CHROMOSOME SEGREGATION MECHANISMS’
R. BRUCE NICKLAS
Department of Zoology, Duke University, Durham, North Carolina 27706
ABSTRACT
Most aspects of chromosome distribution to the daughter cells i n meiosis and mitosis are now understood, at the cellular level. The most striking evidence that the proposed explanation is valid is that it correctly predicts the outcome of experiments on living cells in which the experimenter ( 1 ) can determine the distribution of any chosen chromosome to a chosen daughter cell, (2) can induce a mal-orientation, and (3) can stabilize a mal-orientation, causing non- disjunction of a chosen bivalent. Recent reviews of chromoscrme distribution mechanisms are also considered, in an attempt to clarify the remaining un- solved problems.
INHERITANCE depends upon the equal distribution of chromosomes to the daughter cells in mitosis and upon the orderly segregation of chromosomes to the gametic nuclei in meiosis. It was asserted in a recent review that controlled distribution and segregation are now probably understood at the cellular level (NICKLAS 1971). Little that is novel can be added here, but perhaps an outline with diagrams will be useful, covering the proposed explanation and the most striking evidence-experimental control of chromosome segregation. Additional discussion and literature references will be found in the review.
Meiotic bivalents have been the chief objects of study, so their behavior will be illustrated followed by comments on other meiotic units and on mitosis. Finally, the conceptions of chromosome distribution presented in two other recent reviews
(LUYKX 1970; BAJER and M O L ~ B A J E R 1972) will be considered.
MEIOTIC BIVALENTS
Two principles prescribe exactly what must be explained if controlled chromo- some distribution is to be understood. The first is “kinetochore orientation”: as diagrammed at the left in Figure 1, each half-bivalent becomes associated with a particular spindle pole via chromosomal spindle fibers attached to a defined chro- mosome locus-the kinetochore or centromere. For example, half-bivalent “a” is said to be oriented to pole 1. And kinetochore orientation, occurring soon after formation of the spindle, determines chromosome distribution in anaphase-in our example, half-bivalent “a” moves to pole 1 (Figure 1, right), to be included in the daughter nucleus formed there. A corollary is that the usual equal distribu-
The investigations in the author’s laboratory were supported in part by grant GM-13746 from the Division of General Medical Sciences, Public Health Service. This article was written at the Max-Plan&-Institute fiir Biologie, Abt. BeermaM, Tubingen, Federal Republic of Germany, while the author held a John Simon Gnggenheim Memorial Fellowship in this stimulating and hospitable atmosphere. Special thanks are due Mn. ERICK FnIEBERG for the drawings.
206 R. B. NICKLAS
Background
,I \
I
I \ I \
\ \
I
‘
p2MAL-ORIENTATION
,
I ,
I \ I
I
I \
I 1
I 1
\
,’
Pl \I I
I I
IS
BY
FOLLOWED RE-ORIENTATION
FIGURES 1 and 2.--Diagrams illustrating what must be explained: kinetochore orientation (Figure 1) and reorientation (Figure 2).
Explanation
\ i
\
‘
p2’
PREFERENTIAL CHROMOSOME
OR I EN TAT ION STRUCTURE
8
FORCES TOWARD FORCES TOWARD
OPPOSITE POLES VERSUS ONLY ONE POLE
-TENSION - -NO TENSION-
@
FIGURES 3 and 4.-Diagrams illustrating the proposed explanation of initial Orientation (Fig- ure 3) and of reorientation (Figure 4). The arrows and numbers in Figure 3 indicate the fre- quency of orientation of the upper half-bivalent to the upper uersus the lower pole. See text.
208 R. B. NICKLAS
organization to face in opposite directions and if preferential orientation to the “faced” pole is a fact, then partner half-bivalents must tend to orient to opposite poles. This is one portion of OSTERGREN’S ( 195 1 ) orientation theory.
Second, the reorientation of initially mal-oriented bivalents must be explained.
