Copyright 0 1993 by the Genetics Society of America
Cell Cycle Arrest
of
cdc
Mutants and Specificity
of the RAD9 Checkpoint
Ted A. Weinert*'+ and Leland H. Hartwell"
*Department of Genetics, University of Washington, Seattle, Washington 98195, and 'Department of Molecular and Cellular Biology, University of Arizona, Tucson, Arizona 85721
Manuscript received August 24, 1992
Accepted for publication January 15, 1993
ABSTRACT
In eucaryotes a cell cycle control called a checkpoint ensures that mitosis occurs only after
chromosomes are completely replicated and any damage is repaired. The function of this checkpoint
in budding yeast requires the RAD9 gene. Here we examine the role of the RAD9 gene in the arrest
of the 12 cell division cycle (cdc) mutants, temperature-sensitive lethal mutants that arrest in specific
phases of the cell cycle at a restrictive temperature. We found that in four cdc mutants the cdc rad9
cells failed to arrest after a shift to the restrictive temperature, rather they continued cell division and
died rapidly, whereas the cdc RAD cells arrested and remained viable. The cell cycle and genetic
phenotypes of the 12 cdc RAD mutants indicate the function of the RAD9 checkpoint is phase-specific
and signal-specific. First, the four cdc RAD mutants that required RAD9 each arrested in the late S/
G2 phase after a shift to the restrictive temperature when DNA replication was complete or nearly
complete, and second, each leaves D N A lesions when the CDC gene product is limiting for cell
division. Three of the four CDC genes are known to encode DNA replication enzymes. We found
that the RAD1 7 gene is also essential for the function of the RAD9 checkpoint because it is required
for phase-specific arrest of the same four cdc mutants. We also show that both X- or UV-irradiated
cells require the RAD9 and RAD17 genes for delay in the GP phase. Together, these results indicate
that the RAD9 checkpoint is apparently activated only by DNA lesions and arrests cell division only
in the late S/G2 phase.
"
T
HE cell division cycle in eucaryotes proceeds bya defined sequence of events where late events
often depend upon completion of early events [re-
viewed in HARTWELL a n d WEINERT (1 989)]. T h e de-
pendency of events appears critical for t h e high fidel-
ity of chromosome transmission; an error in the order
of events, if uncorrected, could lead to chromosome
mutations (chromosome loss, gain, rearrangements)
resulting in abnormal growth or cell death. Controls
that enforce the dependency of events are called
checkpoints and are defined by mutations that allow
a late event to occur without completion of an early
event. Checkpoints may order events in development
as well as in the eucaryotic cell cycle (CUTTING et al.
1990). Distinct checkpoints in cell division appear to
regulate the dependence of mitosis on DNA replica-
tion [see HARTWELL a n d WEINERT (1 989) a n d ENOCH
a n d NURSE (1 9 9 l ) ] and the dependence of anaphase
on spindle function (LI and MURRAY 1991 ; HOYT,
TOTIS a n d ROBERTS 199 1). We anticipate that check-
points may regulate the order of additional cell cycle
events.
This study examines the checkpoint that enforces
the dependence of mitosis on completion of DNA
replication. Genes identified by mutations that allow
mitosis without completion of DNA replication in-
clude the R A D 9 gene in budding yeast (WEINERT a n d
HARTWELL 1988), the R C C l / p i m I + gene in a hamster
Genetics 1 3 4 63-80 (May, 1993)
cell line a n d in fission yeast (NISHIMOTO, EILEEN and
BASILICO 1978; MATUSOMOTO a n d BEACH 1991), the
bimE gene in filamentous fungus (OSMANI et al. 1988), the cdc2+, cdc25+, weel+ a n d mikl+ genes in fission
yeast (ENOCH a n d NURSE 1990; LUNGREN et al. 1991;
ROWLEY, HUDSON a n d YOUNG 1992), and more re-
cently five rad+ genes also in fission yeast (ROWLEY,
SUBRAMANI and YOUNG 1992; AL-KHODAIRY and
CARR 1992). Study of the mutants and corresponding wild-type gene products should provide insight into
both the physiological role of the checkpoint controls
and the molecular events that enforce the dependency
of events. A proposed molecular mechanism for a
checkpoint control involving the cdc2+ protein kinase
is addressed in t h e DISCUSSION.
T h e role of t h e R A D 9 checkpoint in budding yeast
has been examined by comparing cell cycle pheno-
types of wild-type a n d rad9 null mutant cells. When
unirradiated, rad9 null mutants are viable and have
generation times and cell cycle kinetics indistinguish- able from wild-type cells but lose chromosomes spon- taneously a t a higher rate than wild-type cells (WEI- NERT a n d HARTWELL 1990). Thus the R A D 9 check- point is normally nonessential for cell viability though
it does contribute to genomic stability. When cells
contained DNA lesions due either to X-irradiation or
to defects in DNA ligase, wild-type cells delayed o r
64 T. A. Weinert and L. H. Hartwell
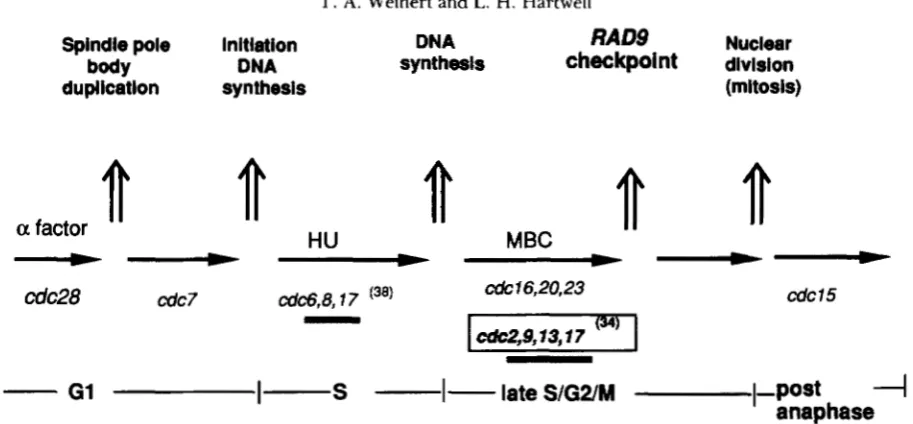
Spindle pole initiation DNA
RAD9
Nuclearduplicatlon synthesis (mitosis)
DNA synthests
checkpoint
divisiona
factor
HU
MBC
cdc28 cdc7 cd&,8,17 cdc16,20,23
cdc2.9.13.1 7 (
-
34)cdcl5
I-
G1l-
S
-1-
late
S/G2/M
1-post
4
anaphase
FIGURE 1 .-Dependent events in the yeast cell cycle. The relationship between cell cycle events (top), arrest of cdc mutants after shift to the restrictive temperature, and phases of the cell cycle. For example cdcljr mutants after shift to the restrictive temperature nearly complete or complete DNA synthesis and arrest in the late S/G2/M phase. Origins and features of the model are discussed in the text. Independent or interdependent relationships are not shown. cdc mutants that cause an increase in mitotic recombination when limiting for cell division are underlined (HARTWELL and SMITH 1985). cdc mutants within the open box require intact RAD9 and RAD17 genes for cell cycle arrest at the restrictive temperature (this paper; HARTWELL and WEINERT 1989). CDC genes whose enzymatic functions are know include: CDC28 protein kinase (LORINCZ and REED 1984); CDC8 thymidylate kinase (SCLAFANI and FANGMAN 1984); CDC9 D N A ligase (JOHNSTON and NASMYTH 1978); CDC17 DNA polymerase I
UOHNSON
et al. 1985; CARSON 1987); and CDC2 D N A polymerase I11 (SITNEY, BUDD and CAMPBELL 1989;BOULET et al. 1989).
cells continued cell division without delay and died (WEINERT and HARTWELL 1988, 1990; HARTWELL and WEINERT 1989). T h e RAD9 gene is not essential for DNA repair per se because a delay in G2 imposed by incubation with a microtubule poison suppressed the radiation sensitivity of rad9 mutants (WEINERT and HARTWELL 1988). T h e RAD9 checkpoint pro- vides a cell cycle delay to allow completion of DNA replication and repair before mitosis.
