Copyright 0 1988 by the Genetics Society of America
Characterization of
New Punch
Mutations: Identification
of
Two
Additional
Mutant Classes
Elaine R. Reynolds and Janis
M.
O’Donnell’
Department of Biological Sciences, Carnegie Mellon University, Pittsburgh, Pennsylvania 15213 Manuscript received June 19, 1987
Revised copy accepted March 9, 1988
ABSTRACT
The Punch locus of Drosophila melanogaster which encodes the pteridine biosynthetic enzyme, GTP cyclohydrolase, is genetically complex. Lethal alleles of the locus resolve into an array of interallelic complementation groups, and at least one class of mutations is developmentally specific, affecting GTP cyclohydrolase activity only in the heads of adults. All previously isolated Punch alleles were identified on the basis of a mutant eye color phenotype. By screening mutagenized chromosomes over Punch region deficiencies, we have now isolated new alleles on the basis of lethal and visible phenotypes. Most of these alleles fall into previously identified genetic classes, but two new classes of mutations were also found. One class contains two alleles that behave as dominant lethal mutations in some genetic backgrounds. The other class represents a second developmentally specific set of alleles that affect the function of the Punch locus only during embryogenesis.
T
HE Punch (Pu) locus of Drosophila melanogaster is complex, both in terms of its genetics and mutant phenotypes (MACKAY and O’DONNELL 1983; MACKAY, REYNOLDS and O’DONNELL 1985; REYNOLDS and O’DONNELL 1987). The locus encodes the first enzyme in the pteridine biosynthetic pathway, guan- osine triphosphate cyclohydrolase (GTP CH)(MACKAY and O’DONNELL 1983; WEISBERG and O’DONNELL 1986). Since pteridines are required in Drosophila as eye pigments and as cofactors for several metabolic enzymes, numerous cellular pro- cesses should be affected by their absence (PHILLIPS and FORREST 1980; WAHL et al. 1982). Accordingly, most P u homozygotes die during embryogenesis. When heterozygous with wild-type P u alleles, these mutant alleles cause decreases in GTP CH activity in both larvae and adults. In contrast, mutants in a developmentally specific class, representing about 10% of all Pu mutants generated, have normal or nearly normal GTP CH activity in larvae and in the combined thoracic and abdominal tissues of the adults, but have less than 10% of normal GTP CH activity in adult heads. Such mutants are, therefore, severely deficient in pteridine pigments in the eyes of adults. They are homozygous viable and are ap- parently normal for the embryonic functions of the locus. These mutant alleles fully complement the vital functions of the homozygous lethal alleles, but fail to complement their eye pigmentation function
(MACKAY and O‘DONNELL 1983; MACKAY, REYNOLDS and O’DONNELL 1985).
P u lethal complementation experiments defined
Author to whom all correspondence should be addressed.
Genetics 119 609-617 (July, 1988).
multiple interallelic complementation classes, and we noted that surviving trans-heterozygotes displayed allele-specific differences in their abilities to comple- ment for the various mutant phenotypes. That is, some allele combinations that permitted only poor trans-heterozygote viability complemented almost fully for adult eye pigmentation, while others resulted in almost normal viability frequencies, but poor ability to synthesize eye pigments (MACKAY, REYNOLDS and O’DONNELL 1985).
When the phenotypes of embryos homozygous for lethal P u alleles were examined, evidence of further complexity was obtained. Embryos homozygous for noncomplementing alleles and for one class of com- plementing alleles showed a phenotype identical to the phenotype of catecholamine-deficient mutants, i.e., the cuticle is weak and normally melanized cuti- cular structures are unpigmented (REYNOLDS and O’DONNELL 1987; WRIGHT 1987; K. WHITE, personal communication). This phenotype was expected since a pteridine is required as a cofactor for catecholamine biosynthesis in other organisms (NAGATSU, LEVITT and UDENFRIED 1964; MIWA, WATANABE and HAY- AISHI 1985) and probably serves the same function in Drosophila. Embryos homozygous for any of the alleles in the remaining complementation classes dis- play the unpigmented phenotype to varying degrees and, in addition, have a new and unpredicted phe- notype, a segment pattern defect (REYNOLDS and ODONNELL 1987). Thus, there is also a suggestion of functional complexity in the embryonic lethal
Pu
mutants.mutants in our mutagenesis experiments, all of which were performed by screening for new mutants with eye color phenotypes (MACKAY and O'DONNELL 1983). Since we are interested in the organization of the P u locus and its regulation during development, it is important to define the full range of P u functions. Therefore, we turned to mutagenesis screens that required no restrictions on phenotypes. Among the P u mutants derived in these experiments are two new classes of mutants that could not have been recovered previously. One class contains alleles that confer dominant lethality under some conditions, and the other class contains alleles that perform normally in adult eyes but are incapable of providing a vital embryonic function. We describe here the characterization of alleles in these two classes, as well as other alleles that fall into previously defined classes.