DIETZ
(1958) suggested that any given reorientation does not certainly lead to a bipolar orientation, but rather, orientation occurs and recurs until the bipolar orientation is achieved because only this orientation is stable. As observed in living cells, any mal-orientation is soon followed by reorientation while the bi- polar orientation whenever achieved, almost always persists unaltered through anaphase. This reveals the proper question: what causes differential orientation stability? The proposal is diagrammed in Figure 4: tension forces confer stability, and only in the bipolar orientation (left) do the normal spindle forces act i n opposite directions, producing tension; in the absence of tension (e.g., as at the right), any orientation is unstable, that is, orientation is likely. The required difference in spindle forces certainly exists as evidenced by a slight stretching ofbivalents when and only when the bipolar orientation has been attained. This explanation was tentatively advanced by DIETZ (1958).
Experimental Tests
Micromanipulation of bivalents in living insect spermatocytes has made possi- ble very direct tests of possible distribution mechanisms. Spermatocytes from
Melanoplus differentialis, a grasshopper, have usually been used, but the methods have also been applied successfully to several other materials. Numerical values given below are for Melanoplus spermatocytes.
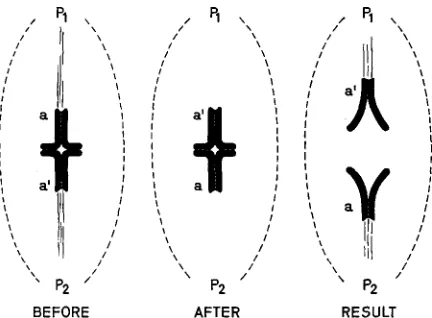
Znitial orientation: Bivalents can be detached from the spindle and placed where desired within the cell by using a fine glass micromanipulation needle. “Detached” is a n appropriate word: the pre-existing kinetochore-pole associa- tions have been broken. Preferential orientation was readily tested by placing detached bivalents at a variety of aogles to the spindle: 84% ( n = 25) oriented initially to the pole one half-bivalent’s kinetochores most nearly faced (see Figure 3, left; (NICKLAS 1967). This unambiguously verifies the proposal for initial orientation because the required feature of chromosome structure is established beyond doubt for most bivalents (and where absent, as in long, flexible bivalents, the frequency of successful initial orientation is decreased, just as expected; review: NICKLAS, 1971, p. 261). The explanation predicts that orientation, and hence distribution, can be determined experimentally, given that the experiment is possible. I t is: a bivalent with the orientation diagrammed in Figure 5,
/ p 1
I
I I
\ 1
\ \
\ \
p2
\ \
\ \
\ \
I
I
I I
I I I
/
I
/ /
BEFORE AFTER RESULT
FIGURE 5.-Diagrams illustrating the experimental control of chromosome distribution. See text.
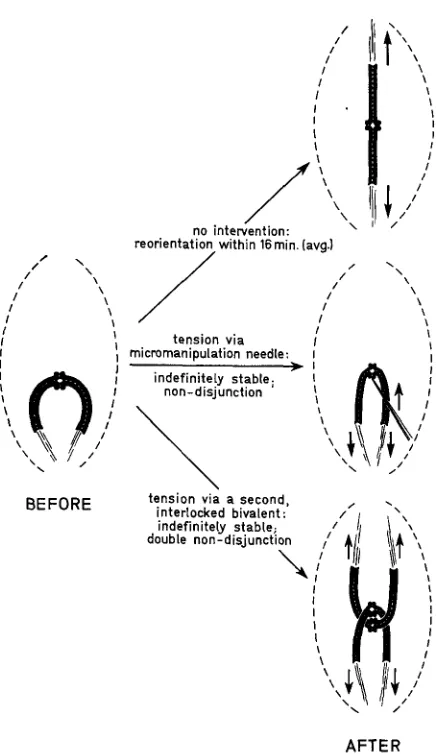
Reorientation: The required experiment is obvious: if spindle fiber tension stabilizes bipolar orientations, then application of tension by artificial means should stabilize normally unstable orientations. Such an unstable orientation is diagrammed in Figure 6 (“Before”) together with various outcomes (“After”). If there is no intervention (uppermost “After” diagram)
,
reorientation rapidly follows (rapidly relative to the six hours between spindle formation and anaphase in this material). But application of tension in either of two ways (lower two diagrams) makes the mal-orientation stable indefinitely-until termination of the experiment or until anaphase (NICKLAS and KOCH 1969;HENDERSON
and KOCH 1970). In the latter case, nondisjunction occurs, experimentally induced in a chosen bivalent or bivalents. Two additional features of these experiments should be noted. First, whenever the artificially applied tension is relaxed for several minutes, reorientation almost always follows: the artificially stabilized mal-orientation is again unstable. Second, the mal-orientations were induced experimentally by a process demonstrating again a preferential orientation in the initial orientation process. Thus when a detached bivalent is bent with the micro- manipulation needle so that the kinetochores of both partcer half-bivalents face the sane pole, both usually orient to that pole, producing the configuration shown at the left in Figure 6. Interlocking, mal-oriented bivalents have also been observed without micromanipulation, following a heat shock during meiotic pro- phase; these too achieve a n apparently stable orientation and undergo double nondisjunction in anaphase (Buss andHENDERSON