In this report we examine the function of the RAD9
checkpoint in cell division from the interactions of
rad9 with cdc mutants, temperature-sensitive lethal mutants that arrest in specific phases of cell division after shift to the restrictive temperature. We antici- pated that interactions between rad9 and cdc mutants might reveal the specificity of the RAD9 checkpoint as well as the basis for cell cycle arrest in cdc mutants. For many CDC gene products and their corresponding mutants the biochemicaI and ceIl cycle functions have been characterized (see Figure 1) although the basis for the cell cycle arrest is unknown. In principle, the arrest of cdc mutants may be due directly to the primary lesion in the CDC gene product or a second- ary consequence of a checkpoint control. For example
cdc28 mutants encode a temperature-sensitive protein kinase which at the restrictive temperature arrests cell division in G I (or in G2) (HARTWELL et al. 1973; PIGGOT, RAI and CARTER 1982; SURARA et al. 1991; REED and WITTENBERC 1990; reviewed in NURSE 1990). Presumably cell cycle progression requires the
activity of substrates of the CDC28 protein kinase that remain unphosphorylated and therefore inactive at the restrictive temperature; cdc28 mutants therefore arrest as a direct consequence of the CDC28 lesion. In contrast, cdc9 mutants encode a temperature-sensitive DNA ligase (JOHNSTON and NASMYTH 1978) which at restrictive temperature results in unligated Okazaki fragments and cell cycle arrest that requires the RAD9
gene (HARTWELL and WEINERT 1989); arrest in cdc9
mutants is a consequence of a checkpoint.
We find here that the cell cycle arrest of 4 cdc
mutants (of 12 tested) at the restrictive temperature requires the RAD9 checkpoint. From the phenotypes of the cdc mutants examined, together with studies of irradiated and hydroxyurea-treated cells, we conclude that function of the RAD9 checkpoint is signal- and phase-specific; it is activated only by DNA lesions that arrest cell division specifically in the late S/G2 phase. Further support for these conclusions comes from our studies of cdc mutants together with a second check- point gene, RAD1 7.
MATERIALS AND METHODS
Yeast strains: T h e yeast strains used in this study (Table
1) are isogenic with strain A364a except where noted.
Strains were derived using standard genetic techniques
(SHERMAN, FINK and HICKS 1986). T h e original strain con-
taining the radl7-1 allele, X-55C (MATa RAD17-1 ade2-I
obtained from the Berkeley Stock Center) was noncongenic with A364a, the strain background used in these studies.
cdc Mutants and the RAD9 Checkpoint
TABLE 1
Yeast strains
65
Strainn Genotype
7859-10-2b MATa 7859-7-4b h2c2al 7867-2-1 h6clal h7c4al 7843-3-1 h8clal 7844-1-3 598-3c 7851-3-2c 7845-7-4
h l 6 c l a l
h15c2al
h l 7 c l a l 7848-9-4 h20clal h23clal 7850-2-4 h28clal 7868-13-3 7845-7-1 7848-2-1 7846-8-2 7849-1-3 7842-6-3 208-7-2 209-8-1 209-8-2 156-2-4 156-2-2 158-1-3 158-2-3 180-4-4 180-2-3 21 1-1-2 2 1 1-2-3 2 10-3-2 2 10-2-4 179-9-3 179-9- 1 155-3-2 155-7-3 228-1-3 228-2-3 208-4-1 208-2-3 159-3-2 159-7-1 7870-1-1 7870-1-4 7870 MATa MATa MA Ta MATa MATa MATa MA Ta MA Ta MATa MATa MA T a MA Ta MA Ta MA T a MA Ta MA Ta MATa MA Ta MA Ta MATa MA T a MA Ta MA Ta MA Ta MATa MA T a MATa MATa MA Ta MATa MA T a MA Ta MATO MA T a MA Ta MATa MA Ta MA Ta MATa MA Ta MA T a MATa MA T a MATa MA Ta MATa MA Ta MATa MATa MATa MATa/MATa
his7 leu2 trpl ura3
his7 leu2 trpl ura3 rad9A::LEU2 cdc2-2 his7 u r a l
cdc2-2 rad9A::LEU2 his7 u r a l u r a 3 cyh2 canl cdc6-1 his7 u r a l
cdc6-1 rad9A::LEU2 cdc7-4 his7 u r a l
cdc7-4 rad9A::LEU2 his7 u r a l u r a 3 cdc8-1 his7 u r a l
cdc8-1 rad9A::LEU2 his3 t r p l u r a l cyh2
cdc9-8 ade2 ade3 trpl leul-1 ura? SCE::URA3 canl cyh2 sap3 cdc9-8 rad9A::TRPl ade2 ade3 his3 leul leu2
cdcl3-1 his3 trpl ura3 canl
cdcl3-1 rad9A::LEU2 his? t r p l u r a l u r a 3 c a n l cyh2 cecl6-1 his7 u r a l
cdcl6-1 rad9A::LEU2 his3 trpl ura3 cyh2 canl cdc15-2 his7 u r a l
cdc15-2 rad9A::LEU2 his3 trpl canl cdcl7-1 his7 u r a l
cdc17-I rad9A::LEU2 his3 t r p l u r a l cyh2 cdc20-1 his7 u r a l
cdc20-1 rad9A::LEU2 his3 trpl ural canl cdc23-1 his7 u r a l
cdc23-1 rad9A::LEUP his7 cyh2 cdc28-1 his7 u r a l
cdc28-I rad9A::LEU2 his3 trpl ural ura3 cyh2 rad17-1 ura? his7
cdc2-2 ura3 his7 canl
cdc2-2 r a d l 7 - 1 u r a 3 his7 canl Ade- cdc6-1 canl
cdc6-1 r a d l 7 - 1 u r a l t r p l cdc7-4 u r a l u r a 3 his7 canl cdc7-4 r a d l 7 - 1 u r a l his7 cdc8-1 t r p l u r a l u r a 3 his7 cdc8-1 r a d l 7 - 1 t r p l u r a 3 his7 cdc9-8 ade2 leul ura3 trpl canl
cdc9-8 rad17-1 ade2 ade3 trpl ura3 his7 canl cdcl3-1 trpl ura3 his7 canl
cdcl3-1 radl7-1 trpl ura? canl cdc15-2 his7 canl
cdc15-2 r a d l 7 - 1 t r p l u r a l u r a 3 his7 canl cdcl6-1 ura3 trpl his7 his3
cdcl6-1 radl7-1 ural ura3 cdcl7-1 ura3 his3 leu2 trpl cdcl7-1 radl7-1 ura3 his3 leu2 cdc20-1 his7
cdc20-1 r a d l 7 - 1 u r a l u r a 3 his7 canl cdc28-1 his7 trpl canl
cdc28-I r a d l 7 - 1 his7 u r a l u r a 3 t r p l c a n l u r a l his7
r a d l 7 - 1 u r a l his7 ura3
r a d l 7 - l / + u r a 3 / + m a l / + his7/+ t r p l / + c a d / + c d c l 3 / +
Strains with an “h” designation (e.g., h2c2al) from the collection from the HARTWELL laboratory. Strains were generated in this study
WEINERT and HARTWELL (1 990). HARTWELL and WEINERT (1989). with the following exceptions, as noted below.