MATERIALS AND METHODS
Strains: All Drosophila strains were maintained in half- pint plastic bottles at 25". Animals to be used for enzyme assays were the progeny of 10- 15 males and 20-30 females per culture bottle. Unless otherwise noted, P u mutations are as described in MACKAY and ODONNELL (1983) and MACKAY, REYNOLDS and O'DONNELL (1985). The P u alleles E I 8 a , SHC, rSH8, rK8-2, K5-2, rFAMI-2, rPF12a and rPF76 were generated in the laboratory of ROBERT BOSWELL, using either EMS or ENU as mutagens (R. BOSWELL, personal communication). Other mutations are described in LINDS-
LEY and GRELL (1968).
Mutagenesis: New Pu region mutations were obtained by EMS mutagenesis of males homozygous for second chromosomes marked with dp cn a p x sp, shu b cn bw or cn bw sp, according to the method of LEWIS and BACHER (1968). FI males carrying the mutagenized second chromosome balanced over S M l , a t Cy cn2 spz were pair-mated to females heterozygous for a deficiency of the Pu region of the second chromosome and the SM1 balancer. T h e deficiency chromosome most frequently used in these experiments was D f ( P R ) P W , a diepoxybutane-generated deletion ex- tending from approximately 57B 17-57Dll. FP progeny were screened for lethal and visible mutations.
Complementation tests: For complementation experi- ments, 10 males and 20 females were mated and maintained in half-pint bottles at 25". All new mutants were mated to representative Pu mutants in reciprocal combinations. In most cases, at least 400-500 progeny were scored.
Enzyme assays: GTP CH activity was determined as described in MACKAY and O'DONNELL (1983), with the following modifications: 60 young adult heads or climbing third instar larvae were homogenized in 400 ~1 of the appropriate homogenization buffers to prepare extracts for assay.
Egg counts: Percentages of embryonic lethality were determined as described in MACKAY, REYNOLDS and O'DON-
NELL (1985).
RESULTS
Generation and Identification of New
Pu
Alleles: T h eF2
progeny of mutagenized males were screened for the absence of the Cy+ class in deletion hetero-homozygous viable embryonic lethal
I I1
X17. ZS, 1. AA4. WM2: 331 a F36, K11. MP1 and other deficiencies
b 2, AA1, AA17, V3, V9, V14. VIS, P I . P21. W10, X 2 5 2 2 ,
PH30: 222, TR6: PD16' TR3: WMI: WM4: WM5: TR1: TRS: SHC: PJ15: E18A: SH8: KS-2: FAMI-2: PFl2.a'
V
WEST, KS-2: (WE757
ill iVa
P30, P42, JE5' S47, P I ! . Z19. WM6'
IVb IVC
P43 19. TR7: WM3: WMTPF7b'
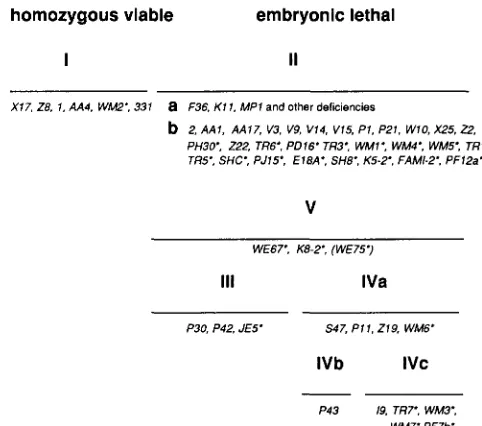
FIGURE 1.-Viability complementation map of Pu alleles. This map is based on data reported in MACKAY, REYNOLDS and O'DON-
NELL (1985) and on data described in this report. Homozygous
viable, eye-specific alleles comprise class I. Class I1 includes non- complementing alleles and classes 111 and IV alleles complement each other to varying degrees. Class V represents embryonic- specific alleles that, in most cases, fail to complement other lethal Pu mutations. The new alleles described in this report are indicated with asterisks.
New Punch Mutants 61 1
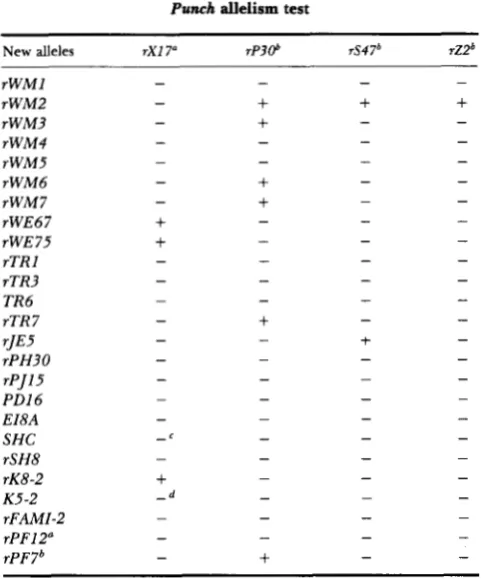
TABLE 1
Punch allelism test
New alleles rX17" rP306 rS47' rZ2'
rWM1 rWM2 rWM3 rWM4 rWM5 rWM6 rWM 7 rWE67 rWE75 rTR I rTR3 TR6 rTR 7 rJE5 rPH30 rPJl5 PD16 EI8A SHC
rSH8
rK8-2
rFAMI-2
rPF12" rPF7b K5-2
* Complementation for eye pigment phenotype only. All com- binations, except those noted in footnotes c and d , are fully viable. Complementation for viability only. All surviving truns-heter- ozygotes show some degree of eye pigmentation defects.
