Copyright 0 1996 hy the Genetics Society of America
In Viuo
Analysis Reveals That the Interdomain Region of the Yeast Proliferating
Cell Nuclear Antigen Is Important for DNA Replication and DNA Repair
Neelam S.
Aminand
ConnieHolm
Department of Pharmacology, Division of Cellular and Molecular Medicine, University of California ut Sun Diego, L a Jolla, California 92093-0651
Manuscript received May 20, 1996
Accepted for publication July 1, 1996
ABSTRACT
To identify the regions of the proliferating cell nuclear antigen (PCNA) that are important for function
in vivo, we used random mutagenesis to isolate 10 cold-sensitive (Cs-) and 31 methyl methanesulfonate- sensitive (Mms‘) mutations of the PCNA gene (POL30) in Saccharomyces cereuisiae. Unlike the Mms’ mutations, the CsC mutations are strikingly clustered in the interdomain region of the three-dimensional PCNA monomer structure. At the restrictive temperature, the CsC pol30 mutants undergo a RAD9
dependent arrest as large-budded cells with a 2c DNA content. Defects in DNA synthesis are suggested by a significant delay in the progression of synchronized pol30 cells through S phase at the restrictive temperature. DNA repair defects are revealed by the observation that CsC pol3Omutants are very sensitive to the alkylating agent MMS and mildly sensitive to ultraviolet radiation, although they are not sensitive to gamma radiation. Finally, analysis of the chromosomal DNA in pol30 cells by velocity sedimentation gradients shows that pol30 cells accumulate single-stranded DNA breaks at the restrictive temperature. Thus, our results show that PCNA plays an essential role in both DNA replication and DNA repair
in vivo.
T
0 ensure the precise and rapid incorporation of bases at a replication fork, a high degree of pro- cessivity is demanded of the DNA polymerase machin- ery. Processivity is enhanced by a DNA “sliding clamp” protein, which serves to anchor the polymerase machin- ery onto the DNA during DNA elongation (reviewed in So and DOWNEY 1992; WYMAN and BOTCHAN 1995). The sliding clamp protein in eukaryotes is the proliferating cell nuclear antigen (PCNA), a protein originally identi- fied as an S phase marker in rapidly dividing cells (BRAVO and CELIS 1980). PCNA belongs to a family of proteins that are structurally and functionally similar even in evolutionarily distant organisms. For example, the crystal structures of yeast PCNA and the subunit of Escherichia coli DNA polymerase 111 are virtually super- imposable, even though their amino acid sequences bear little resemblance to one another (KONG et al. 1992; KRISHNA et al. 1994; and reviewed in WYMAN andBOTCHAN 1995). These proteins play a vital role in en- hancing the efficiency of DNA replication, because their donut-shaped structures are suited for encircling DNA and sliding along its length (KONG et al. 1992; KRISHNA et al. 1994).
Biochemical studies have revealed a clear require- ment for PCNA in DNA synthesis (TAN et al. 1986; BRAVO et al. 1987; PRELICH et al. 1987) and repair in vitro (NICHOLS and SANCAR 1992; SHIVJI et al. 1992; MA-
Corresponding author; Connie Holm, Department of Pharmacology, Division of Cellular and Molecular Medicine, University of California at San Diego, 9500 Gilman Dr., Mail Code 0651, La Jolla, CA 92093- 0651. E-mail: [email protected]
Genetirs 1 4 4 479-493 (Octohcr, 1996)
TSUMOTO et al. 1994). A key event in in vitro replication is the loading of PCNA onto primer-template junctions of the DNA by the “clamp loader” (replication factor C) in an ATP-dependent reaction (LEE et al. 1988; LEE and HURWITZ 1990; TSURIMOTO and STILLMAN 1991; YODER and BURGERS 1991; FIEN and STILLMAN 1992; PODUST et al. 1995). The DNA-PCNA complex thus formed serves as an anchor for the polymerase
S
ma- chinery, which then elongates DNA (LEE and HURWITZ 1990; TSURIMOTO and STILLMAN 1991). In addition to being a processivity factor for pol6,
PCNA can also enhance the in vitro elongation function of another polymerase, DNA polymerase t (BURGERS 1991; LEE et al. 1991; PODUST et al. 1992). Further biochemical stud- ies show that PCNA is required for DNA repair in vitro (NICHOLS and SANCAR 1992; SHIVJI et al. 1992; MATSU- MOTO et al. 1994). For example, the PCNA-dependent polymerases appear to be required for in vitro nucleo- tide excision repair, in which -30 nucleotides are ex- cised around the damaged DNA site, followed by a “gap filling” reaction (SHIVJI et al. 1992). PCNA might also play a role in the base excision repair pathway to process abasic sites in the DNA, as shown in Xenopus lamis ovar- ian extracts (MATSUMOTO et al. 1994). In summary, the roles of PCNA have been fairly well characterized in vitro. Of course, past experience with DNA polymerase I in E. coli (DE LUCIA and CAIRNS 1969) makes it clear that it would be imprudent to assume that PCNA must have an in vivo role in DNA replication simply on the basis of its in vitro properties.480 N. S. Amin and C. Holm
synthesis, PCNA may play a role in cell cycle progres- sion. Mammalian PCNA has been found in complexes containing cell cycle factors such as cyclins and, cyclin- dependent kinases (CDKs), and it has been shown to directly bind the inhibitor of CDKs, p21, in nontrans- formed cells (XIONG et al. 1992, 1993; ZHANG et al. 1993; FLORES et al. 1994; WAGA et al. 1994). Furthermore, p21 is able to inhibit the SV40 in vitro replication reaction through its association with PCNA (WAGA et al. 1994). Perhaps the interaction of PCNA and p21 provides im- portant regulatory cues to halt DNA replication in the cell when DNA damage occurs without affecting PCNA- dependent DNA repair (LI et al. 1994). The in vivo functions of many cell cycle-related genes have been previously elucidated by the isolation of conditional mutants in yeast. Thus, creation of PCNA conditional mutants of yeast may prove valuable in deciphering its function in vivo.