66 T. A. Weinert and L. H. Hartwell
we consider this a null mutation (WEINERT and HARTWELL
Media: Cells were grown either in complete or rich me-
dium as noted (SHERMAN, FINK and HICKS 1986; HARTWELL
1967). For studies using radiation, X rays were delivered by
a Machlett OEG60 tube and U V by a Stratalinker 1800 set at 40 J/m'. Hydroxyurea was obtained from Sigma, pre-
pared immediately before use as a 2 M solution and used at
a final concentration of 0.2 M at 30". Methylbenzimadazole- 2-yl-carbamate (MBC) was obtained as a gift from Du Pont, prepared as a 10 mg/ml stock solution in 100% dimethyl- sulfoxide (DMSO) and used at a final concentration of 100
Tests of cell cycle arrest in cdc mutants: Cells were
grown to logarithmic phase (at 2-5 X lo6 cells/ml) in com-
plete liquid medium at the permissive temperature (23")
and shifted to the restrictive temperature (34", 36" or 38"
as noted in Tables 2 and 5 ) for 4 hr. (We obtained similar
results when cells were grown in rich or complete medium
for the strains we tested; data not shown.) Four cell cycle
parameters were analyzed: cell and nuclear morphology,
first cycle arrest, cell viability and maximum permissive temperature for growth (each is discussed below). The fate of at least 100 cells was scored for each of the phenotypes.
We also determined the DNA content for a subset of strains
as indicated in Table 2.
Nuclear and cell morphology: Cells were fixed with 70% ethanol, stained with the DNA-binding dyes 4,6-diamino-2-
phenylindole (DAPI) (PRINGLE et al. 1989) or with propi-
dium iodide after RNase treatment for 2 hr (BAUM et al.
1988; WEINERT and HARTWELL 1990), and viewed using fluorescence and light microscopy.
First cycle arrest: A "microcolony assay" was used to
determine whether cdc mutants arrested in the first cell
cycle after a shift to the restrictive temperature. Cells grown in liquid medium at the permissive temperature were soni-
cated to disrupt clumps and plated on prewarmed agar plates
in complete medium. After 4 hr at the restrictive tempera- ture random fields of microcolonies that were well dispersed were viewed by light microscopy to score cell cycle arrest.
The number of buds in each of at least 100 microcolonies
(each produced by a single cell) were recorded. Eleven of
the cdc RAD mutant strains tested here arrest as large-
budded cells at the restrictive temperature. Therefore in
the microcolony assay any cell gives rise to either 2 buds (1
large-budded cell) or 4 buds (2 adjacent large-budded cells)
depending on whether the cell was before or after (respec-
tively) the execution point for the cdc mutant at the time of
temperature shift. T h e cdc28 mutants studied here arrest as
unbudded (GI) cells and therefore generate microcolonies that contain either 1 or 2 unbudded cells. We calculated the percentage of cells that show a first cycle arrest from the
number of microcolonies that contained 2 or 4 buds (ar-
rested) divided by the total number of microcolonies. Cells
that gave rise to microcolonies that contained three or 2 5
buds we concluded did not undergo an efficient first cycle arrest. Typically an efficient first cell arrest is indicated if
>80% microcolonies contain either 2 or 4 buds; however,
comparison of mutants that arrest less efficiently also proves
informative.
T h e microcolony assay may in some instances over-
estimate the first cycle arrest; for example cells that are
before the execution point at the time of temperature shift and generate four buds (instead of 2 buds) during the 4-hr incubation would be scored as "arrested." (Time-lapse pho- tomicroscopy, though laborious, avoids this particular prob-
lem. We have used both the microcolony and time-lapse
photomicroscopy assays to test for first cycle arrest in cdc9
1990).
pg/ml.
and cdc9 rad9 strains and found that results from the two
assays are quantitatively similar; 79% and 80% of cdc9 cells
and 36% and 43% of cdc9 rad9 strains, respectively showed
a first cycle arrest [HARTWELL and WEINERT (1989) and
Table 2 in this study.]) T h e microcolony assay does not
reveal the nuclear morphology of the cells. Therefore, we find results from both the microcolony assay, the cell and nuclear morphology assay, and the cell viability assays (see
below) are required to assess whether mutants arrest cell
division synchronously at the restrictive temperature. Cell viability: Cell viability was determined for cells in- cubated at the permissive temperature of 23" and after 4
hr at the restrictive temperature in liquid culture. Cells were
plated on solid agar medium and colony formation deter-
mined by light microscopy usually after 1 day of growth at
the permissive temperature; viable cells give rise to "macro-
colonies" that contain typically > 50 cell bodies (buds) and
inviable cells to microcolonies of < l o cell bodies. In some
instances viability was determined from visible colony for- mation after 3 days of incubation. At least three determi-
nations were made for each strain, and the mean values and
standard deviations are shown (Tables 2, 3 and 5 ) . We also
determined the cell viability for many cdc RAD and cdc rad9
strains after incubation for up to 8 hr at the restrictive
temperature [see HARTWELL and WEINERT (1 989); data not
shown]. We found that the differences in viability after 4 hr
at the restrictive temperature was representative of the
differences in viabilities from later times and permits cor- relation with cell cycle arrest phenotypes which were also
determined after 4 hr. We do not consider viabilities that
differ by less than twofold between the cdc RAD and cdc rad
strains as significantly different especially if other pheno- types are similar.
DNA content: Flow cytometry was performed on cells
that were fixed with 70% ethanol, digested with RNase for
2 hr at 37", stained with 5 0 pg/ml propidium iodide (PI),
and diluted into 5 pg/ml PI just prior to analysis by flow
cytometry (BAUM et al. 1988). At least 5000 cells were
counted in each analysis. T h e position of the 1C and 2C
peaks were confirmed where tested with a factor- or MBC-
arrested cultures (arrested in G I and GP, respectively; data
not shown).
Maximum permissive temperature for growth Cells
were streaked for single colonies onto agar plates containing
complete medium and incubated at 23 O , 2 5 O , 28 O , 30°, 32"
or 36" for 2-4 days. T h e maximum permissive temperature
is the highest temperature tested at which visible colony
formation was comparable to that seen at the permissive
temperature of 23". At temperatures higher than the max-
imum permissive temperature typically no or a few visible
colonies are formed. cdc6 rad9 strains consistently formed a
few colonies (papillation) at one temperature interval higher
than the maximum permissive temperature.
The role of RAD17 in cell cycle control: tests for cell cycle arrest after X-irradiation: T h e cell cycle arrest in
RAD and radl 7 cells after X-irradiation was determined by
time-lapse photomicroscopy (WEINERT and HARTWELL
1988). Logarithmically growing cells were plated on solid
agar slabs (in rich medium), X-irradiated ( 2 krad) and pho-
tographed at the time of plating and after incubation for 8-
10 hr at 23 O . Pairs of photographic negatives were projected
and the fate of individual cells determined.