Heteroallelic class exhibits partial lethality. Heteroallelic class is inviable.
ing complementation groups. These have been sub- jected to a separate analysis, and the results are
described in a separate report
(J.
O'DONNELL and R. BOSWELL, unpublished observations). All mutants that failed to complement at least onePu
testor allele are listed in Table 1.Thirteen new alleles behave as standard noncom- plementing Pu mutations. They are lethal in combi- nation with P U ' ~ ' ~ , and Pur'', the represen- tatives of classes 11, I11 and IV, respectively. Progeny heterozygous for any of these alleles and the eye- specific allele, rXI7, are viable, but have mutant eye color. Two of the noncomplementing alleles (rWM4 and T R 6 ) have been included in all subsequent characterizations in order to verify that they affect P u function and GTP CH activity as predicted from previous studies. T h e remaining noncomplementing alleles are not considered further in this report.
Six alleles are capable of intragenic complementa- tion. Five of these (rWM6, rTR7, rWM3, rWM7 and rPF7b) complement the class I11 allele, rP30, while one ( r J E 5 ) complements the class IV allele, rS47. Inter se complementation tests of these new alleles against all previously identified complementing al- leles reveal that each new allele behaves in a predict-
able manner and that each falls within a previously identified complementation group. The complemen- tation data are presented in Table 2. As in previous complementation tests (MACKAY, REYNOLDS and O'DONNELL 1985), the viability frequencies are allele- specific and range as low as 1% of expected to essentially 100% of expected. Similarly, all survivors showed allele-specific effects on eye pigmentation and ranged from poorly pigmented to nearly wild- type eye color.
One allele, rWM2, appears to be a new member of the eye-specific class (class I). It was originally isolated as an eye color mutant in the deficiency chromosome screens and subsequent tests show that PurwMz hem- izygotes are fully viable (data not shown). The allele in combination with each lethal Pu allele similarly results in progeny that exhibit normal viability fre- quencies, but strong eye color phenotypes. PurwM21
eye-specific mutants, they have severe eye pheno- types. Presently, this mutant is homozygous lethal, presumably as a result of a second site lethal mutation on the second chromosome since the deficiency trans-
heterozygote is viable.
Five mutants gave atypical results in the P u allelism tests. These mutants, designated WE67, WE75, K8-2, K5-2 and SHC, behaved as Pu mutants in some of the tests, but each had genetic characteristics that had not been previously observed for any P u alleles. These characteristics define two new classes of Pu mutations.
WE67
and
KS-2: WE67 and K8-2 failed to com- plement the lethality of rP30, rS47 and rZ2, and by these criteria appear to possess strongly mutantPu
alleles. However, both appear to complement fully the eye color phenotype of rX2 7. In all previous cases, lethal allele combinations with rXI7 resulted in prog- eny with very strong eye color phenotypes. Both mutants were crossed inter se to all complementing mutants, and P uwas also crossed to 11 noncom- ~ ~ ~ ~ ~ plementing mutants. In each cross, at least 500-1000 progeny were screened, and in no case were trans- heterozygotes observed (data not shown).WE75: Another mutant, WE75, also produced truns-heterozygous progeny with wild-type eye color when crossed to PurX1', while failing to complement the lethality of several other
Pu
mutants. Unlike rWE67 and rK8-2, however, this allele in combination with several other Pu alleles permits some viability. trans-Heterozygous combinations with complement- ing alleles in general survive at about half of the expected frequency, and combinations with some normally noncomplementing alleles, survive to more limited extents, usually at 10% or less of expected values (Table 3). The most reasonable explanation of PurwE75 is that it is a hypomorphic allele that seems to affect embryonic function more severelyTABLE 2
Interallelic complementation of Punch mutants"
Class 111 Class IV New alleles
rP42 rP30 rP11 rS47 1219 119 rP43 rJE5 rWM3 rWM6 rWM7 rTR7 rPF7b
Class I11
rP42 rP30
Class IV
r P I I
rS4 7 r Z I 9 r19 rP43
New alleles
rJE5 rWM3 rWM6 rWM7 rTR 7 rPF7b
- -
+
+
+
+
+
-+
+
+
+
+
--
+
+
+
-+
+
+
+
+
+
+
+
A ''
+
" indicates viable heteroallelic progeny and a"-
" indicates failure to observe hereroallelic progeny in sample sizes of 500- 1000individuals. All crosses were performed in both directions. The data from both sets of crosses were combined since identical results were obtained in all cases. The results of crosses between previously tested mutants, those indicated as classes I1 and IV, are reproduced here for clarity. The crosses are described in detail in MACKAY, REYNOLDS and ODONNELL (1985).