1971). It should be noted, however, that all these experiments are formally identical, and the proposed explanation of reorientation is not established beyond doubt.COMMENTS
21 0 R. B. NICKLAS
/' '\
/ \
I \
I
I
\
\
I 1,
I
I I
I
I I
\ \
1
I I
I
I I I I
\
\ I
\ /
I
tension v i a I 1 I
I
I I I
micromanipulation needle: I I
* I
indefinitely stable; I
1
non-disjunction
1
\ I
\ I
\ I
\
\ \tension via a second,
interlocked bivalent: indefinitely stable; double non-disjunction BEFORE
A F T E R
FIGURE 6.-Diagrams illustrating mal-orientation (left) and its consequences (right) without experimental intervention (uppermost diagram) and with experimental intervention (lower two diagrams). The arrows indicate the direction of tension forces.
As for chromosome distribution in somatic mitosis and the second meiotic division, all the prerequisites for initial orientation and reorientation considered above are present here as well, but decisive observations are lacking because orientation patterns in mitosis are often unclear in living cells. Nevertheless, initial orientation very probably results in successful orientation of sister chroma- tids to opposite pole by the same preferential orientation-kinetochore placement (“chromosome structure”) rules established for bivalents. It is now seen as in- creasingly likely, however, that reorientation plays a quantitatively minor role. Thus recent electron microscopic observations suggest that delayed orientation of one sister kinetochore is far more frequent than mal-orientation of both kineto- chores to the same pole (Roos 1973; MOL&BAJER and
BASER,
in preparation). The possibility of delayed orientation was anticipated, and its potential for facili- tating appropriate bipolar orientation was realized, byOSTERGREN
long ago (1951, especially pp. 146-148; see also NICKLAS 1971, pp. 264 and 268). Nothing is known about the causes of delayed orientation, of its restriction to one chroma- tid‘s kinetochore, or of the eventual Orientation that follows.The reviews of LUYKX (1970) and of BAJER and MOL~-BAJER (1972) provide a fresh stimulus to considering some unanswered questions about chromosome distribution. The basic observations, experiments, and the principles of initial orientation as given here have not been disputed, but the interpretation of later events is not yet resolved. A basic problem is the extent to which deterministic as opposed to statistical processes govern reorientation. For clarity the extreme views may be contrasted: all of the several possible types of mal-orientation are con- verted automatically to the one appropriate orientation by some precise, flawless mechanism (s)
,
uersus reorientation that is entirely non-specific as to outcome, simply recurring until the appropriate orientation arises entirely by chance because only the appropriate orientation is stable. I would summarize the current situation as follows. (1) Only for the statistical hypothesis(DIETZ
1958) is there any positive evidence: (a) the required difference in stability (probability of reorientation) is observed; (b) the probable origin of stability differences (pres- ence or absence of spindle fiber tension) has been successfully tested experi- mentally; and (c) bivalent reorientation as seen in living cells appears to be non-specific; at least it is highly variable in timing and in outcome-either half- bivalent or both may reorientZ (review: NICKLAS 1971, p. 264). Moreover, non- specific reorientation plus stabilization of the appropriate orientation is in princi- ple sufficient by itself tcj account for the facts. (2) Conversely, there is no direct evidence for any determinate mechanism in