The recent determination of the crystal structure of S. cerevisiae PCNA shows that the structure of this mole- cule correlates well with its ability to associate with the DNA helix. As predicted from the E. coli
0
subunit crys- tal structure, the three-dimensional (3-D) structure of PCNA consists of a homotrimeric ring with a central cavity having properties that enable it to embrace and slide along the DNA. Each monomer of PCNA has two structurally similar domains with an interdomain region made up of p sheets lying below a loop connecting the two domains (KNSHNA et al. 1994). It will be important to determine which of these regions is essential for in- teractions with replication and repair proteins such as the replication factor C, DNA polymerasesS
and e, and the cell cycle regulatory factors, cyclins, CDKs and p21. The crystal structure of yeast PCNA has facilitated the creation of deletion and/or targeted mutations in the structural gene, which have begun to provide some information on residues required for protein function. Using alanine scanning and deletion mutations, FU-KUDA et al. (1995) have identified regions of human PCNA that may be important for interaction with RFC and pol
S
in vitro; the effects of these mutations were not examined in vivo. Concentrating on the yeast protein, AWAGARI et al. (1995) also performed targeted muta- genesis. Most of their mutations had little effect when introduced into living cells. A minority confer a UV- sensitive phenotype in yeast cells; these mutations are scattered around the PCNA structure. A single cold- sensitive mutant was obtained using random mutagene- sis; this mutant appears to be defective in in vitro PCNA trimer formation, even at the permissive temperature (AWAGARI et al. 1995). Additional conditional mutants of PCNA remain to be created to probe the functional relevance of this protein in the cellular environment.To identify portions of PCNA that are important for function in vivo, we performed large scale random mu- tagenesis of the structural gene for PCNA (POL30) in
S.
cerevisiae, and we screened for phenotypes in vivo.This approach is powerful for the isolation of condi- tional mutants of a given gene, because it is not based on any prior predictions of the functional importance of specific residues. Using this method, we identified several mutations that cause sensitivity to the DNA alkyl- ating agent MMS, and we find that they are scattered around the PCNA structure. In addition, we find a strik- ing clustering of cold-sensitive mutations in a single region of the PCNA molecule between the two domains of the monomer. Through genetic, cytological, and bio- chemical studies we show that PCNA is required for efficient DNA replication and repair in vivo.
MATERIALS AND METHODS
Strains and media: All the pol30 yeast strains used in this study have an S288C background and were created using methods outlined in SHERMAN et al. (1986). They are listed in Table 1.
For the integration of pol30 alleles into the genome, we used the LEU2 gene as a selectable marker and verified the integra- tion of the alleles at the POL30 locus by Southern analysis; strains bearing integrated pol30 alleles were used for all physio logical experiments. First, a SmaI-Hind111 (blunt) fragment of plasmid pJ252 (LEU2) (JONES and PRAKASH 1990) was s u b cloned into the StuI site of plasmid pCH1565 (POL30), to create plasmid pCH1572 (LEUZ.POL30), in which LEU2 is inserted upstream of the coding sequence of POL30. Plasmid pCH1565 contains a 3-kb yeast genomic sequence including the POL30
gene, and the StuI site is located 528 bp upstream of the POL30
ATG site. Plasmid pCH1572 was digested completely with MluI,
and partially with SacI to replace its POL30 sequence with MluI-
Sac1 fragments of plasmids pCH1594pCH1601 containing
pol30100 to pol30107 alleles, respectively. This resulted in the creation of plasmids pCH1573 to pCH1580 (LEU2.po130100 to
LEU2.po130-107). The POL30 and pol30 genes were then inte- grated into the genome by transforming strains CH594 and/ or CH595 with the SacI fragments from plasmids pCH1572 to pCH1580 (LEU2.po130-100 to LEU2.po130-107). Southern analy- sis (SOUTHERN 1975) confirmed the correct integration of the alleles in the genome. The LEU2.POL30 strain CH2165 shows no phenotypes different from the parental strain, CH595
(POL30). The Cs- alleles conferred a stronger phenotype when integrated into the genome compared to when they were plas- mid-borne, possibly due to differences in expression levels.
Rho' strains were used for flow cytometq and were created by growing the rho' strains CH2165 (POL30), CH2161 (Po130
104) and CH2163 (pol30103) twice in 25 pg/ml ethidium bro- mide in YEPD to stationary phase (SHERMAN et al. 1986). These cells were plated for single colonies on YEPD and tested for their ability to grow on YEP plates containing 2% glycerol (a nonfermentable carbon source). In addition, fluorescence mi- croscopy of 4', 6'diamidincl2-phenylindole (DAP1)stained cells showed the absence of mitochondrial DNA in the rho" strains.
Cells were grown inYEPD (rich medium) and SD (synthetic dextrose or selective) medium. YEPD is 1% yeast extract, 2% bactopeptone, 2 % dextrose, with or without 2% bacto-agar; SD medium is 0.67% yeast nitrogen base, 2% bacto-agar, and 2% dextrose. For synthetic complete (SC) medium, 20 mg of uracil, adenine, and tryptophan, 30 mg of lysine, and 60 mg of leucine were added to 1 liter of SD medium.
Molecular biology: All molecular biology techniques were performed as described in Current Protocols in Molecular Biology (AUSLJBEI. et al. 1988).
In Vivo Analysis of PCNA
TABLE 1
S. cereuisiae strains used in this study
481
Strain Genotype" Source
CH1887 CH2027 CH2097 CH2 134 CH2 160 CH2161 CH2 162 CH2 163 CH2164 CH2 165
MATa ade2-101 leu2Al bs2-801 t,plAl urc MATa leu2 ura3-52 RAD9: : L E U 2
MATa ura3 trpl m c l - 1
MATa leu2 ura3-52 trpIA63 POL30:: LEU2 MA Ta led-3, I1 2 ura3-52 LEU2.po130-107 MA Ta leu2-3,112 ura3-52 I;EU2.pol30-l04' MA Ta leu2-3,112 ura3-52 LEU2.pol30-102 MA Ta leu2-3, 11 2 ura3-52 LEU2.pol30-I07 MATa leu2-3,112 ura3-52 LEU2.po130-104' MATa leu2-3, 11 2 ura3-52 LEUZ.POL3g
CH325 MATa lys2-801 ura3-52 his4-539 top2-4 HOLM et al. (1985)
CH533 MATa adel ade2 lys2 ural his7 tyrl gall cdc9-l HARTWELL et al. (1973)
CH552 MATa adel ade2 uraI his7 tyrl gall cdc28 HARTWELL et al. (1973)
CH594 MATu ku2-3, 112 ura3-52 D. BOTSTEIN
CH595 MA Ta leu2-3, I 1 2 ura3-52 D. BOTSTEIN
CH774 MATa adel ade2 his7 lys2 tyrl ural cdcl7-2 HARTWEI.I. et al. (1973)
CH1639 MATa leu2 ura3 RAD52: : URA?' SPELL et al. (1994)
CH1807 MATa ura3-52 his4-539 lys2-801 cdc44-5. URA3 HOWELL et al. (1994)
CH1886 MATa ade2-101 leu2Al lys2-801 t r p l A l ura3-52 CLlC44.URA3' This laboratory
13-52 cdc44-8. URAY This laboratory
MCALEAR et al. (1996) T. WEINERT
[pCH1511 (POL30 URA3)I" This study This study This study This study This study This study This study
CH2250 MATa leu2 ura3-52 LEU2.po130-1 03 RAD9: : L E U 2 This study
CH2252 MATa leu2-3,112 ura3-52 LEUZ.po130-104 [rho'] ' This study
CH2253 MATa leu2-3,112 ura3-52 LEU2. POL30 [ rhoq This study
CH2254 MATa leu2-3,112 ura3-52 LEU2.po130-103 [rho"] This study
CH2255 to CH2270 MATa leu2 ura3-52 trplA63 POL3O::LEU2 [pCH1594 to pCH1609 This study
(pol30100 to po130-115 TRP1)Ih
Plasmids as well as rho' strains indicated by [ 1.