TABLE 3
PurwE75 trans-heterozygote viability frequencies" and eye colorb
Percent frequency of
PU1WE75 p PU.WE7J d eye color Relative
Class I1 (noncomplementing)
P U Z 0 0
PUP' 0 0
P U S H C 0 0
3
-
-
p U r W M l 0 0
pu7TR6 0
puvTR5 0 6 + 3 - + 4
p u r M 1 7 0 0 + 4
-
-
-
Pur" 2 6 + 5
p U T V 1 4 2 2 + 4
P d V 9 2 0.3 + 4
P U T Z Z 9 15 + 4
pun22 22 24 + 4
pUrTR3 46 54 + 5
purTRI 76 61 + 4
Class 111 (complementing)
pU7P30 45 41 + 5
p u r P 4 2 58 68 + 5
p u V E y 51 72 + 5
Class IV (complementing)
pUrTR7 0 0 -
p u ~ W M 3 0 0 -
pu.z19 4 3 + 4
puTP43 45 70 + 5
PUT' 66 23 + 5
p u r P I I 59 60 + 5
p u , S 4 7 66 51 + 5
" For each cross, a minimum of 500-1000 progeny were scored.
Viability was calculated as percentage of the expected number of progeny in the tram-heterozygous genotype class.
*
Eye color was subjectively scored on a+
1-+
5 scale, with+
5 being wild type.than the eye function. Pending a more thorough biochemical analysis of this mutant, we have classified it with rWE67 and rK8-2 since their phenotypic characteristics are similar.
K5-2:
lK5-', which confers a dominant eye colorphenotype, exhibited a quite different behavior. This allele not only failed to complement the lethality of
the Puletha' testor alleles, but it was lethal in combi-
nation with the homozygous viable mutation r X l 7
(Tables 1 and 4). Since we had never before observed lethal allele combinations with rX17, we performed control matings of lK5-' to dp cn a p x sp, the parental strain of r X l 7 , and to cn P u + 1 (2)Cb-1 bw splSM5, a
P u + strain with a different genetic background. Since
K5-2 was generated on a chromosome marked with
shv b cn bw, we would ordinarily expect any second-
site lethal mutations to be complemented in these crosses. However, we obtained no K5-2 transheter- ozygous progeny in either experiment. This allele therefore behaves as a dominant lethal mutation under some circumstances.
New Punch Mutants 613
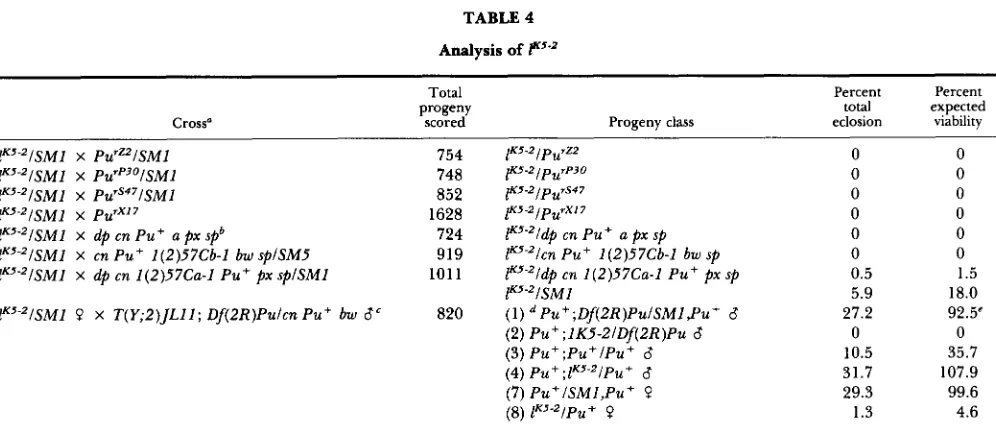
TABLE 4
Analysis of FsKss
Cross"
Total progeny
scored Progeny class
Percent total
Percent eclosion
expected viability lKs-'ISMl X PuTzzlSMI
~ 5 " I S M l X PU'~-)~ISMI lK5-zISMl X P U ' ~ ~ ~ / S M I F5"?2/SM1 X P U ' ~ ' ~ lK5-'ISM1 X dp cn Pu+ a p x spb
lK5-'lSMl X cn Pu+ 1(2)57Cb-I bw splSM5 lK5"ISM1 X dp cn 1(2)57Ca-l Pu + p x splSM1 FS-'ISMl P X T(Y;Z)JLlI; Df(2R)Pulcn Pu+ bw 6'
754 748 852 1628 724 919 101 1
820
1KS-21p~rZ2 1K5-ZlpurP30 lK5-Zlpu~S47 ~KS-21p~"Yl7
F5-'ldp cn Pu+ a p x sp lK5"lcn Pu+ 1(2)57Cb-1 bw sp lK5"ldp cn 1 (2)57Ca-l Pu+ p x sp F5"ISMI
(1) Pu+ ;Df(2R)PuISMI ,Pu+ 6 (2) Pu+;IK5-2IDf(ZR)Pu 6 (3) P u + ; P u + l P u + 6 (4) Pu+;lKJ"iPu+ 6 (7) Pu+ISMI,Pu+ P
(8) lK5"IPu+ P
0
0 0
0 0 0 0.5 5.9 27.2
0
10.5 31.7 29.3 1.3
0
0
0
0
0
0
1.5
18.0 92.5'
0 35.7 107.9 99.6 4.6
Crosses were performed reciprocally in all cases, except where directionality is indicated. Results of reciprocal crosses were similar, so
dp cn Pu+ a p x sp is the parental chromosome for all of the preceding Pu mutant chromosomes.