Genetic linkage of cell-cycle arrest defect and radiation sensitivity in r a d l 7 mutants: We tested for cosegregation of cell cycle arrest-defect and radiation sensitivity in strains
produced from a r a d l 7 / + heterozygote (strain 7870). Log-
arithmically growing cells were plated on solid agar media
cdc Mutants and the RAD9 Checkpoint
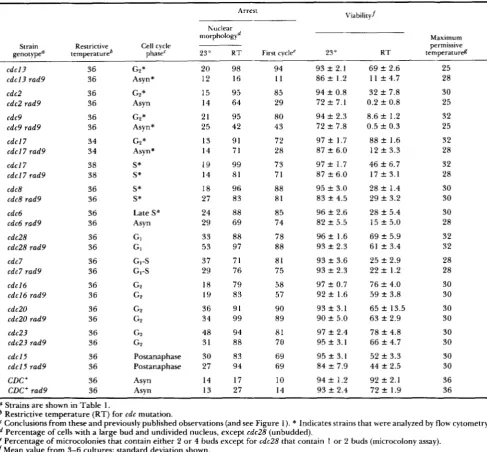
TABLE 2
Requirement for RAD9 in cell cycle arrest of cdc mutants
67 Arrest Viabilityf morphologyd Nuclear Maximum Strain genotypea Restrictive temperatureb
Cell cycle permissive
phase' 23" R T First cyclee 23" t e m p e r a t u r d R T c d c l 3
cdcl3 rad9
cdc2 cdc2 rad9
cdc9 cdc9 rad9
cdcl7 cdcl7 rad9
cdcl7 cdcl7 rad9
cdc8 cdc8 rad9
cdc6 cdc6 rad9
cdc28 cdc28 rad9
cdc7 cdc7 rad9
cdcl6 cdcl6 rad9
cdc20 cdc20 rad9
cdc23 cdc23 rad9
cdcl5 cdcl5 rad9
CDC+ CDC+ rad9 36 36 36 36 36 36 34 34 38 38 36 36 36 36 36 36 36 36 36 36 36 36 36 36 36 36 36 36 GP* Asyn* G2* As y n
G2* Asyn*
GP * Asyn *
S* S*
S* S*
Late S* Asyn GI G1 GI-S G,-S GP GP GP GP G P G2 Postanaphase Postanaphase Asyn Asyn
20 98 12 16
15 95
14 64
21 95 25 42
13 91 14 71
19 99 14 81
18 96
27 83
24 88 29 69
33 88
53 97
37 71 29 76
18 79 19 83
36 91
34 99
48 94
31 88
30 83 27 94
14 17 13 27
94 1 1 85 29 80 43 72 28 73 71 88 81 85 74 78 88 81 75 58 57 90 89 81 70 69 69 10 14
9 3 f 2.1 86 f 1.2
94 f 0.8 72 f 7.1
94 f 2.3 72 ? 7.8
97 f 1.7 87 f 6.0
97 f 1.7 87 f 6.0
95 f 3.0 83 f 4.5
96 f 2.6 82 f 5.5
96 f 1.6 93 f 2.3 93 f 3.6 9 3 f 2.3
97 f 0.7 92 f 1.6
93 f 3.1 90 f 5.0
97 f 2.4 95 f 3.1
95 f 3. I 84 f 7.9
94 f 1.2 93 f 2.4
69 f 2.6 1 1 f 4.7
32 f 7.8 0.2 f 0.8
8.6 f 1.2 0.5 f 0.3
88 f 1.6 12 f 3.3
46 f 6.7 17 f 3.1
28 f 1.4 29 f 3.2
28 f 5.4 15 f 5.0
69 f 5.9 61
*
3.4 25 f 2.9 22 f 1.276 f 4.0 59 f 3.8
65 f 13.5 63 f 2.9
78 f 4.8 66 f 4.7
52 f 3.3 44 f 2.5
92 f 2.1 72 ? 1.9
25 28 30 25 32 25 32 28 32 28 30 30 30 28 32 32 28 28 30 30 30 30 30 30 30 30 36 36
a Strains are shown in Table 1.
Restrictive temperature (RT) for cdc mutation.
Conclusions from these and previously published observations (and see Figure 1). * Indicates strains that were analyzed by flow cytometry. Percentage of cells with a large bud and undivided nucleus, except cdc28 (unbudded).
Percentage of microcolonies that contain either 2 or 4 buds except for cdc28 that contain 1 or 2 buds (microcolony assay).
f Mean value from 3-6 cultures; standard deviation shown.
g Maximum permissive temperature of colony formation.
cycle arrest was determined from the percentage of micro-
colonies that contained either 2 (1 large-budded cell) or 4
buds (2 large-budded cells) 10 hr after plating and X-
irradiation. Cell lethality was determined from macrocolony
formation 24 hr. after X-irradiation at the same dose. Ra-
diation sensitivity was determined by exposing freshly rep-
licated patches of cells to 8 krad of X rays twice at 8-hr
intervals and scoring growth of survivors after incubation
for 2-3 days at 23". The cell cycle arrest defect was quan-
tified using a convenient metric calculated from the per- centage of X-irradiated cells that arrest divided by cell
inviability at the same dose (WEINERT and HARTWELL 1988,
1990). Typically the ratio of cell cycle arrest to cell inviability
is >0.8 for arrest-proficient strains (e.g., RAD) and (0.3 for
arrest-deficient strains (e.g., rad1 7).
Delay of nuclear division (mitosis) by irradiation: T o
quantify the delay in nuclear division by DNA damage, cells growing logarithmically in rich liquid medium were first
synchronized in the G P phase by plating on agar plates
containing the microtubule inhibitor MBC and incubated
for 3 hr at 23". DNA damage was induced in the MBC-
blocked G P cells by irradiation (X rays, 4 krad; or UV, 40
J/m') and cells were immediately resuspended in medium, washed twice to remove the MBC and resuspended in fresh medium. The kinetics of progression through mitosis was
determined by anal sis of nuclear morphology. Aliquots of
cells were fixed wit
K
70% ethanol at specific times, stainedwith DAPI and nuclear morphology determined by fluores- cence microscopy.
RESULTS
68 T. A. Weinert and L. H. Hartwell cycle arrest in
12
cdc mutants which have been char-acterized physiologically and genetically, including some for which the biochemical functions are known (see legend to Figure 1). We reasoned that interaction between the R A D 9 checkpoint and any of these cdc mutants would provide information on the basis for cell cycle arrest in the cdc mutants and on the phase and signal specificity of the R A D 9 checkpoint. Figure
1 shows the dependent events of the cell cycle defined by these cdc mutants (PRINGLE and HARTWELL 1981)
as modified by the results presented here (see DISCUS- SION). The phase of cell division where cdc mutants arrest at their restrictive temperatures is in some cases ambiguous. We can not at present distinguish by cytological or biochemical criteria late S phase (as defined below), the G2 phase, and metaphase (see
NURSE 1985). These distinctions are particular rele- vant because the R A D 9 checkpoint acts in this part of the cell cycle, and because better resolution would allow comparisons to checkpoint controls in other organisms where the three phases are better distin- guished. Formally we must describe the function of the R A D 9 checkpoint as acting in the late S/G2/M phase (Figure l), but we will refer to its function in the text as in “late S/G2 phase” reflecting our bias that it acts near the end or immediately after D N A repli- cation. T h e arrest of cdclb, cdc20, cdc23 and MBC- treated cells is referred as in “G*/M” because arrested cells in each case appear to contain fully replicated, intact sister chromosomes (KOSHLAND and HARTWELL
1987). The order of function of cdc mutants that are required in late S/G2 phase and those required in G2/
M phase has not been formally established beyond their order with respect to the block imposed by the microtubule inhibitor, benomyl (WOOD and HART- WELL 1982).
We used two sets of criteria to test the cell cycle arrest of cdc R A D and cdc r a d 9 mutants after shift to a restrictive temperature. A direct measure of cell cycle arrest comes from determination of the nuclear and cell morphology, and from whether the cells arrested in the first cell cycle (scored by observing microcolonies on solid medium) 4 hr after shift to the restrictive temperature. A second criterion was the effect of a r a d 9 mutation on cell viability when cells were limited for the cdc function. CDC function was limited either by incubation for 4 hr at the fully restrictive temperature (36”) or by continuous incu- bation at an intermediate temperature. For some CDC mutants, we predicted that limiting the CDC function generates errors that can be repaired provided cell division is delayed by the R A D 9 checkpoint. For these CDC mutants we expected the cdc rad9 mutant would have a lower viability compared to the cdc R A D mutant because cell division with uncorrected errors would be lethal. We measured synthetic lethality at both the
fully restrictive temperature where cdc R A D mutants arrest because they have insufficient CDC function to complete cell division as well as at an intermediate temperature where cdc R A D mutants delay because the residual CDC function may be limiting but suffi- cient for cell division (see for example HARTWELL and SMITH 1985; SCLAFANI and FANGMAN 1986). For ex- ample, cdc9 R A D mutants require the R A D 9 gene both for arrest and viability at the fully restrictive temper- ature (see Table 2; HARTWELL and WEINERT 1989)
as well as for viability, and presumably for delay of cell division, at intermediate temperatures (CDC9 ap- parently is limiting for cell division at intermediate temperatures because the percent of large-budded cells with an undivided nucleus increases with increas- ing temperature of incubation; data not shown). In sum, the two criteria of morphological cell cycle arrest and cell viability provide a comprehensive test for the role of the R A D 9 checkpoint in arrest of cells limited for a CDC gene product.