*
:: is the symbol used for disruption of a gene with the URA3+ or LEU2+ genes as markers.'These strains have the URA3+ or LEU2' genes adjacent to a described gene but do not disrupt the gene.
gene (MUHLRAD et al. 1992) and the plasmid shuffle technique (BUDD and CAMPBELL 1987) were used to create new mutants of yeast PCNA. The POL30 gene in plasmid pCH1565 was muta- genized by PCR amplification in 24 independent reactions; half the reactions contained 1 PM MnC12. Plasmid pCH1565specific primers were used to ampllfy the POL30 gene using Perkin Elmer taq polymerase. The entire coding region was mutagen- ized by using a T3 primer (5"GGGAAATCACTCCCAATTA-3') and the primer 5'CCTGCGCGTAACCACCAG3'. The ampli- fied DNA containing the mutagenized POL30 gene, and a Sac1
vector fragment of pCH1565, were then transformed into strain CH2134 (POL3O::LEU2 [pCH1511(POL30, URAto allow for ])j recombination during transformation (MUHLRAD et al. 1992). The transformants were replica plated onto 5-fluoroorotic acid (5FOA)containing plates to select for the loss of pCHl511
(PO1230), and they were simultaneously screened for phenotypes at 37", 14", or in the presence of 0.017% MMS, and 0.04 M HU. We confirmed that the phenotypes conferred in each mutant were due to the pol30 gene by isolating the plasmids canying the mutant alleles (ROBZYK and KASSIR 1992), and retesting the phenotypes they conferred after transformation into the parent strain, CH2134 (POL3O::LEU2 [pCHl5ll{POL30, URA3]]).
From the 8000 colonies screened, we obtained a total of 42 Cs- and/or Mms" mutants; in addition, 1.3% of all colonies screened were null mutants. Twenty-eight of the 42 mutants recovered were produced from mutagenesis reactions lacking Mn2+. We used dideoxy sequencing (Sequenase reagent kit from US Bio- chemical Corp.) to identify the mutations in 10 Cs- and six Mms' pol30 genes. Mutations in the coding region of POL30 are listed in Table 2. We also found additional mutations, dele- tions (-) or insertions (+) of bases in the 5' or 3' noncoding
regions of POL30 in some of the mutant genes. They are as follows: A776G in po130100,
-bnx
and -&35 in p0130-102, T-,%Cand A_,,G in pol30-107, -b47 in po130108, in po130109,
b u t i T in pol30110, Tw& in pol301 11, && in po13O-112, T,& and Ayq7G in pol3n113, -A- 1x5 and A- in p0130-115; number- ing is from the A of the ATG start codon.
Synthetic lethality: To obtain double mutants of pol30 with several cell cycle-related mutant genes, we crossed the cold-sensi- tive strains CH2160 (po130103) and CH2164 (poZ30104) with the heat-sensitive strains CH325 (top2), CH557 (cdc28), CH533
(cdc9), and CH774 (cdcl7). For the pol30 X cdc44 cross, we crossed the coldsensitive strain CH1807 (cdc44-5) or CH1887
(cdc44-8) (these strains carry identical cdc44 mutations) with each of strains CH2166 (POL?O), CH2158 (po130101), CH2161
(po1?@104), CH2159 (pol?0-100), CH2162 (pol3@102), CH2163
(po130-103), CH2168 (po130107), CH2169 (po130-105), CH2170
(pol3@106), and CH2171 (poW0-108). The diploid strains were sporulated, and tetrads were dissected and allowed to germinate at 30" in the case of the cdc44 X pol30 cross or at 25" for the remaining crosses. Between 33% and 100% of the expected double mutants were observed at 25" for each of the pol30crosses with tOp2, cdc9, cdcl7, and cdc28 in six to 14 tetrads. No double mutants were obtained in any of the crosses between cdc44
strains and pol30 strains. Between nine and 21 double mutants were expected in these latter crosses.
Cell and nuclear morphology: To observe the cellular and nuclear morphology of POL30 and pol30 cells at the permis- sive and restrictive temperatures, strains CH2165 (POL30),
482 N. S. Amin and C. Holm
TABLE 2
Mutations in the coding region of POL30
Allele Nucleotide change(s)" Amino acid change(s) Phenotype" Location'
pol?o-lOo
pOz?@lol po1?0-102 p013o-1
or'
pol?0-108
pol?o-l I 2 pOl?o-ll? p01?0-109
pol?O-114
po1?0-115
Csf. Mms'
Cs-, Mms'
Ck', Mms'
Cs-, Mms'
Cs-, Mms' Cs+, Mms'
Cs-, Mms' Cs-. Mms'
Cs-, Mms' Cs", Mms'
Cs+, Mms'
Cs+. Mms'
Cst. Mms'
Cs+, Mms'
P.
c:
P,
c
P,c
P,c
P,
c
P,c
P, C P.c
P, C P P P
P
P
"Numbering of the bases starts at the A (No. 1 ) of the first ATG of the POL?O open reading frame.
"pol?O-105 is only weakly MMS-sensitive, and pol?O-107 has a weak Cs- phenotype.