Translocated region is 56F-57F (LYTTLE 1984). Two clases of female progeny, those carrying the Df(2R)Pu chromosome, are not considered here because hypoploidy for the 56F-57F region of chromosome 2R is a completely lethal condition, regardless of the homologous chromosome.
the data have been pooled for this summary.
Numbers in parentheses identify the progeny classes illustrated in Figure 2.
The percent expected viability was based on the average number of adults obtained from the three progeny classes, ( l ) , (4) and (7),
which eclose at normal frequencies.
bers of that class. These flies developed very slowly and did not eclose until the cultures were 17-20 days old. Interestingly, K5-2 has a dominant lethal effect even when balanced against the SMZ chromosome derived from the other parental strain. These prog- eny were obtained at only 18% of the expected frequency for this class. Furthermore, they exhibit the same developmental delay as their K5-2ldp cn Z(2)Ca-Z Pu+ a p x sp siblings. In contrast, the dp cn
Z
(2)Ca-Z Pu+ a p x splSMI progeny began eclosing by day 10. We emphasize here that we do not observe a retardation of development nor excessive lethality in the K5-2fSMl stock. Viability frequencies of this stock will be reported below.K5-2 appears to have less drastic effects in combi- nation with the SMl chromosome described above as well as other rearrangement chromosomes tested (data not shown). We performed a final cross to try to determine whether these effects were, in fact, related to pairing phenomena. For this test we utilized a strain in which a section of the second chromosome, encompassing the P u locus, has been translocated to the Y chromosome, T(Y;2)JL-ZZ (LYTTLE 1984). The males that were crossed to K5-21SMZ females had a normal second chromosome, a normal segment of the second chromosome on the Y chromosome and a correspondingly deleted second chromosome. Dele- tion heterozygote females, which do not carry the translocation, are lethal, regardless of the composi- tion of the homolog. The cross and the expected progeny classes are illustrated in Figure 2 and the
viability frequencies of the progeny are presented in Table 4. The progeny classes most relevant to the question of pairing effects on viability are 2 and 8. Both classes possess one normal P u region and the K5-2 chromosome. In the male progeny, however, the normal P u region is on the Y chromosome and is not paired with the K5-2 chromosome, while in the female progeny the entire second chromosomes should be paired. We obtained no male survivors from this progeny class, but we did recover a small number of the comparable female class (4.6% of the expected progeny). Thus, the evidence rules out the transvection hypothesis as an explanation for the effects of K5-2. It is more likely that the strain in which K5-2 is maintained carries a suppressor of the dominant lethality, a possibility that we intend to investigate.
SHC: SHC resembles K5-2 in that it is a dominant eye color mutant that fails to complement the lethality of all lethal P u alleles tested, and it shows lethal effects in combination with the two eye-specific al- leles, rXZ7 and rZ (Table 5 ) . It differs from K5-2 in that the lethality conferred on the transheterozygous combinations is not complete. Furthermore, no evi- dence of lethal effects were obtained in crosses to the Put strain, dp cn a p x sp.
GTP
C H
assays: The new P u mutants that behaved614 E. R. Reynolds and J. M. O'Donnell
Pu +
- .
--
-"
Pu -
cn Pu +b w
Male progeny classes
Pu +
1
_ . "
Pu -
SMICycnPu+
-
Pu+/Pu+/Df Pu
PU+ cn Pu+bw
3
-.-
PU+/PU+/ PU+
" SMICycnPu+
PU+ cn Pu'bw
-.-
-"
Pu'/ Pu+
/
K5-2x - . -
"
K5-2
SMI Cy cn Pu+
c
Female progeny classes
Pu -
.
"" SMICycnPu+
Df Pu
/
Pu+Pu -
"
"
DfPu
/
K5-2cn Pu+bw
. -
-"
SMlCycnPu'
PU+
/
PU+-
cn Pu'bw' K 5 - 2
P u + / K5-2
FIGURE 2.-Chromosomal arrangement of progeny from the cross of T(Y;Z)JL-II 6 X lK5"ISMI P . Thin lines represent the X
and Y chromosomes; bars represent chromosome 2. The P u - chromosomes carry a deletion of 56F-57F. Individuals having only one dose of the 56F-57F are inviable under all conditions, and therefore progeny classes 5 and 6 are not expected.
severely diminished activities, usually in the range of 3-10% of normal, as previously observed (MACKAY, REYNOLDS and O'DONNELL 1985).
T h e three alleles that by genetic criteria appear normal with respect to their ability to function in adult eyes, rWE67, rK8-2 and rWE75, also have very high GTP CH activities. In fact, in each case, the wild-type heterozygotes have greater than wild-type activity in adult heads. Similarly, these alleles behave as wild-type alleles in combination with the rXI 7
allele. In rWE67 and rWE75 heterozygotes there appears to be a partial loss of larval enzyme activity, but rK8-2 larvae have' normal enzyme activities. It is clear that whatever the mutant effects of these three alleles on early Pu function, the effects do not extend to Pu function in the pigmentation of adult eyes.