reorientation and no single, sufficient mechanism has been proposed. Thus the molecular model of LUYKX (1970- Figure 20 and associated text) cannot account for the reorientation of mal- oriented bivalents lying more or less stationary near one pole, and this is theirThe probability of kinetochore reorientation in crane fly spermatocytes is influenced by such factors as the length of time since the last reorientation and the distance of the kinetochore from the pole (DIETZ 1969; and unpublished obser- vations cited by BMER and MOLPBAJER 1972, p. 208). Therefore the reorientation of one particular half-bivalent in a
bivalent is often more likely than reorientation of the partner half-bivalent. However, the critical question here is whether or not the factors underlying different reorientation probabilities act as a repair mechanism, detecting and
212 R. B. NICKLAS
usual situation at the time of reorientation. The same limitation applies to OSTERGREN’S “co-orientation by pulling hypothesis” ( BAUER, DIETZ and ROBBE-
LEN 1961; NICKLAS 1967; review: BAJER and MOL~BAJER 1972, pp. 2lOff). But
directed reorientation, limited to specific situations, is not ruled out, and in one case actually seems likely. Thus electron microscopic investigations disclose fre- quent mal-orientations involving a single chromatid kinetochore in which most of the microtubules are associated with the facing pole but a few run toward the opposite pole (review: LUYKX 1970). On the model of LUYKX (1970), the latter, smaller number of microtubules would be specifically lost, while the others would remain, leading with minimal change to the appropriate oriectation. His or some similar model may well be necessary to explain rectification of this particular type of mal-orientation. (3) We are ignorant even of the events surrounding re- orientation (e.g., the changes in microtubule disposition near chromosomes dur- ing known stages in reorientation)
,
let alone their particular c a ~ s e s . ~ BAJER and MOL~-BAJER (1972 p. 215), on the contrary, conclude that reorientation is under- standable now in terms of known spindle fiber and aster transport properties. This has the great merit of properly directing our attention to these properties, rather than to properties of reorienting kinetochores them~elves.~ However, the factors involved in even one individual reorientation still remain to be identified and their connection to be observed particulars of chromosome behavior remain to be made plain. Correlated study of the same chromosome in the living cell and by electron microscopy should provide the missing facts. One misleading statement requires comment:“It
is quite probable, therefore, that contrary to the suggestion of NICKLAS, a chromosome at the pole is in fact under tension before reorienta- tion.” (BAJER and MOL~-BAJER 1972, p. 214). The “tension” to which they refer cannot be that considered by NIcKLAs-tension forces along the axis of the bivalent-because a bivalent at the pole plainly is not stretched by mitotic forces before orientation, and when stretched by artificial means, does not reorient (see Figure 6, above). Rather, as suggested by their own remarks adjacent to those quoted, they presumably mean that twisting or pushing forces on a n individual half-bivalent may be the immediate cause of reorientation. This is certainly plausible and of course it is not at all in conflict with the postulated role of tension in orientational stability.CONCLUSIONS
Chromosome distribution at mitosis and meiosis is understood at the cellular level, except for some features of reorientation. Moreover, molecular hypotheses are readily suggested, and those for initial orientation phenomena are especially
“Particular” is used here to refer to the immediate causes of a particular reorientation, as opposed to the general causal role proposed for spindle fiber tension indetermining whether or not orientation is likely to occur (for a brief statement of the problem and speculation, see NICKUS and KOCH 1969, pp. 4748).