Arrest in the late S/G2 phase of four cdc mutants requires the RAD9 gene, We found that the cell cycle arrest of cdc2, cdc9, cdcl3 and c d c l 7 after shift to the restrictive temperature required an intact R A D 9 gene. Results of tests of morphological arrest and viability of cdcl? and c d c l 7 mutants are presented in detail below.
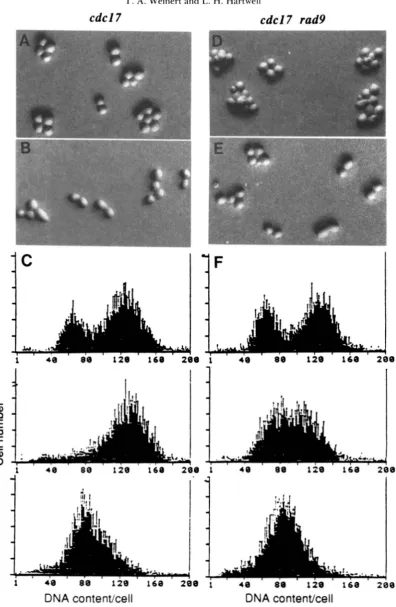
c d c l 3 R A D mutants shifted to the restrictive tem- perature for 4 hr arrested in the first cell cycle (Figure 2A) as large-budded cells with an undivided nucleus (Figure 2B) with approximately a 2C D N A content (Figure 2C, bottom). After 8 hr at the restrictive temperature most c d c l 3 R A D mutant cells remained arrested while cell growth continued (Figure 2A, in- set). In contrast, c d c l 3 r a d 9 mutants failed to arrest after shift to the restrictive temperature. The double mutant cells continued to divide for up to about 8 hr at nearly the rate of CDC cells incubated at 36”
(Figure 2D and inset; data not shown). The cell and nuclear morphology (Figure 2E) and D N A content (Figure 2F, bottom) of c d c l 3 r a d 9 cells after 4 hr at
36” resembled that of the asynchronous, exponen- tially dividing c d c l 3 r a d 9 culture at 2 3 ” . The cell viability of c d c l 3 r a d 9 mutants at the fully restrictive temperature was also significantly lower than that of the c d c l 3 R A D strain (Table 2), and the maximum permissive temperature was also different in the two strains, though in an unexpected way (see below). Comparing the data presented in Table
2,
we con- clude that nearly all c d c l 3 cells at the restrictive tem- perature require the R A D 9 gene for arrest and viabil- ity.cdc Mutants and the RAD9 Checkpoint
cdcl3
cdcl3 rad9
.
..,".... .. ..% . . "%"-."""/"69
DNA contentkell
DNA contentkell
FIGURE 2.-Cell cycle arrest of cdcl? and cdcl? rad9 strains. (A-C) cdcl?; (D-F) cdcl3 rad9. (A and D) Microcolony test of first cycle
arrest. Cells were grown in liquid medium at 23". plated on agar slabs and incubated at 36" for 4 hr. Insets: cells incubated at 36" for 8 hr. Photographs in A. D, and insets are same magnification. (B and E) Cell and nuclear morphology. Magnification is different from cells shown in A and D. (C and F) Flow cytometric analysis of D N A content in cells grown at 23" (top) or at 36" for 4 hr (bottom).
70 T. A. Weinert and L. H . Hartwell
cdcl7 cdcl7
rad9
rl
1
I
lF
1 40 88 1 2 0 160 208 1 40 80 120 168 2 0 0
DNA
contentkell
DNA
content/cell
FIGURE 3.-Cell cycle arrest of cdcl7 and cdcl7 rad9 strains. (A-C) cdcll; (D-F) cdcl7 rad9. (A and D) Microcolony test of first cycle
arrest at 34". (B and E) Microcolony test of first cycle arrest at 38". (C and F) Flow cytometric analysis of DNA content per cell in cells incubated at 23" (top) 34" for 4 hr (middle) or 38" for 4 hr (bottom).
71
i
t
I
i D
DNA contentkell
DNA contentlcell
FIGURE 4.-Cell cycle arrest of cdcd and cdcd rad9 strains. (A and B) cdcd; (C and D) cdcd rad9. (A and C) Microcolony test to determine
first cycle arrest at 36"; (B and D) flow cytometric analysis of DNA content per cell in cells incubated at 23" (top) or 36" for 4 hr (bottom).
4 hr cdcl7 RAD mutants did arrest in the first cell cycle in late S/G2 phase (Figure 3, A and C, middle), whereas cdcl7 rad9 did not arrest in the first cell cycle (compare Figure 3, A and D) and did not arrest with a late S/G2 phase content of DNA (Figure 3F, middle). In addition, the cell viability was decreased more dramatically in the cdcl7 rad9 strain compared to
cdcl7 RAD strain after incubation at 34" than after incubation at 38" (Table 2). We conclude that the cell cycle arrest and viability of cdcl7 mutants in late S/ GP phase requires the RAD9 gene but arrest of cdcl7
mutants in early S phase does not. (We speculate that the 2-3-fold lower viability of cdcl7 rad9 strains com- pared to cdcl7 RAD at 38" may reveal a requirement for some delay at the RAD9 checkpoint to repair DNA lesions formed at 38" after temperature shift down.)
We also found that two additional cdc mutants, cdc2
and cdc9, of the 12 cdc mutants tested required the
RAD9 gene for cell cycle arrest; cdc2 rad9 and cdc9
rad9 cells failed to cell cycle arrest and lost cell viability rapidly after 4 hr at the restrictive temperature com- pared to cdc RAD strains [Table 2 and HARTWELL and WEINERT (1989)l. We measured DNA content at the restrictive temperature and found that cdc2 RAD and
cdc9 RAD, but not the cdc rad9 mutants, arrest with an apparent 2C DNA content after 4 hr (data not shown). We conclude that arrest of cdc2 and cdc9
mutants in the late S/G2 phase requires the RAD9
gene. (From the data in Table 2 we can conclude that at least half of the cdc RAD cells for cdc2, cdc9 as well as for cdcl7 require the RAD9 gene for cell cycle arrest, though these quantitative estimates of cell cycle arrest may be underestimates; see MATERIALS A N D METHODS).
We also observed synthetic lethality in cdc2 rad9, cdc9 rad9, and cdcl7 rad9 mutants as measured by the maximum permissive temperature; the cdc rad9
72 T. A. Weinert and L. H . Hartwell
I
I
FIGURE 5.-Cell cycle arrest in X-irradiated RAD and rad 17 cells. Time lapse photomicroscopy of X-irradiated wild type (A and B) and radl 7 (C and D) strains. (A and C) Cells plated and X-irradiated
(2 krad) at t = 0 hr; (B and D) progeny of cells, t = 10 hr. Arrows indicate unbudded G I cells and their progeny, and asterisks indicate large-budded postanaphase cells and their progeny.
TABLE 3
Cell cycle response to hydroxyurea of
RAD+, rad9 or rad17 cells
Percent arrest Strain
genotype' H u b morphologvC First cvcled Viabilitv' Nuclear
RAD+
-
16 14 94 f 3.0+
85 87 73 f 13.3rad 9
-
12 8 95 f 1.6+
83 83 76 2 10.1rad I7
-
13 7 86 f 4.2+
95 92 83*
1.8'
Strains are RAD+ (7859-10-2); rad9 (7859-7-4); rad17 (208-7- 2).H U , hydroxyurea at final concentration at 0.2 M.
Percentage of cells with large bud and undivided nucleus. Percentage of microcolonies that contain either 2 or 4 buds. Mean value from triplicate samples; standard deviation shown.
ture for colony formation than did the cdc RAD mu- tants (Table 2). This result implies that cells with suboptimal levels of these CDC gene functions gener- ate viable progeny only when the RAD9 checkpoint prevents cell division until errors are corrected.