Both SHC and K5-2 have clear-cut effects on GTP CH activity. Aside from the slight eye color phenotype of these mutants, the assay data are the only data that allow us to conclude that the K5-2 mutation is indeed a Pu mutation, since its dominant lethality prohibits any conclusions based on complementation analyses.
TABLE 5
Analysis of pH'
Percent Total expected progeny viability of Cy' Cross scored class
lsHCISMI P X Purz21SMI d 305 0
PuVz2ISMI P X lsHCISMI 6 292 0
lsHcISMI 0 X Pu'JE51SMI 8 364 0
PuflE5/SMI P X lsHCISMI 8 832 0
lsHCISMI P X P U ' ~ ~ ~ I S M I d 559 0
P U ' ~ ~ ~ I S M I 0 X lsHCISMI d 719 0
lsHCISMI 0 X P U ' ~ ' ~ ~ 1134 26
PUTXI 7 ISM1
P X lsHCISMI d 687 54
lsHCISMl 0 X Pu"6 583 28
Pu" 0 X lSHCISMI 6 373 94
lsHCISMl 0 X dp cn Pu+ a p x sp6 394 116 dp cn P u + a p x sp P X lsHCISMI d 160 81
Viability of new
Pu
mutants: All lethalPu
mutants examined so far die during embryogenesis o r at larval hatching (MACKAY, REYNOLDS and ODONNELL 1985; REYNOLDS and O'DONNELL, 1987). We deter- mined the lethal phase of representatives of the new group of mutants and found in each case that all homozygous Pu lethality could be assigned to the embryonic period. The results of these experiments are presented in Table7.
No death above background levels was observed at later stages, except the larval death of SMlISMI homozygotes. Note that the amount of death observed in the test of K5-2 precisely corresponds to the Mendelian proportion of K5-2homozygotes in a cross of K5-2iSMI males and fe- males. There is no evidence during embryogenesis, nor at any later stage for dominant lethal effects of
K5-2 in this genetic background. We also determined the time and extent of lethality in progeny derived from crosses of K5-2lSMI to both dp cn a p x sp and rXI 7. In each case, we expect that 50% of the progeny should be K5-2 heterozygotes. In each case, we ob- serve approximately 50% lethality during embryo- genesis or the first larval instar. No excessive lethality is observed at any later stages, and the only adult survivors are the SMI heterozygotes.
DISCUSSION
New Punch Mutants 615
TABLE 6
GTP CH activity in new mutants
Pu genotype
Relative enzyme
activity in heads" activity in prepupae" Relative enzyme
P u + ISM1
'
100 100Pu + lPu"y' 65 f 10
pu7xXI7 IPu"y' PurwM41SMl
PuTR61SMl 45 f 3 40 f 10 PuTTR31SMI
Pu'/~"ISML 84 f 5 49 f 10 pu?m/pu'X'7
PuTWM3ISMI
PurTR71SMI 93 f 27 10 f 10
P U ' ~ ~ ~ ~ I S M I
12 f 3
56 2 2 38 f 5
IPurXl7 5 2 1
pUrWM4
puTR61pu"Y17 3 f l
79 f 5 17 f 1
pu~TR3~purXl 7 7 2 2
20 f 5
63 f 3 34 f 5 pu~WM3~purX17 6 f 5
pUrTR7fpUrX17 3 f 1
55 f 2 56 f 10
144 f 10 53 f 6 125 f 10 43 f 5 PurK8"ISMl 156 f 8 90 f 7
P U ~ ~ - ~ I S M I 53 f 5 57 f 8 PusHciSMl 49 2 7 32 f 6 pUrWE67/pUrX17
pu~WE75/pu"Yl 7
purK8-2~pu"YI 7 68 f 10 puK5-2{pu"YI 7 -
puSHCfpurXl 7 -
a The enzyme activity of the Pu+lSMl strain was 69.3 f 5.6
nmol of formate releasedlhrlmg protein in crude extracts of adult heads and 5.0 f 0.4 nmol of formate releasedlhrlmg protein in crude extracts of prepupae. A minimum of three separate prepa- rations and determinations were performed for each strain.
* The
Pu+ chromosome used for these assays was dp cn a px sp,the parental chromosome for many of the mutant chromosomes in this test.
poorly viable.
periments (MACKAY and O'DONNELL 1983; MACKAY, REYNOLDS and O'DONNELL 1985).
Previous complementation experiments show that Pu has both vital and eye pigmentation functions that are partially independent (MACKAY, REYNOLDS and O'DONNELL 1985). It should therefore be possible to obtain mutations that affect only one of the two functions. We had previously identified a class of mutations that affect only the eye pigmentation and not the vital function of Pu. One of the new classes of Pu alleles obtained in the current experiments establishes that the opposite class can also be readily obtained. Each of the three alleles identified to date fails to complement the viability defect of all other Pu lethal alleles, but shows no loss of eye pigment function. Crosses of mutants of this class to mutants of the eye-specific class result in full complementation of both eye pigmentation and viability defects.