'Phenotypes were determined for all mutations when carried on a plasmid (P). In addition, phenotypes conferred by an integrated, chromosomal (C) allele were also determined for some alleles. In general chromo- somal alleles had slightly stronger phenotypes than plasmid-borne alleles.
"Because k 4 G alone confers cold sensitivity in pol?O-lOl, we assume that R& also confers cold sensitivity in the double mutant po130-IO?. The D2*?N mutation appears to improve growth somewhat at permissive temperature.
or 14" (10.5 hr). Cells were then fixed in 3.7% formaldehyde (Fisher Scientific) for at least 3 hr before washing and sonicat- ing. The percentage of cells with single, small or large buds was scored for each cell type. The fixed cells were also ana- lyzed for nuclear staining using DAFT and the antibleach dia- zabicyclo[2.2.2]-octane (DABCO). Between 200 and 700 cells were scored for nuclear and cellular morphology.
Flow cytometry: To determine the DNA content of expo- nentially growing cultures at 35" and 14", the rho" strains CH2253 (POL?O), CH2252 (pol?O-I04), and CH2254 (p0130-
IO?) were grown to log phase at 35". The cultures were split and aliquots were incubated at 35" for 2.5 hr, or 30" for 3 hr, or 14" for 24 hr. To prepare them for flow cytometry, samples were sonicated, diluted and prepared for microfluorometric analysis by staining with propidium iodide, as described (HUT-
TER and EIPEI, 1979). A Becton Dickinson fluorescence acti- vated cell sorter was used to determine the DNA content of the cells in each sample. To observe the DNA content of cells released from G1 arrest into the restrictive and permissive temperatures, rho" strains were grown to log phase in pH 4.0 YEPD at 35", and then treated with 5 pg/ml of a factor (Sigma) for 2.5 hr at 35". The cells were observed under the microscope to confirm cell cycle arrest in G1. The a factor was then washed away, and the cells were resuspended in pH 4.0 YEPD. Half of each culture was incubated at 35" and the other half was incubated at 14". Samples were collected over time and prepared for flow cytometry. Between 10,000 and 20,000 cells of each type were analyzed for their DNA content in these experiments.
S phase checkpoint assay: To examine the effects of pol30 mutations on the S phase DNA damage checkpoint, we used the MMS asynchrony experiment described in Pl\uL,ovlcIi
and H~RrnT1.1. (1995). Strains CH2165 (POL?O), CH2161
(pol?O-104) and CH2097 ( m e c l - I ) were grown to log phase at the semipermissive temperature of 30" in YEPD medium, and then treated with 0.015% MMS. One milliliter samples were collected for viability and flow cytometry analysis at 0, 2, 5, and 8 hr time points after MMS addition. The samples were sonicated and diluted. For viability assessment, the sam- ples were plated on YEPD plates and incubated at 35" for 20 hr, and the number of viable microcolonies were scored as having > I 5 cells under the microscope. The viability of
POL?O cells after 5 hr in MMS was 57% compared to pol?O
cells, which was 3%. To analyze the DNA content of cells, we prepared cells for flow cytometry as previously described
(HIJTTER and EIPEI. 1979).
First-cycle arrest experiment: Cell cycle progression was monitored at the restrictive temperature in plate assays as described in WEINERT and HARMLL (1993). Strains CH2165
( P O L N ) , CH2163 (poZ?O-lO?), CH2027 (rud9), and CH2250
(poZ?O-lO? rud9) were grown to log phase at 35". The cells were sonicated, diluted and plated on prechilled or pre- warmed YEPD plates, and incubated at 14" and 35", respec- tively. The number of cell cycle divisions was monitored over time by scoring the number of cell units (1, 2 , 3 to 4, and >4) in each microcolony for up to 9 hr at 35" or 33 hr at 14". The percentage of cells that have more than four cell bodies at 14" after 33 hr was normalized for the number of colonies that have more than four cell bodies after 9 hr at 35", due to the reduced viability (54%) of strain CH2250
In Vivo Analysis of PCNA 483
Viabfity assay: We examined viability using a method de- scribed by WEINERT and HARTWELL (1993). Strains CH2165
(POL?O), CH2163 (p0130-103), CH2027 (rad9), and CH2250
(po130-103 rad9) were grown to early log phase at 35" in liquid YEPD. The cultures were then shifted to 14", and aliquots were collected approximately every 10 hr for 40 hr. The sam- ples were sonicated, diluted, and plated on YEPD plates. After 20 hr of incubation at 35", the viability of each cell type was determined by scoring the number of microcolonies with more or less than 10 cells under the light microscope.
Spontaneous mutation frequencies: We determined the mutation frequencies in POL?O and poUOstrains using a cana- vanine-sensitivity assay ( M C ~ E A R et al. 1994). Two to three independent cultures of strains CH2165 (POL30), CH2163
(poZ30-103), and CH2161 (po130-104) were grown to mid-log phase at the semi-permissive temperature of 30" in liquid YEPD. The cells were sonicated, and appropriate dilutions were spread on SD plus 20 mg/l uracil plates and on SD plates containing 60 mg/l canavanine and 20 mg/l uracil. The plates were incubated at 30" for 4 days, after which the number of viable colonies per plate was counted, and the number of canavanine-resistant colonies per
lofi
viable cells plated was calculated.Sensitivity to DNA damaging agents: The sensitivity of
POL30 and poU0 strains to different DNA damaging agents was assessed using W radiation, gamma radiation and MMS. Strains CH2165 (POL30), CH2163 (pol30-103), CH2161
(pol30-104), CH2027 ( r a d 9 ) , and CH1639 (rad52) were grown exponentially at 35", diluted, and plated on YEPD to obtain a density of 300 cells/plate. The plated cells were then exposed to 254 nm UV radiation doses from 0 to 200 J/m' using a 1800 Stratalinker (Stratagene), and incubated at 35" for 2 days or 25" for 4 days before the number of viable colonies was scored. In the case of gamma irradiation, cells diluted in water were exposed to 0-20 Krad doses from a CslS7 radiation source before plating. For assaying MMS sensi- tivity, cells diluted in water were incubated in 0.3% MMS for 0-40 min. After exposure to gamma radiation or MMS, cells were plated on YEPD medium and incubated at 35" for 2 days or 25" for 4 days before the number of viable colonies was scored. Viability curves were generated in three independent experiments.