T h e maximum permissive temperature for c d c l 3 rad9 is exceptional because the cdcl3 rad9 double mutant formed colonies at a higher temperature (28") than did cdcl3 RAD mutant (25"). (The colony for-
TABLE 4
&segregation of radiation-sensitivity and defect in cell cycle arrest
Strain lethality arrest Arrest/lethality RAD
Percent Percent
7870-1-1 42 46 1.09
+
1-2 81 4 0.05
-
1-3 66 63 0.95
+
1-4 66 1 0.02
-
2- 1 79 6 0.08
-
2-2 63 54 0.86
+
2-3 72 1 0.01
-
2-4 40 40 1 .oo
+
3-1 73 2 0.03
-
3-2 64 49 0.77
+
3-3 59 10 0.17
-
3-4 42 40 0.95
+
RAD+ 48 42 0.92
+
radl 7 71 6.9 0.10
-
Meiotic products from a diploid strain 7870 heterozygous for
radl 7 were analyzed for cell cycle defect and radiation sensitivity.
Cell lethality, cell cycle arrest, and radiation sensitivity were deter- mined as described in MATERIALS AND METHODS. Results from 3 of the 12 tetrads analyzed are shown, and the averages of all 24 RAD+
and 24 rad I7 spores tested are given.
mation of cdcl3 rad9 cells is not due to suppressors of
cdcl3-nor of rad9; data not shown.) We think the reason cdcl3 rad9 cells form a colony at intermediate temperature is an exaggeration of its unusual pheno- type at high temperature (36") where cdcl3 rad9 cells sustain several cell divisions before cessation of divi- sion (see Figure 2); at the intermediate temperature
cdcl3 rad9 cells sustain more divisions (about 20), enough to form a colony. We posit that lethal DNA damage that leads to cessation of cell divisions requires more cell divisions to accumulate at intermediate tem- perature us. high temperature. This hypothesis pre- dicts that cdcl3 rad9 cells generated at the interme- diate temperature should contain a high fraction of inviable cells. This prediction was confirmed; the vi- ability of cdcl3 rad9 cells from colonies formed at the intermediate temperature (28") was only 27.5 f 5.5%
(n = 6) (viability as tested by colony formation at 23") whereas the viability of cdcl3 rad9 cells grown at permissive temperature (23") was 86% (see Table 2)
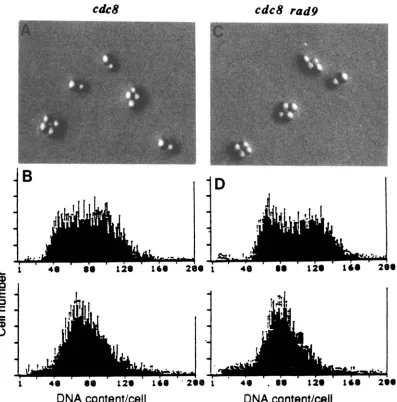
Arrest of cdc mutants in early S phase does not require the RAD9 gene: T h e efficient arrest of cdcl7 RAD and cdcl7 rad9 mutants at 38" in early S phase described above prompted a further test of whether arrest in early S phase does not in general require the
RAD9 gene. We found that cell cycle arrest in cdc8
mutants at the restrictive temperature and in hydrox- yurea-treated cells, conditions that block DNA repli- cation in early S phase, did not require the RAD9
gene (Tables 2 and 3 and Figure 4; and see Figure 1). We also examined the microtubule morphology in
cdc Mutants and the RAD9 Checkpoint 7 3
TABLE 5
Requirement for RADZ7 in cell cycle arrest of cdc mutants
Arrest
Viabilityf
morpholo- Nuclear
gyd
Strain Restrictive
Maximum genotypea temperatureb Cell cycle phase' 23" R T First cyclee 23" RT
permissive temperatureg cdc 13
cdcl3 rad17
cdc2 cdc2 rad17
cdc9 cdc9 rad 17
cdcl 7
cdcl7 rad17
cdcl7 cdcl7 rad17
cdc8 cdc8 radl 7
cdc6 cdc6 radl 7
cdc28 cdc28 radl 7
cdc7 cdcl rad 17
cdcl6 cdcl6 rad 17
cdc20 cdc20 rad17
cdcl5 cdcl5 rad 17
CDC+ radl 7
36 36 36 36 36 36 34 34 38 38 36 36 36 36 36 36 36 36 36 36 36 36 36 36 36 GP Asyn Gz Asyn G P Gn-postanaphase GP
As y n
S
S S
S
Late S
Asyn G , Gl G,-S GI-S Gz GP G2 GP Postanaphase Postanaphase
As yn
9 1 1 26 22 15 20 11 22 1 1 22 13 13 18 18 31 19 14 19 15 24 25 35 13 16 15 87 18 83 70 9 3 47 82 63 6 3 65 92 91 93 68 83 84 78 68 67 69 94 86 86 86 10 90 20 95 63 90 79 66 44 64 68 91 85 89 83 87 86 30 36 81 81 87 90 75 85 14
90 f 2.1 88 f 3.2
94 f 1.7 64 f 4.8
95 f 1.7 56
*
3.9 82 f 7.8 83 f 5.382 f 7.8 83 f 5.3
98 f 1.5 87 f 3.2
98 k 2.6 86 f 2.8 8 5 f 6.3 91 f 1.9
84 f 11.9 90 f 2.2
98 f 0.8 91 f 3.3
95 f 1.9 95 f 0.8
82 f 6.2 8 5 f 2.9 93 k 2.5
73 f 10.8 25 53 f 11.2 30 67 f 4.6 30
0.9 f 0.5 25 10 f 0.5 32 I .4 f 0.9 28 81 f 4.2 32 13 f 0.9 30 47 f 8.9 32
36 f 6.0 30 39 f 5.6 30
33 f 17.3 30 16 f 2.8 30 4.0 f 2.6 28 61 f 1.3 32 62 f 6.1 32
32 f 0.8 28 37 f 6.2 28
68 f 2.9 30 31 f 2.9 30
38 f 5.0 30
55 f 2.5 30
42 f 1.3 30 50 f 6.2 30
89 f 1.2 36
a Strains are shown in Table 1.
Restrictive temperature (RT) for cdc mutation.
Conclusions from these and previously published observations (and see Figure 1). Percentage of cells with a large bud and undivided nucleus, except cdc28 (unbudded).
Percentage of microcolonies that contain either 2 or 4 buds except for cdc28 that contain 1 or 2 buds (microcolony assay). f Mean value from 3-6 cultures; standard deviation shown.
g Maximum permissive temperature of colony formation.
both RAD and rad9 cells contained predominantly short spindles, as expected for cells arrested in medial nuclear division (BYERS and GOETSCH 1974). We con- clude that arrest of the cell cycle in early S phase when DNA replication is inhibited does not require the RAD9 gene.
Arrest of cdc6 mutants: cdc6 was the only cdc
mutant tested that showed an equivocal requirement for RAD9 for cell cycle arrest at the restrictive tem- perature; fewer cdc6 r a d 9 cells were arrested as large buds with an undivided nucleus than in cdc6 RAD cells although measurements of cell viability and first cycle arrest were similar in the two strains. In addition, cdc6 rad9 cells did have a slightly lower maximum permis- sive temperature than cdc6 RAD strains. We also meas- ured the DNA content in the cdc6 RAD strain by flow cytometry (data not shown), and found that cdc6 RAD
cells shifted to 36" for 4 hr contained less DNA than
cdc mutants that arrest in late S/G:, (e.g., in cdcl3-
arrested cells in Figure
2C,
bottom) but more DNA than that of mutants arrested in early S phase (e.g., as in cdc8 in Figure 4B) after 4 hr at the restrictive temperature. From our results we conclude that mostcdc6 RAD cells at the restrictive temperature do not require RAD9 for arrest.