Thus BAJER and MOLE-BAJER (p. 215) reject the idea that kinetochore reorientation is “autonomous” which pre- sumably means “due to kinetochore properties alone”. That is a good point. However, since they consistently use “autonomous reorientation’’ to refer to Dietz’ statistical conception of reorientation, apparently they reject this as well.
reasonable and testable (review: NICKLAS 1971, pp. 273ff). It must be noted, how- ever, that our understanding is limited to the majority of eukaryote chromosome systems: those with localized kinetochores and physical connections between chromosomes which regularly orient to opposite poles. Chromosome distribution in organisms with holokinetic chromosomes poses at least some special problems
(recent observations and older literature: COMINGS and OKADA 1972; OZMENT 1972). Preferential distribution of chromosomes that are not physically linked provides truly unique challenges and opportunities (reviews: LUYKX 1970; NICK-
LAS 1971).
LITERATURE CITED
BAJER, A. S. and J. MO&-BAJER, 1972 Spindle Dynamics and Chromasome Movements. Intern. Rev. Cytol. (Suppl. 3.) Edited by G. H. BOURNE and J. F. DANIELLI. Academic Press, New York.
BAUER, H., R. DIETZ and C. R~BBELEN, 1961 Die Spermatocytenteilungen der Tipuliden. 111. Das Bewegungsverhalten der Chromosomen in Translokations-heterozygoten von Tipula oleracea. Chromosoma 12: 116189.
Buss, M. E. and S. A. HENDERSON, 1971 The induction of orientational instability and bivalent interlocking at meiosis. Chromosoma 35: 153-183.
COMINGS, D. E. and T. A. OKADA, 1972 Holocentric chromosomes in Oncopeltus: kinetochore plates are present in mitosis but absent in meiosis. Chromosoma 37: 177-1952.
DIETZ, R., 1958 Multiple Geschlechchrmmmen bei den cypriden Ostracoden, ihre Evolution und ihr Teilungsverhalten. Chromosoma 9: 359-1.40. Bau und Funktion des Spindelapparats. Natunvissenschaften 56 : 237-248.
HENDERSON, S. A. and C. A. KOCH, 1970 Co-orientation stability by physical tension: a demon- stration with experimentally interlocked bivalents. Chromosoma 29 : 207-216.
HUGHES-SCHRADER, S., 1943 Polarization, kinetochore movements, and bivalent structure in the meiosis of male mantids. Biol. Bull. 85: 265-300.
LUYKX, P., 1970 Cellular Mechanisms of chromosome Distribution. Intern. Rev. Cytol. (Suppl.2.) Edited by G. H. BOURNE and J. F. DANIELLI. Academic Press, New York. MO&-BAJER, J. and A. BAJER, in preparation. Relations of chromosome movements modified
by aster activity in the newt.
NICKLAS, R. B., 1967 Chromosome micromanipulation 11. Induced reorientation and the experi- mental control of segregation in meiosis. Chromosoma 21: 17-50. Mitosis. pp. 225-297. In: Advances in Cell Biology. Vol. 2, Edited by D. M. PRESCQTT, L. GOLDSTEIN and E. MCCONKEY. Appleton-Century-Crofts, New York.
NICKLAS, R. B. and C. A. KOCH, 1969 Chromosome micromanipulation 111. Spindle fiber tension and the reorientation of mal-oriented chromosomes. J. Cell Biol. 43: W O .
OSTERGREN, G., 1951 The mechanism of co-orientation in bivalents and multivalents. Hereditas 37: 85-156.
OZMENT, N. E., 1972 Fine structure of meiosis and mitosis in Steatucoccus tuberculatus
Morrison. The structure and function of the diffuse kinetochure. Ph.D. Dissertatian, Depart- ment of Zoology, Duke University, Durham, N.C.
Light and electron microscopy of rat kangaroo cells in mitosis 11. Kinetochore structure and function. Chromosoma 41 : 195-220.
- , 1969