Velocity sedimentation gradients: A previously outlined method was used to culture cells and isolate genomic DNA for analysis in DNA sedimentation gradients (BUDD et al. 1989; MCALEAR et al. 1996). To restrict our analysis to chromosomal DNA from the different cell types, we created rho" strains, which lack mitochondrial DNA (SHERMAN et al. 1986). Strains CH2252 [pol30 (rho")] and CH2253 [POL30 (rho")] were grown in YEPD at 35" to log phase at a cell density -5 X IO6 cell/ml. To 500 pl of cells resuspended in SC media, 60 mCi of 5,6"H uracil (35 Ci/mmol, NEN Dupont) or 10 mCi of
Curacil (60 mCi/mrnol, NEN Dupont) were added. The cells were labelled at 35" for -6 hr. For the analysis of DNA from cells at the restrictive temperature, the samples were shifted to 14" for 24 hr after a 6-hr labeling period at 35". Those cells exposed to MMS were first radio-labeled at 35" for 6 hr, followed by the incubation of cells in 0.3% MMS (Sigma) for 5 min. The samples were washed twice in SC media before allowing cells to recover at 35" for 3 hr, or 24 hr at 14". [The viability of cells treated with 0.3% MMS under experimental conditions is 10% for CH2252 (po130-104) and 60% for CH2253 (POL30).] DNA was isolated and centri- fuged in 15-3076 neutral or alkaline gradients as described
( M G ~ L W R et al. 1996).
For the comparison of the DNA profiles of two different strains, we labeled DNA from one strain with 'H-uracil and the other with '*C-uracil, and sedimented the DNA from both I4
strains in the same gradient. The results were unchanged when the strains were oppositely radio-labeled. All profiles shown are plotted with fraction 1 representing the top of the gradient and fraction 20 representing the bottom of the gradient. The number-average molecular weight of the DNA was calculated as described (GREEN et al. 1974; "EAR et al. 1996). For the neutral gradients, the number-average mo- lecular weights are 9.1 X
lo8
for strain CH2253 [POL30(rho')], and 9.7 X 10' for strain CH2252 [poZ30-104 (rho')], when both strains were grown at 35"; for alkaline gradients the values are 2.7 X
lo8
for strain CH2253 [POL30 (rho')], and 2.7 X 10' for strain CH2252 [pol3@104 (rho')], when both strains were grown at 35". At 14", under neutral condi- tions, strain CH2253 [POL30 (rho')] has a number-average molecular weight of 1.2 Xlo9
and strain CH2252 [pol?@104(rho')] has a value of 1.1 X 10'. We found that differences in number-average molecular weight of 10% or greater were reproducible and significant.
RESULTS
Cold-sensitive pol30 mutations cluster in the 3-D
structure of the protein: To identify regions of PCNA that are important for function in vivo, we used random PCR mutagenesis and the plasmid shuffle technique
(BUDD a n d CAMPBELL 1987; MUHLRAD et al. 1992) to create conditional pol30 mutants. Plasmid pCH1565 bearing the mutagenized POL30 gene was transformed into a yeast strain containing both a chromosomal dis- ruption at the POL30 locus and plasmid pCH1511
(POL30 URA?). Incubation on 5-FOA medium selected for the loss of the POL30 URA? plasmid, and the poten- tial pol30 mutants were screened for sensitivity to heat
( 3 7 7 , cold (14"), 0.04 M hydroxyurea (HU) , a n d
0.017% MMS ( a DNA alkylating agent). From the 8000 colonies screened, 10 cold-sensitive (Cs-) mutants were recovered; all of these were also sensitive t o MMS. In addition, we recovered 31 mutants that were sensitive only to MMS. Neither HU-sensitive n o r heat-sensitive mutants were produced in this screen. For each of the mutants, we confirmed that the mutant phenotype was conferred by the plasmid-borne pol30 gene by isolating the plasmid and transforming it once again into yeast. Analysis of the nature of the pol30 mutations revealed that most of the amino acid changes in the Cs- mutants reside in the C-terminal portion of PCNA. We used dideoxy sequencing to identify the mutations in six Mmss mutants and 10 Cs- mutants (Table 2). We found identical mutations in po130-103and pol30-111 and inde- pendently, in po130-101 a n d p0130-110. Each pair of mu- tants was derived from the same PCR pool, so we assume that members of each pair are identical by descent. Mutations in 15/16 of the alleles occur in conserved or similar residues of known eukaryotic PCNA sequences. Furthermore, six of the eight Cs- mutants have amino acid changes in the C-terminal portion of PCNA, a n d three of these occur in the sequence LAPK in the C- terminal tail. This tetrapeptide region is the longest stretch of conserved amino acids in
S.
cereuisiae, S. pombe,484 N. S. Amin and C. Holm
FIGLIRE 1.-Predicted locations of amino acids altered hy C- and Mms' PCNA mutations. a-carhon traces of the yeast PCNA trimer are shown. Superimposed on this tracing is a space-filling representation of those amino acids that are altered in the Cs-
(A) and Mms' (B) mutants. (A) Cold-sensitive mutations are clustered in the interdomain region of each PCNA monomer. (B) Mutations conferring sensitivity to MMS are scattered around the PCNA structure. Because the specific amino acid changes conferring Mms' in the mutants j10130113 and j~o13O115 are unknown, hoth mutations are depicted in each case. In hoth A and B, the location of each of the amino acid changes is shown in all three monomers of the PCNA trimer. The Insight I1 program was used to create this illustration, using coordinates of the amino acids of yeast PCNA (KRISHNA el nl. 1994).
Mapping the mutations to the crystal structure of PCNA reveals that the Mms" mutations are scattered, but the Cs- mutations are markedly clustered. The mu- tations conferring only MMS sensitivity are distributed around the PCNA molecule (Figure 1B). The positions of the residues conferring MMS sensitivity include the following: (1) the region around the monomer-mono- mer interface (pol30-105, po13O-113, and the K164R mu- tation of pol3@115), (2) the interdomain region of the monomers (po130102, po130-112, and the K242E muta- tion of pol3@115), and (3) the inner periphery of the trimeric ring ( p o 1 3 0 1 1 4 ) . We infer that any small de- crease in the activity of PCNA probably causes a lowered tolerance for MMS. In contrast to the Mms' alleles, the Cs- alleles (which are also sensitive to MMS) exhibit striking clustering (Figure 1A). The eight independent Cs- alleles (poMO-100, po130-101, p0130-103, $10130-104, po130-106, p0130-107, p0130-108, and p0130-109) all cause amino acid changes in the interdomain region of the PCNA monomer, which lies directly beneath a loop connecting the two monomer domains. The clustering of the Cs-conferring mutations in the interdomain portion of PCNA suggests that this region is functionally important in vivo.