Arrest of some cdc mutants in G2/M and of all cdc
mutants in other phases of the cell cycle does not require the RAD9 gene: We tested if the RAD9 gene is required for cell cycle arrest of each of 7 cdc mutants that arrest either in GI (cdc28), at the GI-S boundary
(cdc7), in postanaphase ( c d c l 5 ) , or in G2/M ( c d c l b ,
74 T. A. Weinert and L. H. Hartwell
A. R4DtX-ray B. radlZX-my c. WDtUv D. radl?,UV E. m , u v
h"nT
0 1 2 3 0 1 2 3 0 1 2 3
L
0 1 2 3 0h-rrrrr
1 2 3Time (hours)
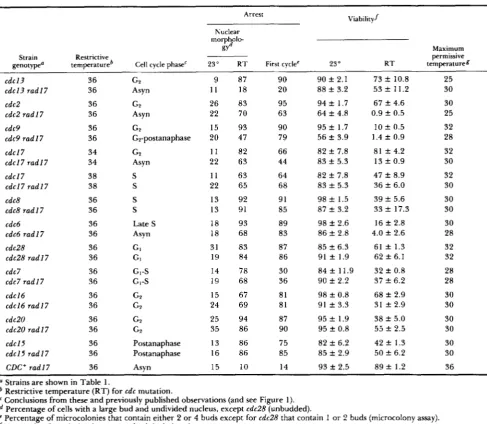
FIGURE 6.-Kinetics of cell cycle delay in X- or UV-irradiated G P cells in RAD, rad9 and radl 7 strains. G P cells were prepared by incubation with MBC, irradiated with UV or X rays, released from the MBC block and analyzed for cell cycle progression using nuclear and cell morphology. T h e percentage of cells arrested was calculated in which the daughter bud was greater than half the diameter (and usually equal to diameter of) the mother bud and with an undivided nucleus. Symbols: open circles, unirradiated; closed circles, irradiated. Dose of X rays was 4 krad and of UV was 40 J/m'. (A and C) RAD; ( B a n d D) radl7; (E) rad9. Strains: in A , RAD (7870-1-1); B, rad17 (7870-1-4); C, RAD
(7859-10-2); D, rad17 (208-7-2), E, rad9 (7859-7-4). We also observed a UV-induced delay in the two additional RAD strains tested [7870-l and 7815-6-4; Weinert and Hartwell (1988)], and a failure to delay in the other rad17 (7870-1-4) and rad9 [7815-6-4AB; WEINERT and
HARTWELL (1988)l strains tested. T h e cell viability in MBC-arrested and unirradiated cells was >80% in all strains tested.
similar in RAD and rad9 strains (Table 2). We con- clude that the arrest of these cdc mutants does not require the RAD9 gene.
R A D l 7 mutants provide a further test of the phase and signal specificity of the RAD9 checkpoint: We wanted to corroborate our findings on the role of the RAD9 checkpoint by analyzing a second mutation that, like rad9, inactivates the checkpoint. We show here first that a mutation in radl 7 results in a defect in cell cycle arrest after irradiation, and then we report the results of tests of cell cycle arrest in cdc mutants containing the r a d l 7 mutation.
We initially identified the R A D l 7 gene as essential for the RAD9 checkpoint because rad17 cells, like rad9 cells, failed to arrest following X-irradiation. T h e cell cycle response to DNA damage from X rays was performed using the methodology and rational devel- oped in earlier studies using the rad9 mutation (WEI- NERT and HARTWELL 1988). T h e response of G I and postanaphase haploid cells to X rays provides a con- venient diagnostic for monitoring cell cycle arrest after DNA damage, In wild-type haploid strains, cells in GI and postanaphase when X-irradiated arrest as large budded cells (2 budded microcolony) and as 2 large-budded cells (4 budded microcolony), respec- tively. These cells are arrested essentially irreversible (see Figure 5) as the result of a signal generated from D N A double-strand breaks that persists unrepaired because in budding yeast their repair requires an intact homologous chromosome (BURNBORG and WIL- LIAMSON 1978; GAME 1983). In mutants which have lost regulation ( e . g . , rad9), cells in GI and postana- phase when X-irradiated will not arrest but continue to divide for several generations.
Wild-type and rad17 cells were exposed to a dose of X rays that generates a few doublestrand breaks
per cell, and the response of individual cells was determined by photomicroscopy (Figure 5). We found that 83% ( n = 42) of wildtype and 3.4% ( n = 59) of rad I7 unbudded (GI) cells when X-irradiated arrested with a large bud (Figure 5). T h e unbudded rad17 cells when X-irradiated typically continued to divide for 1-3 generations, a response typical of rad9 unbud- ded cells exposed to X rays (WEINERT and HARTWELL
1988).
We also examined budded cells exposed to X rays and observed that in rad17 mutants they also failed to arrest. Budded cells consist of both doublestrand break repair-proficient GP cells and repair-deficient postanaphase cells, and their cell cycle responses to DNA double-strand breaks differ; GB cells delay, re- pair the break and resume cell division, whereas pos- tanaphase cells arrest cell division in the G:! phase in the next cell cycle. We found that 54% (n = 108) of the wild type and 4% (n = 94) of the rad17 budded cells exposed to X rays arrested as two adjacent large- budded cells (see Figure 5). Because the fraction of postanaphase cells among the budded cells in RAD and rad17 cultures were similar (43% and 49%, re- spectively, calculated from the nuclear morphologies of fixed cells from an asynchronous population of cells), we conclude that wild-type but not rad17 pos- tanaphase cells exposed to X rays undergo cell cycle arrest. Below we describe additional experimental strategies to analyse the cell cycle delay in GP cells.
cdc Mutants and the RAD9 Checkpoint 75 Delay at the RAD9 checkpoint in RAD, rad9 and
radl
7
cells afterX-
or UV-irradiation: T h e photom- icroscopic studies described above indicate that GI and postanaphase cells after irradiation require the R A D l 7 gene for arrest at the R A D 9 checkpoint in the G2 phase. T h e R A D 1 7 gene, like the R A D 9 gene, should also be required for arrest in GB of cells irra- diated in G2. T o test this prediction, cells were syn- chronized in the G:! phase by treatment with the microtubule inhibitor MBC, X- or UV-irradiated, the tubulin inhibitor was removed and the kinetics of exit from the G2 phase determined (see MATERIALS AND METHODS). We found that, as expected, X- or UV- irradiated wild-type G2 cells delayed in G2 comparedto unirradiated wild-type G2 cells [Figure 6 , A and C; see WEINERT and HARTWELL (1988, 1990)], and X- or UV-irradiated rad17 cells exhibited little delay in G2; rather they completed chromosome segregation nearly as rapidly as unirradiated rad I7 GP cells (Figure 6 , B and D). A UV-induced delay of cell division in wild-type cells has been shown previously (ESPOSITO
1968). We also found that UV-irradiated rad9 G2 cells did not delay (Figure 6E), extending our previous finding that rad9 mutant G2 cells did not delay after x-irradiation (WEINERT and HARTWELL 1988, 1990). We conclude that the checkpoint for arrest after irra- diation requires both the R A D 9 and R A D l 7 genes.
The role of the RAD1 7 gene in arrest of cdc
mutants: Finally, we tested if the cell cycle arrest of cdc mutants at their restrictive temperatures required R A D l 7. T h e cell cycle phenotypes of cdc R A D and cdc r a d l 7 mutants (Table 5) were similar to those of cdc R A D and cdc rad9 mutants (Table
2);
(1) the arrest and viability of cdc2, cdc9 and c d c l 3 at the restrictive temperature of 36" required an intact RAD17 gene; (2) the arrest and viability of cdcl7 required RAD17 at 34" but not at 38"; (3) the arrest and viability of cdc6 mutants was slightly greater in cells with an intact RAD17 gene; and (4) the maximum permissive tem- perature was lower in cdc2, cdc6, cdc9, and cdcl7 mutants that were r a d l 7 , and higher in c d c l 3 r a d 1 7 than in c d c l 3 R A D . Results from the tests for cell cycle arrest and viability of the other cdc mutants examined were similar in R A D and rad17 mutants. Finally in- cubation with hydroxyurea induced arrest efficiently in r a d l 7 mutants (Table 3).T h e phenotypes of c d c l 3 r a d 9 and cdcl3 rad17 mutants differed slightly; cdcl3 rad17 mutants shifted to 36" retained higher cell viability, were capable of
sustaining more cell divisions after shift to 36", and formed colonies at a higher maximum permissive temperature than did c d c l 3 r a d 9 cells (compare Tables 2 and 5; data not shown). Perhaps the rad17 allele retains partial function that may account for the different phenotypes.
