u ; m flies express the e n wing pheno- type more completely than their siblings and have a much higher frequency of wing vein abnormalities-some of which we have interpreted as triplica- tions of the third and first longitudinal wing veins. We briefly discuss the sig- nificance of the fused-engruiled gene interaction.
HE mutation fused ( f u ) (located at 59.5 on the
X
chromosome) and the Tmutation engrailed ( e n ) (located at 62 on the second chromosome) both affect pattern formation. The f u mutation alters or prevents the formation of wing veins 3 and 4 (FAUSTO-STERLING and HSIEH 1978; WURST and HANRATTY 1979), and a recent article (NUSSLEIN-VOLHARD and WIESCHAUS 1980) reports that lethal fused embryos (derived from homozygous fused mothers) have altered larval segment polarity. The engrailed mutation is a controlling element in the formation of the posterior compartments of adult head and thoracic structures(LAWRENCE and MORATA 1976; GARCIA-BELLIDO and SANTAMARIA 1972; MORATA
and LAWRENCE 1975; LAWRENCE and MORATA 1978). In addition, NUSSLEIN- VOLHARD and WIESCHAUS (1980) cite work indicating that en causes the de- letion of the posterior portion of alternating segements in lethal en embryos.
KORNBERG
(1981) describes a more complex pattern of segment abnormalities in lethal en embryos. In the course of experiments designed to examine compart- ment formation in fused wings (FAUSTO-STERLING and HSIEH 1978), we constructed the double mutant fused;engraiZed; in such flies we observed an en- hancement of the expression of engrailed traits. In this paper, w-e detail the pheno-1 Work submitted in partla1 fulfillment of requirements for a Master's Degree in the Division of Biology and
Medicine, Riown University
type of the fu;en fly and discuss the possible significance of this genetic interaction.
MATERIALS A N D METHODS
All flies were grown on a standard sugar-agar-cornmeal-yeast medium at 26". Eclosed adults were preserved in 100% ethanol until mounting. The wings were mounted on slides in per- mount; then the remainder of the fly was boiled in 10% KOH and mounted between 2 cover- slips in FAURE'S medium.
Stocks: Three alleles of fused-ju519, f u 5 9 , andfusgf3-were balanced over C1B (LINDSLEY
and GRELL 1968). All three stocks were obtained originally from R. C. KING. I n addition an outcrossed stock of the most severe fused allele studied here (fu59) was constructed by nine gen-
erations of backcrosses allowing free recombination to the wild type stock Swedish C. The out- crossed stock thus obtained was termed out-X-fu59. The engrailed mutant was balanced over the multiply-inverted chromosome SM5 (LINDSLEY and GRELL 1968).
In preliminary experiments crosses were made between stocks in which the en chromosome also carried the marker cn (cinnabar). The double homozygotes fu/fu;cn en/cn en and fu/Y;cn en/cn en were distinguishable on the basis of eye color (homozygous cn) as well as on the basis
of non-thoracic fused phenotypes such as the absence of ocelli. After studying such homozygotes we were convinced that the homozygous fu;en phenotype was uniquely identifiable, and we conducted all subsequent experiments on an engrailed stock which we outcrossed to remove cn.
The following crosses were then used t o obtain fu;en flies: fu/ClB female virgins were mated to +/Y;en/SMS males. From the offspring, +/fu;+/en females and f u / Y ; + / e n males were selected and mated to each other. The fu en double homozygote lacks ocelli and has a unique wing phenotype which is neither jused nor engrailed.
RESULTS
In order to examine the effect of interactions between fused and engrailed on the development of the engrailed phenotype, we quantitated particular pheno- typic effects i n siblings of the following genotypes: fu/Y;en/en, f u / Y ; f / + (01
+ / e n ) , and +Y;+/+ (or + / e n ) . Each of the three alleles of fused and the out crossed allele of fu5$ were SO examined.