Po130 mutations exhibit synthetic lethality with cdc44 mutations: In vitro studies reveal clearly that PCNA in- teracts with RFC, and many PCNA mutations have been identified that suppress cold-sensitive rfcl (cdc44) muta- tions (MCALEAR et al. 1994; AWAGARI et al. 1995). To determine which of the new PCNA mutations might
have this property, we crossed pol30 strains with cdc44
strains, sporulated the diploids, and examined the h a p loid progeny. We were surprised to find that when the identical mutations cdc44-5 or cdc44-8 were combined with po130100, p0130-101, po130102, p0130-103, p o l 3 0 104, po130-105, po130106, po130107, or po130108, the double mutant was inviable at 30" in every case (data not shown; only these eight pol30 alleles were tested). In control experiments, we showed that there is no synthetic lethality in pol30 cdc17, pol30 cdc28, pol30 cdc9
or pol30 fop2 double mutants. Together with earlier re- sults demonstrating that many pol3Omutations suppress
cdc44 mutations ( M C L F A R et al. 1994; AWACARI et al.
1995), these results confirm the intimate interaction between PCNA and replication factor C, and they reveal that it is possible to create pol30 mutations that either enhance or hinder this interaction.
PCNA mutants arrest with a G2/M DNA content at
the restrictive temperature: The cold-sensitive pol30
mutants exhibit a cell division cycle phenotype (Cdc) at the restrictive temperature. For physiological studies, we chose the two Cs- pol30 mutants with the most nor- mal growth characteristics at the permissive tempera- ture. Both strains CH2163 ( p o l 3 0 1 0 3 ) and CH2161
14"
In V i r m Analysis of PCNA 48.5
B
3
5"
3 0"
1 4"
l?h!!Lkk
:h...
,L(Ll](-
:kp[..
,FIGL~RE 2.-Nuclear morphology and DNA content of po/?O cells. ( A )
Nuclear staining o~l'f~l.30antl Po130 cells. Strains CH2lfi:i (fOI,30), CH2I63 (polXkl03) and CH2161
(jm/?@lO4) were incubated at the restrictive temperature (14") for
10.5 h r ( 1 ..5 generation time) before staining with DAM. (R) DNA con- tent h i s t o p m s of' ffX.30 anrl j d ? 0
cclls at X", 30" and 14". Rho" strains CH2253 (fO1,?0), CH22.54 (po/?@
lo?) and CH22.52 (j10/?0104) were
$
2
incubated at 3.5" (2 hr), 30" (3 hr)Q or 14" (24 hr), and thc DNAcontent
0
\rrts assaved bv flow cytomeq. In the
h i s t o p m s shown here, the x axis represents the relative DNA content
anrl the y axis represents cell num- ber. T h e small peak at the left in some profiles is clcrived from a small number of lysed cells.
2
0Q
1c 2c 1c 2c 1c 2c
ture (14") for 10.5 h r (1.5 generation time). Mhereas only 5% of POL30 cells incubated at 14" are large-bud- ded, 80% of pol30-103, and 72% of po130-104 cells are large-budded. Nuclear staining with DAPI showed that these large-budded cells have a single nucleus at the neck of the bud (Figure 2A). Such a morphology is consistent with that reported by AYKAGARI PI al. (1995) with a single Cs- pol30 mutant. This uniform terminal morphology reveals that pol30 mutants exhibit a cell division cycle phenotype, and it suggests that PCNA is required at a specific time during
S
phase o r mitosis.To address the previously unanswered question of whether pol30 cells arrest at the restrictive temperature with partiallv o r f d l y replicated DNA, we analyzed the DNA content of mutant and wild-type cells by flow cy- tometry (HUTTER and EIITL 1979). Exponentially grow- ing cultures were split, and aliquots were shifted to 35"
o r 14" for 1.5 generation times. Microfluorometric anal-
ysis (FACS; Figure 2R) of these asynchronous cultures showed that 1'01.30 and pol30 cells have similar FACS profiles at 35"; -40% of the cells have a I C DNA con- tent, and 60% have a 2c DNA content. After incubation at 14", however, virtually all of the po/30 cells exhibit a 2c DNA content. Thus, pol3Omutants undergo cell cycle arrest at the restrictive temperature after completing the bulk of DNA replication.
Progression through S phase is retarded in pol30 mu-
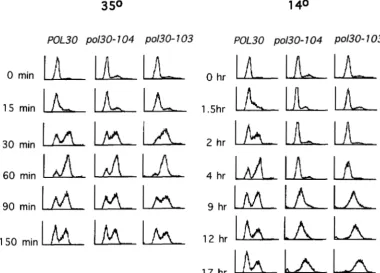
tants: Do /1o/30 cells have a defect that prevents the transition from G2 to mitosis, or does a defect in DNA synthesis cause their arrest? To begin to address this question, we followed the kinetics of DNA replication in
synchronized cultures of po/30 and POL30 cells. Strains CH2254 (po/30-103), CH2252 (po130-104), and CH2253 (POL30) were synchronized in G1 using cr factor, and after the cr factor was washed away the cells were shifted to the permissive o r restrictive temperature. As the cells progressed through the cell cycle after release from arrest, we monitored their DNA content by flow cytome- try (Figure 3 ) . The kinetics of DNA replication were similar in all cell types at the permissive temperature
(35"). At the restrictive temperature, however, DNA syn- thesis was much slower in pol30 mutants than in POL30 cells. In POL30 cells, most DNA synthesis is accom- plished between 1.5 and 4 h r after release from cr factor; in po/30cells, little DNA synthesis is seen until 4 h r after release from a factor. Thereafter, progression through S phase is slow in the mutant cells, and after 17 hr at 14", they arrest with a G2/M DNA content as previously seen with asynchronous cells. This result clearly shows the requirement of PCNA for efficient DNA replication
in vivo.
486 N. S. Amin and C. Holm
3
50
1 40
POL30 ~0130- 7 04 ~0130- 703 POL30 ~0130- 7 04 P0130- 703 0 min
kkk
O h r k k k1 5 min
k
ks-
1.5hrla,
30
minkkk
Z h r k k k60 min
kku
4 h r u k k90 min ~~~ 9 h r h k h
150 min
IM
kk
1 2 h rkkh
17 hr
L
4
!
!