Although the alleles of the "embryonic-specific" class do not cause decreases in the GTP CH activity of adult heads, partial loss of GTP CH activity is observed in the prepupal stage of two of the three
c puK5-2fpurX17 IS . a lethal combination, and PusHclPurx'7 is
TABLE 7
Embryonic viability of new mutantsa
Total embryos No. Percent Mutant strain scored unhatched unhatched
dp cn P u + a px spISMI 600 66 1 1
shv b cn Pu+ bwISM1 520 47 9
520 185
36
PuTR6/SMl 32 165 516 PurTR'ISMl
500 134
27
PugE51SMI 500 128 26
145 29
PurTR7ISM1 990 300 30
P U ' ~ ~ ~ ~ I S M I 26 128 488 ISM1 478 134 28
520 158
30 PurK5"ISMI 410 108 26
PUSHCISMI 440 118 27
pUrWM4 ISMl
pUrWMJ ISM1 500
purWE75
~ ~ 7 K 8 - 2 ISMl
a The only embryonic lethality expected is that resulting from
mutations in Pu plus a small background lethality that is strain dependent. SMlISMI is lethal during the larval period.
mutants. We have observed similar effects on the prepupal activity in eye-specific mutants. We hypoth- esize that the prepupal activity consists of two com- ponents, one shared by embryos and the other by adult heads. Further examination of GTP CH in the prepupal stage will help to clarify this issue. We note a consistent increase in the adult head activity of all mutants in the embryo-specific class. At present, we have no explanation for this increase.
That these new mutations have unique and very specific defects is quite graphically illustrated by their distinctive lethal phenotypes (REYNOLDS and O'DON- NELL, 1987). Other
Pu
lethal mutations result in phenotypes that are consistent with the role of pterin cofactors in the catecholamine pathway (WRIGHT1987). In contrast, the new alleles result in normally pigmented embryos that have defects in segment cuticle patterns. Some of the complementing alleles also exhibit this phenotype, albeit in some cases, to a lesser extent than those alleles that appear to be strictly embryonic-specific. Strikingly, the expression of the segment pattern phenotype versus the "unpig- mented" phenotype by mutants homozygous for a given complementing allele is absolutely correlated with the complementation group into which the alleles fall. Thus, alleles of complementation group I11 appear to be leaky versions of the noncomple- menting alleles of group I1 and share their unpig- mented embryonic phenotype. Alleles of comple- mentation group IV have defects similar to those of the noncomplementing embryo-specific alleles of group V.
J.
though the mutants have dominant eye color pheno- types that are similar to those of other Pu mutants. One might still argue that these mutations, since they are dominant, could represent other genes that in- teract with
Pu
or directly affect GTP CH function. Although we have not yet genetically mapped the K5-2 allele, we have searched extensively for other genes that affect GTP CH. We have located such genes, the most clear-cut being prune and raspberryon the X chromosome (MACKAY and O’DONNELL 1983; see also EVANS and HOWELLS, 1978). The effects of mutations in these genes, however, are restricted to the enzyme in the adult head, while the effects of the dominant lethal mutations are clearly more far- reaching. Cytological observations have provided no additional information. Because animals bearing the outcrossed K5-2 chromosome are so poorly viable, we have not been able to examine this chromosome. T h e SHC chromosome appears to be normal
u.
O’DONNELL, unpublished observations).It is not yet clear whether the dominant lethality of these two alleles is due to a defect in one specific
Pu
function, or whether the alleles are simply exerting strong antimorphic effects on all vital functions of the locus. It is interesting to note, however, that experiments with the SHC alleles suggest a degree of specificity in the effects of this mutation. Crosses between the SHC mutant and certain of the “eye- specific” Pu mutants display a maternal effect that is restricted to the embryonic viability and is not ob- served with respect to the eye pigmentation of sur- viving adults. These adults all have brown rather than black bristles, a previously unobserved pheno- type(J.