Formatiorr of Secondary Sex Comb: BRASTED (1941) and TOKUNAGA (1961) provide data on the size and variability of primary and secondary sex combs in
engrailed flies. Current work interprets the secopdary sex comb, which appears in mirror image to the primary, as evidence for the transformation of the posterior compartment of the first leg imaginal disc into a mirror image of the anterior compartment ( GARCIA-BELLIDO and SANTAMARIA 1972). The data pre- sented in Table 1 show the effects of fused and engrailed on this characteristic aspect of the en mutation. In our experiments the average secondary sex comb i n homozygous en flies contained two or three setae, a size consistent with the reports of BRASTED ( 1 941 ) and TOKUNAGA (1961). The data in Table 1 indicate that males with the fused genotype often form additional sex comb setae, but these secondary setae never appear in mirror image to the primary ones. The frequency of appearance and the size of these extra setae is the same in f u / Y ;
+/SM5 and f u / Y ; + / e n males (cross not shown), a fact which suggests that extra setae are an additional aspect of the fused phenotype.
PATTERN FORMATION I N DROSOPHILA 73
TABLE 1
Effect of fused and engrailed on 2" sex comb formation
.
?number setae (SD) Individuals with
Secondary sex combs' 1' sex comb 2' sex comb 99% (65/66)
68% (41/60)
W % (IO/*) 4% ( 2/52)
100% (68/68) 76% (48/63)
37% (26/71) 0% ( W68)
95% (62/65) 72% (50/69)
9% ( 6/68)
0% ( 0/53) 92% (58/63) 63% (443/64) 12% ( 7/60) 5% ( 3/56)
12.4 (1.4) 11.3 (1.1) 11.3 (0.9) 12.3 (1.4) 13.3 (1.5) 12.2 (2.2) 11.1 (1.1) 11.1 (1.1)
11.2 (1.0)
9.9 (0.9) 10.6 (1.0) 11.0 (1.4) 11.8 (1.3) 11.5 (1.6) 12.1 (1.2) 11.2 (1.1) 11.9 (3.2) 2.8 (1.7) 1.2 (0.6) 2.5 (2.1)
9.1 (2.9) 2.8 (1.5) 1.7 (1.2)
0
10.8 (4.2) 4.0 (2.4) 7.0 (4.2)
0
7.7 (3.9)
2.0 (1.2) 2.0 (1.0) 3.6 (3.2)
* Secondary sex combs were mirrm-image i n engrailed and fused;engrailed flies but were direct in fused and wild-type individuals.
greater (depending upon the fu allele and the genetic background) than in en
alone. Figure 1 shows photographs of fu, en, and fu;en legs. The larger secondary sex combs are disorganized compared to smaller ones, a fact already noted by
BRASTED
(1941). In fact, the difference in secondary sex comb phenotvpe infu;en and en flies is sufficiently striking that one could (although we did not)
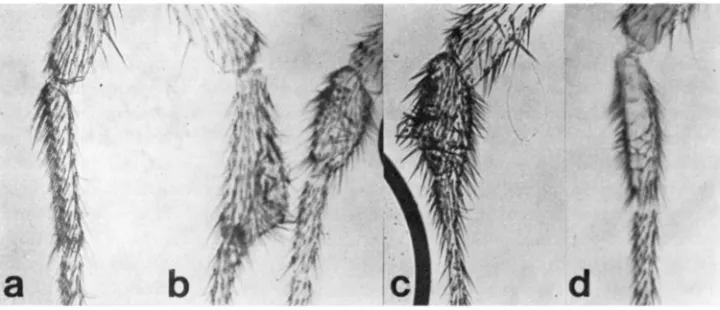
distinguish between the two genotypes solely on the basis of this phenotype. Effects of fu and en o n the development of the third (metathoracic) leg: Both engrailed and fused; engrailed flies show a variety of deformities o n the third leg. These include metatarsal abaormalities increasing in severity from distal puffi- ness to large distal triangular outgrowths (Figure 2b), o r small, twisted out- growths (Figure 2c). The more extremely deformed flies often had metatarsi which were shorter in length by at least one third (Figure 2b). Legs from fu;en frequently showed bald or whorled patches (Figure 2d), while both en and en;fu flies showed a high frequency of reversed bract polarity (Figure 2d) and incom- plete metatarsal segmentation. Figure 2a is a photograph of the metatarsus of the third leg of a phenotypically wild-type fly. The frequencies of the above-described abnormalities in fused, en, wild type, and fu;en flies are given in Table 2. With the exception of the allele fuSefs, for which there is no difference between en
and fu;en siblings, the double mutant combination shows a two and a half to five
FIGURE 1.-A. Primary and secondary sex combs of a leg from a fly of the genotype fubzfs/Y; en/en. “s” points to the secondary sex comb which contains a larger number of setae than the
primary comb. B. Primary sex comb and extra setae (“e”) of a leg from a fly of the genotype f u s z f s / Y ; + / e n (or +/+). C. Secondary sex comb (“s”) of a leg from a fly of the genotype + / Y ; en/en. The primary sex comb is out of the plane of focus.