DISCUSSION
The role of the RAD9 checkpoint in cell division appears specific: Previously we showed that rad9 mutants failed to arrest after X-irradiation or inacti- vation of DNA ligase, but unperturbed rad9 cells were viable and had cell cycle kinetics similar to that of wild-type strains. We concluded that the RAD9 check- point, when activated by DNA lesions, acts as a nega- tive regulator to arrest cells before mitosis until dam- age is repaired. Here we tested more comprehensively the function(s) of the R A D 9 checkpoint primarily by determining if cell cycle arrest of any of 12 cdc mu- tants requires the RAD9 and RAD17 genes. Our re- sults are fully consistent with the hypothesis that the R A D 9 checkpoint is activated only by DNA damage and arrests cells only in late S/G2 phase (the distinction between the late S and GS phase is addressed below). This conclusion is based on phenotypes of the cdc mutants that require the R A D 9 checkpoint for arrest and those that do not. First, the four cdc R A D mutants (cdc2, 9, 1 3 and 1 7 ) that required intact R A D 9 and RAD17 genes for arrest each arrest in the late S/G2 phase at the restrictive temperature. This indicates that the function of the R A D 9 checkpoint is phase- specific. Importantly, in each of the four cdc R A D mutants chromosome damage occurs when cell divi- sion is limited by the function of any of the mutant CDC gene products (HARTWELL and SMITH 1985>, and three of these four CDC genes encode known DNA replication enzymes (DNA ligase and DNA po- lymerase I and 111, see Figure l). These observations are consistent with the hypothesis that DNA lesions formed at the restrictive temperature in each of these cdc mutants activate the RAD9 checkpoint to arrest cell division. We suggest that the DNA lesions may be nicks, gaps or double-strand breaks formed either as a result of errors made during DNA replication by the defective replication enzymes or as gaps present at stalled replication forks in cells that have not com- pleted replication.
Additional observations suggest the RAD9 check- point is required only in arrest of cell division in the late S/G2 phase and is activated only by DNA damage. This conclusion is supported by negative evidence that the R A D 9 and RAD17 genes are not required for the cell cycle arrest of cdc mutants that arrest in either GI, GI-S, early S phase or postanaphase, or for cells arrested in S phase by hydroxyurea. T h e R A D 9 check- point is also not required for arrest of cdc mutants in the Gz/M phase in which chromosome damage does not occur when the CDC gene product is limiting for cell division, or for arrest in G2 by microtubule poisons (WEINERT and HARTWELL 1988, 1990) (Table 3, Fig- ure 6).
76 T. A. Weinert and L. H . Hartwell
other mutants that delay the cell cycle. For example,
recently GERRING, SPENCER and HIETER (1991) found that chll (ctfl) mutants delay nuclear division in the
G2/M phase and that the delay does not require the RAD9 gene. chll mutants also have an increased rate of chromosome loss but have normal levels of mitotic recombination compared to wild-type cells, which in- dicates that DNA lesions are not formed in cells limited by C H L l function. T h e delay in chll mutants may be due to another checkpoint that is required for cell cycle arrest when microtubule function or assem- bly is defective because chll mutants exhibit synthetic lethality in combination with mad2, a checkpoint mu- tant (e.g., the chll mad2 double mutant has lower cell viability than either single mutant) (LI and MURRAY
Is the RAD9 checkpoint required for arrest in late
S
phase as well as for arrest in the GZ phase? T h e RAD9 checkpoint is clearly required for arrest in the GP phase after DNA replication is completed; RAD cells, but not rad9 or rad1 7 cells, arrested with the microtubule inhibitor MBC delay nuclear division when they are X- or UV-irradiated (Figure 6) (Wei- nert and Hartwell 1988, 1990). In the cdc mutants that require the RAD9 checkpoint for arrest we are unable to determine if DNA replication has been completed. The methods employed here and in pre- vious studies (CULLOTTI and HARTWELL 197 1 ; CON- RAD and NEWLON 1983; BURKE and CHURCH 199 1) are not sufficient to determine whether 10% of the genome, for example, is unreplicated in cells that arrest at the RAD9 checkpoint. We are therefore left with two possible interpretations on the extent of DNA replication in these cdc mutants and conse- quently on when the RAD9 checkpoint acts; cdc mu- tant cells at the restrictive temperature either contain unreplicated DNA and arrest in what we call late S phase, or they contain completely replicated DNA but the sister chromosomes contain errors in the form of gaps or nicks and cells arrest in what we call the G2phase. Additional experimental approaches must be developed to determine whether the presence of any unreplicated DNA arrests cells at an earlier S phase checkpoint or at the RAD9 checkpoint.
We consider two models that describe the relation- ships between checkpoint controls and DNA replica- tion, and consider the implications of these models for the results presented here. By either model we imagine that the RAD9 checkpoint can be activated by nicks or gaps present either at replication forks or in fully replicated but defective sister chromosomes.
Model 1. The RAD9 checkpoint is required for arrest both in late S phase and in the G2 phase: This model
suggests the RAD9 checkpoint is required when most but not all chromosome sequences have been repli- cated, and the single-strand gaps or nicks at the stalled 199 1).
replication forks may be sufficient to activate the RAD9 checkpoint. This interpretation predicts that, in the experiments reported here, DNA polymerase I mutants (encoded by c d c l 7 ) at 34" do not complete DNA replication, and arrest occurs because the func- tion of DNA polymerase I is required to complete DNA replication. By the same reasoning CDC2, CDC9 and C D C l 3 gene products are required to complete DNA replication in the corresponding mutants at the restrictive temperature. This model also implies that an earlier S phase checkpoint is active when, for example, 50% of the genome is replicated (e.g., cdcl7 at 38 ") but becomes inactive when 90% of the genome is replicated. We and others have identified mutations in budding and fission yeasts that inactivate the earlier S phase checkpoint (our unpublished results; ROWLEY, HUDSON and YOUNG^^^^; Rowley, Subramani and YOUNG 1992; AL-KHODAIRY and CARR 1992; ENOCH and NURSE 1990). Perhaps sufficient "liscensing fac- tor" (LASKEY, FAIRMAN and BLOW 1989) is released only when 90% of the genome has been replicated and inactivates the earlier S phase checkpoint. After 90% of the genome has been replicated, only the RAD9 checkpoint prevents mitosis until replication and repair of any damage are completed.
Model 2. The RAD9 checkpoint is required for arrest only in the G2 phase: This model suggests the RAD9 checkpoint is required for arrest when sister chro- mosomes are fully replicated but contain nicks or small gaps due to replication errors. By this model, the RAD9 checkpoint is required only when all regions in the genome have been duplicated; replication forks have completed the elongation phase of DNA repli- cation but the defective replication enzymes have left DNA lesions that require repair. This model predicts that, in the experiments reported here, the DNA polymerase I mutants at 34" complete the elongation phase of DNA replication but leave DNA lesions. Cell cycle arrest is due to the DNA lesions whose repair may require DNA polymerase I. (If DNA polymerase I is required in this repair, the DNA polymerase I protein may exist in both a replication and a repair complex.) By the same reasoning, cdc2, cdc9 and c d c l 3 mutants also complete DNA replication at the restric- tive temperature but leave DNA lesions whose repair also requires the function of the corresponding CDC gene product.



![FIGURE 6.-Kinetics and 7815-6-4; Weinert and Hartwell (1988)], and a failure to delay in the other to was 4 krad and of morphology](https://thumb-us.123doks.com/thumbv2/123dok_us/1713568.1217974/12.591.106.488.59.223/figure-kinetics-weinert-hartwell-failure-delay-krad-morphology.webp)