ODONNELL, unpublished observations). T h e bristle phenotype is consistent with the expected role of biopterin as a cofactor in melanin production, but all previous cases of poor cuticular pigmentation in Pu mutants have been restricted to embryos that die before hatching (REYNOLDS and O’DONNELL 1987). It is unclear whether the maternal effect on viability is related to the previously reported maternal effects of the embryonic specific Pu mutations (REYNOLDS and O’DONNELL 1987). K5-2 adults do not exhibit the bristle phenotype or the maternal effect.Initial complementation crosses of the K5-2 mutant showed that even though the mutation can be easily maintained in a balanced stock, is fully viable as a heterozygote in that stock and develops at a normal rate, it was lethal in combination with all tested Pu mutant alleles, as well as wild-type chromosomes. T w o possibilities for the viability of the K5-2 balanced strain were either that the structure or composition of the multiply inverted S M I chromosome sup- pressed the dominant lethal phenotype, perhaps by the disruption of homolog pairing, or that a second site suppressing mutation had arisen in the stock. A
pairing-dependent dominant lethality would be sim- ilar to a transvection effect, a phenomenon that has been observed in several other genetically complex Drosophila loci and which is defined as pairing- dependent allelic complementation of mutant genes (LEWIS 1954; JACK and JUDD 1979; GELBART 1982). In the present case, the phenomenon would be a synapsis-dependent negative complementation of a wild-type allele. However, subsequent crosses per- mitted us to rule out the possibility that homolog pairing was a prerequisite for the dominant lethality. They also permitted us to further examine the con- ditions of the lethal interactions. In every case, re- gardless of the genetic background, when the K5-2 mutation is outcrossed it has strong dominant lethal effects. Even in those cases where some K5-2 progeny are obtained, the progeny develop very slowly and eclose approximately 5-7 days later than non-K5-2 siblings. Because poor viability and very slow devel- opment are also observed when the homolog is the
S M l chromosome from another strain, it seems prob-
able that a second site suppressor of the lethality has arisen in the strain. The crosses of K5-2 described in this report permit us to eliminate the X chromosome as a site for the suppressor since the lethal phenotype is not affected by the directionality of the cross. Similarly, it does not segregate with K5-2 itself, permitting the elimination of one of the second chromosomes as the site of the suppressor. Further experiments to separate the suppressor of dominant Pu lethality from the strain and map it are in progress. T h e successful isolation of such a suppressor should provide access to other genes that interact with Pu during embryogenesis.
We thank BOB BOSWELL for sharing mutant strains. Expert technical assistance for portions of this work was provided by
JOELLE LEVITT and CLARE CHUNG. This work was supported in part by National Institutes of Health grant GM26757 and in part by National Science Foundation grant 8608696.
LITERATURE CITED
EVANS, B. A., and A. J. HOWELLS, 1978 Control of drosopterin synthesis in Drosophila melanogaster: mutants showing an altered pattern of GTP cyclohydrolase activity during development. Biochem. Genet. 16: 13-26.
GELBART, W. M., 1982 Synapsis-dependent allelic cornplemen- tation at the decapentaplegic gene complex in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 79: 2636-2640.
JACK, J. W., and B. H. JUDD, 1979 Allelic pairing and gene regulation: a model for the zeste-white interaction in Drosophila mehnogaster. Proc. Natl. Acad. Sci. USA 76: 1368-1372. LEWIS, E. B., 1954 The theory and application of a new method
of detecting chromosomal rearrangements in Drosophila melan- ogaster. Am. Nat. 88: 73-89.
LEWIS, E. B., and F. BACHER, 1968 Method of feeding ethyl methane sulphonate (EMS) to Drosophila males. Drosophila Inform. Serv. 43: 193.
LINDSLEY, D. L., and E. H. GRELL, 1968 Genetic Variations of
New P u n c h Mutants 617
LYTTLE, T . W., 1984 ChrCmisomal control of fertility in Droso- phila melanogaster I. Rescue of T(Y;A)BB"'58 male sterility by chromosomal rearrangement. Genetics 106: 423-434.
MACKAY, W. J., and J. M. ODONNELL, 1983 A genetic analysis of the pteridine biosynthetic enzyme, guanosine triphosphate cyclohydrolase, in Drosophila melanogaster. Genetics 105: 35- 53.
MACKAY, W. J., E. R. REYNOLDS and J. M. ODONNELL,
1985 Tissue-specific and complex complementation patterns in the Punch locus of Drosophila melanagaster. Genetics 111: MIWA, S. WATANABE, Y. and 0. HAYAISHI, 1985 6R-~-elythro-
5,6,7,8-Tetrahydrobiopterin as a regulator of dopamine and serotonin biosynthesis in the rat brain. Arch. Biochem. Bio-
NAGATSU, T., M. LEVITT and S. UDENFRIED, 1964 Tyrosine hydroxylase: the initial step in norepinephrine biosynthesis. J. Biol. Chem. 239: 2910-2917.
PHILLIPS, J. S., and H. S. FORREST, 1980 Ommochromes and pteridines. pp. 542-623. In: The Genetics and Biology of Droso-
885-904.
phys. 239: 234-241.
phila, Vol. 2d, Edited by M. ASHBURNER and T. R. F. WRIGHT. Academic Press, London.
REYNOLDS, E. R., and J. M. ODONNELL, 1987 An analysis of the embryonic defects in Punch mutants of Drosophila melanagaster.
Dev. Biol. 123: 430-441.
WAHL, R. C., C. K. WARNER, V. FINNERTY and K. V. RAJAGOPALAN,
1982 Drosophila melanagaster ma-I mutants are defective in the sulfuration of desulfomolybdenum hydroxylases. J. Biol. Chem. 257: 3958-3962.
WEISBERG, E. P., and J. M. O'DONNELL, 1986 Purification and characterization of GTP cyclohydrolase I from Drosophila melanogaster. J. Biol. Chem. 261: 1453-1458.
WRIGHT, T. R. F., 1987 Genetic and molecular organization of the dense cluster of functionally related, vital genes in the dopa decarboxylase region of the Drosophila melanogaster gen- ome. pp. 154-172. In: Results andproblems in CellDijferentiation,
Vol. 14, Structure and Function of Eucatyotic Chromosomes, Edited by W. HENNIG. Springer-Verlag, Berlin.