on them, in comparison to only infrequent occurrence in en flies. Similarly. the frequency of flies in which the metatarsus of the metathoracic leg is shorter than normal increases only in the double mutants. And finally, (again with the ex- ception of f u b 2 f s ) , there is a substantial increase in the frequency of abnormalities in metatarsal shape (from two to four times) in the double mutants as compared to their en siblings.
Table 2 shows the effect of f u and en on the number of transverse bristle rows in the metatarsus of the third leg. As can be seen from the data, there is a marked reduction in tho number of transverse rows in fu;en flies (many of which have
FIGURE 2.-A. Metatarsus of the third leg of a phenotypically wild-type fly. B, C, and D.
PATTERN FORMATION IN DROSOPHILA 75
S
N O 0 0 O O M O M r O O O m o o 0
n a a a
x m o o m m m o O C O M O o o c o o
- -
c o + w o o m o lglnng ~ M c o O h " O o o c o o - m m 0 m - c b o a m m s
i CI
D O 0 0 M O O 0 W M O O o m 0 0
A. FAUSTO-STERLING A N D H. SMITH-SCHIESS
none at all). Frequently, the proximal portion o l the metatarsus in such flies is quite normal, while crowding of bristles, disorganization of rows, and reversal of bract polarity are striking at the distal end. If the transverse rows lie in the pos- terior compartment of the metathoracic leg, as one might infer from compart- mental lines on other legs (STEINER 1976) as well as from their location (POODRY
and SCHNIEDERMAN 1976), then a reduction in the number of transverse rows might well be expected in cases where there is a strong tendency to transform the posterior into the anterior compartment. One interpretation of our data is that en alone results in a transformation too weak to cause a reduction in the number of transverse rows, but that i n combination with fused the transforma- tion in the leg compartment is much stronger.
Effects of the fu;en combination o n wing development: Figure 3 shows dia- grams of wild type (3a), engrailed (3b), and fused (3c) wings. The drawings in- dicate the location of the anterior-posterior compartment border as well as the following diagnostic anatomical features: sensillae (duplicated in en wings) on longitudinal veins I (LV I) and I11 (LV 111), the dorsal protrusion of veins 111
and IV, the ventral protrusion of longitudinal veins I, I1 and IV, and the presence of a triple row of bristles along the anterior edge of the wing (duplicated in en
wings). As shown in Figure 3c, fused wings often contain an extra sensilla along LV 111. Veins in the posterior Compartment of fused;engrailed wings were clas- sified on the basis of their dorsal or ventral protrusion and the presence of sensil- lae. LAWRENCE and MORATA (1976) suggest that the engrailed phenotype does not always fit neatly into the scheme suggested by GARCIA-BELLIDO and SANTA-
MARIA (1972). Although in the following interpretation we attempt to follow the
latter’s scheme, we are aware that we may at times be dealing with a unique phenotype rather than straight-forward duplication of the structures of the anterior wing compartment.