L
L
L
h
FIGURE 3.-Progression of pol30 cells through S phase at 14". Strains CH2253 [POL30 (rho")], CH2254 [pol3@103 (rho")], and CH2252 [po130-104, (rho")] were arrested in G1 by treatment with a factor at 35" for 2.5 hr. After the a factor was washed away, the cultures were released to the permissive (35") or restrictive temperature (14"). The DNA content of the cultures was monitored over time by flow cytometry. In the histograms shown here, the x axis represents the relative DNA content and the y axis represents cell number.
MMS, we monitored the DNA content of pol?0 mutants by flow cytometry before and after treatment with 0.015% of MMS (Figure 4). As expected, MMS treat- ment causes POL30 cells to traverse S phase slowly, whereas mecl cells traverse S phase rapidly. In pol30 mutants, S phase is retarded at least as much as it is in
POL30 cells. Thus, the regulatory mechanisms control-
ling the rate of DNA replication appear to be intact in pol30 mutants.
The $0130 arrest-phenotype is dependent on the
RAD9 checkpoint: The slowing of DNA synthesis ob- served in pol?0 mutants in the absence of MMS at the restrictive temperature suggests that pol?0 mutants may undergo cell cycle arrest due to defects in the DNA. To determine whether the DNA damage checkpoint gene RAD9 plays a role in the cell cycle arrest of pol?0 mu- tants, we constructed a pol?0 rad9double mutant strain. Cultures of strains CH2165 (POL?O), CH2163 (polj'0-
IO?), CH2027 (RAD9::LEU2) and CH2250 (Po1?0-10?
RAD9::LEU2) were grown exponentially before the
cells were shifted to permissive or restrictive tempera- ture. After 33 hr at the restrictive temperature of 14", 100% of the viable POL30 RAD9 cells and 93% of the viable POL30 rad9 cells formed microcolonies with greater than four cell bodies; only 2% of viable po130- 103 RAD9cells formed such microcolonies. In contrast with the pol?0 RAD9 result, we found that
72%
of the viable poW0 rad9 cells formed microcolonies withgreater than four cell bodies. This result is clearly con- sistent with a RADPcheckpoint-mediated arrest. The second characteristic of a RAD9 related arrest is that viability is expected to drop in a pol30 rad9 strain incu- bated at the restrictive temperature. When we exam- ined the viability in strain CH2250 (pol?@I0? rad9) at the restrictive temperature, we found a rapid loss of viability in the double mutant (Figure 5). Thus, it ap- pears that the RAD9 gene product causes a cell cycle arrest in poU0cells due to the presence of DNA damage. Similar results were obtained with a p01?0-104 m c 2 - I double mutant (N. A M I N and C. HOLM, unpublished
data).
In Vivo Analysis of PCNA 487
0 hr.
2 hr.
POL30
~0130-
104
mec 7
7 t . I L
FIGURE 4.-Assessment of S phase checkpoint integrity in pol?O cells. Exponentially growing strains CH2165 (POL?O),
CH2161 (poZ?0-104), and CH2097 (mecl-1) were treated with
MMS at a final concentration of 0.015%. Flow cytometry was used to monitor the DNA content of the asynchronously divid- ing cultures incubated in the presence of MMS over time. The x axis represents the relative DNA content and the y axis represents cell number.
canr colonies per
lo6
viable cells plated for po130-10? and 122.4 ? 74.0 for po130-104) than in the POL30 strain (0.1 ? 0.1 canr colonies per lo6 viable cells plated). These results are consistent with the previously observed elevation in spontaneous mutations in other pol30 mutants (AWAGARI et al. 1995). The high sponta- neous mutation frequencies in pol30 mutants suggest that there may be high levels of DNA damage in pol?0 strains.poU0 cells accumulate single-stranded DNA breaks:
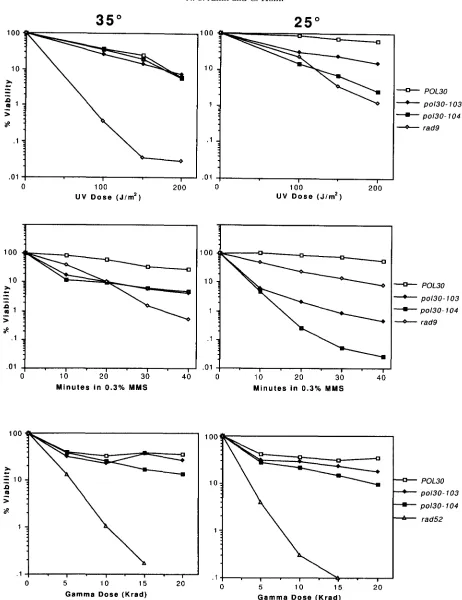
To determine whether elevated levels of DNA damage might be due to a deficiency in the ability to repair damaged DNA, we tested the sensitivity of pol?0 strains to different types of DNA damaging agents. Cultures of strains CH2165 (POL30), CH2163 (pol?0-103), and CH2161 (poMU-104) were grown exponentially at the permissive temperature. Samples were treated either with varying doses of UV radiation or gamma radiation, or for varying times in MMS. The viability of cells was then determined on plates after growth at permissive (35") or semi-permissive temperature (25") (Figure 6). Although Po130 cells were not sensitive to gamma radia- tion at either 35" or 25", they were 10-10' fold more sensitive to MMS than were the POL?O control strains after incubation at both temperatures. In contrast, the
* PO130
-
PO130"---t rad9
--" ~0130 rad9
"I . I . , . I . > 0 5 1 0 1 5 20
Hours at 14'
FIGURE 5.-Viability ofpo1?@10? rad9strain at 14". Cultures ofstrains CH2165 (POL?ORAD9), CH2163 (poWO-l0?RAD9), CH2027 (POL30 rad9), and CH2250 (po1?@10? rad9) were grown to log phase at 35" and then shifted to the restrictive temperature (14"). Samples were collected over time and plated for viability at 35" (described in MATERIALS AND METH- ODS). For each strain, the percentage of viable cells at each time point was compared to the percentage at zero time; the ratio between the two is plotted here.
p o U 0 strains were similar to POL30 strains in their sensi- tivity to UV radiation at 35", but they were somewhat sensitive (six- to 20-fold) to W light compared to wild- type cells at 25". These results suggest that PCNA may have little involvement in the repair of double-stranded breaks in vivo, but they suggest that PCNA plays some role in the recovery from UV-induced damage, and a key role in the recovery from MMS-induced damage.