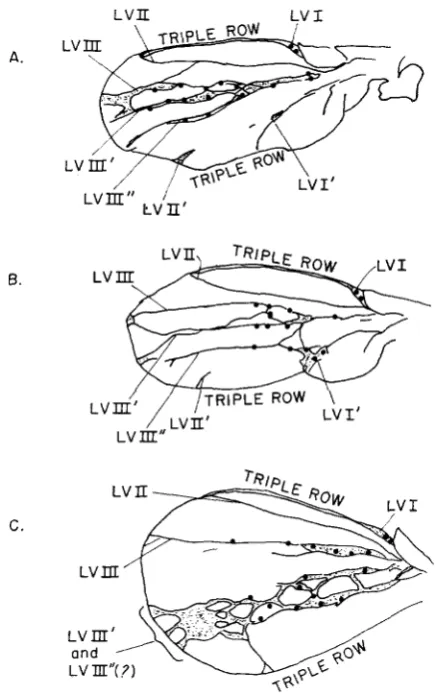
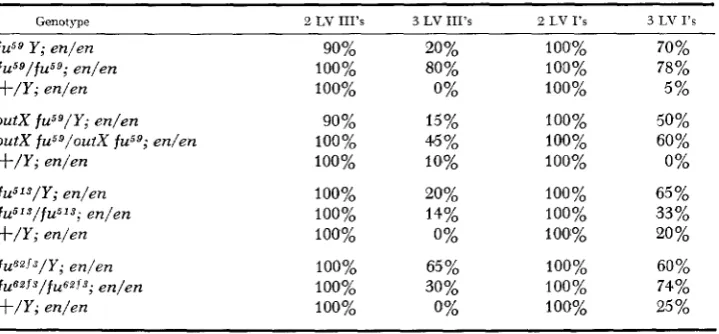
I n the course of analyzing vein formation in fu;en flies, it became apparent that some wings show morphological evidence for triplication of LV I and LV 111. Drawings of three such wings appear in Figure 4. In the wing illustrated in Figure 4c a vein plexus replaces any distinct LV I1 duplication, and evidence for LV 111’ multiples comes from the formation of a n extra row of dorsal sensillae. Table 3 shows the frequency of multiple vein formation, a frequency consider- ably elevated in all of the fu;en combination. I n addition t o the complex vein phenotype, we observed that LV I’ formed completely in fu;en flies with a higher frequency than for en alone. I n en flies LV I’ was complete in from 3% (in the cross with o u t - X - f ~ ~ ~ ) to 50% (in the cross with fuGgfls) of the cases while the frequency of completely formed LV I’ in fu;en flies was 57% for fu5”, 85% f o r
f u S g , 93% for fuGZfsand 98% f o r ~ u t - X - f u ~ ~ .
DISCUSSION
The data presented in this paper show the following:
PATTEKN FORMATION IN DROSOPHILA 77
A. A
PO
B.
A
C.
ANT
\
P O S T
FIGURE 3.-A. Drawing of a wild type wing showing the anterior (“ant”)/posterior (“post”) compartment border, the dorsal (D) o r ventral (V) protrusion of the longitudinal wing veins
(LV I through LV V), as well as the location of sensillae (heavy black dots) along the veins. B. Drawing of an engraikd wing showing the transformation of the posterior compartment into a secondary anterior compartment (ANT’), the dorsal or ventral protrusion of longitudinal veins I through I11 (LV I-LV 111) as well as the location of duplicated veins (LV’) and s e n d - lae. C. A drawing of a fused wing showing the location of the anterior/posterior compartment border. All abbreviations as in 3A.
currence of secondary sex combs rises from between 46%-86% in en to 92%-
100% in fu;en brothers.
2) The number of metatarsal transverse rows in the metathoracic leg is reduced by a factor of one-and-a-half to two in fu;en flies compared to their en siblings, while the frequency of reversed bract polarity, bristle abnormalities, and mis- shapen metarsi increases greatly.
A.
B.
C.
L V I I L V I
5
L V
L V
LV"
FIGURE 4.-A, B, and C all show camera lucida tracings of the vein pstterns and location of sensillae (heavy black dots) of three different wings from fused;engrailed flies. A' indicates a duplicated structure while a" suggests a triplicated one. LV = longitudinal wing vein.
some of which we have interpreted as triplications of the third and first longi- tudinal veins.
W e view these data as evidence that the X-linked mutation fused enhances the expression of the second chromosome mutation engrailed. The data on increased size and frequency of secondary sex combs are straightforward. Those on altera- tions in the metatarsus of the third leg require some interpretation. The work of
POODRY
and SCHNEIDERMAN ( 1976) locates the metathoracic, metatarsal trans-P A T T E R N F O R M A T I O N IN DROSOPHILA 79
TABLE 3
Frequency of occurrence of triplications of partial or completely formed LV I l l and LV I
GenotJQe 2 LV 111’s 3 LV 111’s 2 LV 1’s 3 LV 1’s
fu59 Y; en/en fu59/fu59; en/en
+/Y; en/en
outX fuS9/Y; en/en outX fu59/outX fu59; en/en
+/Y; en/en
fu513/Y; en/en f u 5 I ~ / f u 5 1 ~ ; en/en
+/Y; en/en
fuBZJ$/Y; en/en
fu@2fs/fu@zf$; en/en
+/Y; en/en
90% 100%
looyo
90% 100% 100% 100% 100% 100% 100% 100% 100% 20% 80% 0% 15% 45% 10% 20% 14% 0%65 % 30% 0% 100% 100% 100% 100% 100% 100% 100% 100% 100% 100% 100% 100% 70% 78% 5% 50% 60% 0%
65 % 33% 20 % 60%
74%
25 %
n = 39 to 41 for each genotype.
the anterior developmental compartment. Whatever the anatomical interpreta- tion, the effect on leg development is more severe in the double mutants than in
en alone.
In analyzing wing vein formation, we classified extra veins on the basis of their dorsal or ventral protrusion and on the presence o r absence of sensillae
(LAWRENCE
andMORATA
19 76; GARCIA-BELLIDO and SANTAMARIA 19 72). The presence of rows of extra sensillae were of particular importance in the interpre- tation of vein triplications. The interpretation of complex vein patterns such as those found in fu;en wings will always be open to discussion. In looking at some wings we would agree with LAWRENCE and MORATA (1976) that the phenotype is unique; however, many of the wings from fu;en flies show what seems to us to be clear evidence for triplication of anterior compartment veins, especially of LV111. As
with
other phenotypes, the complex vein formation-however one inter- prets it-occurs far more frequently in fu;en than in en siblings. WHITTLE(1976) reported that the costal mutation causes triplications and even quadrupli- cations in the anterior wing compartment and suggested that such complex patterns may result from regeneration following cell death, an interpretation which could as well apply to our results. Similar findings exist for leg imaginal discs (GIRTON 1981).
We do not yet know why the genes fused and engrailed interact. In lethal fused
requires for survival. Similarly, KORNBERG (1981) has presented the most recent evidence that the engrailed function is required only in posterior compartment cells. The fact that these two genes interact suggests that they may contribute important, albeit different, products to the same developmental pathway, one necessary both f o r maintaining the integrity of the posterior compartment and for proper segment formation in the embryo.
L I T E R A T U R E C I T E D
BRASTED, A., 1941
FAUSTO-STERLING, A. and L. HSIEH, 1978 GARCIA-BELLIW, A. and P. SANTAMARIA, 1972 GIRTON, J. R., 1981
KORNBERG, T., 1981
LAWRENCE, P. and G. MORATA, 1976
An analysis of the expression of the mutant engrailed i n Drosophila melano-
Pattern formation in the wing veins of the fused Developmental analysis of the wing disc in the Pattern triplications produced by a cell lethal mutation in Drosophila. engrailed: A gene controlling compartment and segment formation in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 78: 1095-1099.
Compartments in the wing of Drosophila: A study of the engrailed gene. Dev. Biol. 50: 321-337. __ , 1978 Major genes which control de- velopmental pathways in Drosophila. Abst. Int. Conf. on Molec. and Develop. Biol. of Insects p. 98.
LINDSLEY, D. and E. GRELL, 1968 Genetic variations of Drosophila melanogasier. Carnegie Inst. Wash. Publ. 627.
MORATA, G. and P. LAWRENCE, 1975 Control of compartment development by the engrailed gene in Drosophila. Nature 255 : 614-61 7.
NUSSLEIN-VOLHARD, C. and E. WIESCHAUS, 1980 Mutations affecting segment number and polarity in Drosophila. Nature 287: 795-801.
POODRY, C. and H. SCHNEIDERMAN, 1976 Pattern formation i n Drosophila melanogaster: the effects of mutations on polarity i n the developing leg. Wilhelm Roux Arch. 180: 175-188. STEINER, E., 1976 Establishment of compartments in the developing leg imaginal discs of
Drcsophila melanogaster. Wilhelm Roux Arch. 180: 9-30.
TOKUNAGA, C., 1961 The differentiation of a secondary sex comb under the influence of the gene engrailed in Drosophila melanogaster. Genetics 46: 157-1 76.
WHITTLE, J. R., 1976 Clonal analysis of a genetically caused duplication of the anterior wing in Drosophila melanogasier. Dev. Biol. 51: 257-268.
WURST, G. and W. HANRATTY, 1979 Studies on the developmental characteristics of fused mutants of Drosophila melanogaster. Can. J. Genet. Cytol. 21 : 335-346.
Corresponding editor: A. T. C. C~RPENTER gasier. Genetics 27: 347-373.