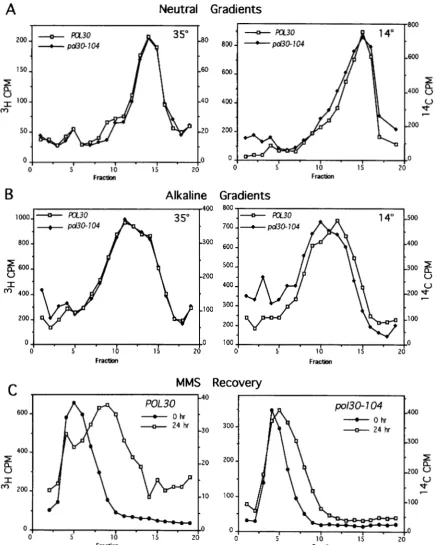
To determine whether pol?0 strains exhibit DNA damage in the form of single- or double-stranded breaks, we used velocity sedimentation to analyze DNA from pol30 and POL30 cells grown at the permissive or restrictive temperature. Single-stranded DNA breaks are revealed in alkaline gradients, and double-stranded DNA breaks are visualized in neutral gradients (BUDD et al. 1989; M C ~ E A R et nl. 1996). Strains CH2252 [pol30- 104 (rho")] and CH2253 [POL30 (rho")] were grown to log phase at 35"; and each of the strains was labeled with "H-uracil or I4C-uracil for 2 6 hr at 35". Both alka- line and neutral profiles of the DNA from POL30 and pol?0 cells were very similar when the cells were grown at 35" (permissive temperature; Figure
7,
A and B). When the cells were shifted to 14" after radio-labeling, however, the profiles differed. Whereas poU0 and POL30 strains exhibited indistinguishable DNA profiles in neutral sucrose gradients (peaks appeared in fraction 15 for both POL3Oand pol?Ocells), in alkaline gradients the peak fraction for POL30 DNA is 12 and the peak fraction for pol30 DNA is 10. To determine if this differ- ence in peak fractions is significant, we calculated the number-average molecular weights of the DNA, and we found that DNA from p o U 0 cells has a 22% lower value(2.1 X 10') than the DNA from POL30 cells
(2.7
X488 N. S. Amin and C. Holm
3 5 "
2 5 "
.01
;
I I I0 100 200
U V D o s e ( J / m 2 )
100
1 0
1
. I
.o 1
0 100 200
U V D o s e ( J / m 2 )
100
10 %
._
-
.-
e
n
m l
ae
-
>
.1
.01
0 10
.01 I I
I I
20 30 40 0 10 20 30 40
M i n u t e s i n 0.3% MMS M i n u t e s i n 0.3% MMS
100
1 0
8 .
1
. I I
0 5 1 5 1 0 20
I . I I .1
0 5 10 1 5 20
G a m m a D o s e ( K r a d ) Gamma Dose (Krad)
POL30
pOl30- 103
-
pO130- 104 rad9-
POL30-
pol30- 103-
pOl30- 104 rad9-
POL30 ~ 0 1 3 0 - 103-
~ 0 1 3 0 - 104-
rad52FIGURE 6.-Sensitivity of pol30 strains to DNA damaging agents. Strains CH2165 ( P O L 3 0 ) , CH2163 (poZ3@103), CH2161
A
In Vivo Analysis of PCNA
Neutral Gradients
489
150
z
V
I 100-
m
0 5 10 15 2
Fraction
B
Alkali1
0 5 10 15 2
Fraction
0 5' 10
15
2Fraction
e Gradients
-400 800-
.-
KX30. 700
-
+ pd3O-104-300 600
-
500
-
-200400
-
300
-
-1 00
200
-
1
-0 100 I
,
0 5 I 10 I 15 I 2Fraction
BOO
600
z
400 0
V
d
-
200
3
500
400
300
h
VW
200
*
7
100
0
f -
MMS
Recovery
L
600
400
z
V
I
m 200
0
0 5 10 15 20
300
200
100
0
~ 1 3 0 -
7
04
oo--t 0 hr
-
2 4 hr.
300
-
-300
200
-
-200 0
u
100
-
-1 00 c
Fractim Fraction
FIGURE 7.-Velocity sedimentation analysis of chromosomal DNA in pol30 and POL30 strains. Strains CH2253 [POL30 (rho')] and CH2252 [po130-104 (rho')] were grown to log phase at 35" before labeling with either 'H or I4C uracil. Where indicated, cells were additionally exposed to 0.3% MMS for 5 min, and then allowed to recover before sedimenting DNA in alkaline gradients. Chromosomal DNA was isolated from the labeled cells, and it was sedimented on alkaline or neutral 15-30% sucrose gradients. (A) Neutral gradient profiles of DNA isolated from strains grown at 35" or 14". (B) Alkaline gradient profiles of DNA isolated from strains grown at 35" or 14". (C) Alkaline gradient profiles of DNA isolated from strains exposed to 0.3% MMS for
5 min and allowed to recover for 0 or 24 hr at 14".
490 N. S. Amin and C. Holm
cells were then exposed to 0.3% MMS for 5 min, and samples were shifted to 14" or 35" to allow for recovery from MMSinduced damage. The neutral DNA profiles were similar for both POL30 and pol30 strains after incu- bation at 35" or 14" (data not shown), indicating an insignificant level of double-strand breaks. The alkaline DNA profiles from cells recovering at 35" were also simi- lar for POL30 and pol30 strains (data not shown). How- ever, when the cells were incubated at 14" after MMS treatment, we saw minimal recovery from MMS-induced damage in pol30 strains. Whereas the peak fraction of DNA from strain CH2253 [POL30 (rho')] shifts from fraction 5 to fraction 9 after 24 hr at 14", the peak fraction from strain CH2252 [pol30 (rho")] shifts only from fraction 4 to fraction 5 (Figure
7C).
Thus, pot30mutants are defective in processing DNA damage that has been caused by alkylation in vivo, as revealed by an accumulation of single-stranded breaks but not double- stranded breaks.
DISCUSSION
We have shown that yeast PCNA is required for effi- cient DNA replication and DNA repair through the isolation and analysis of cold-sensitive (Cs-) and MMS- sensitive (Mms') alleles of the yeast PCNA gene (POL30). The Cs- mutations are clustered in the inter- domain region of the PCNA monomer structure, but the Mmss mutations are scattered around the protein. The Cs- pol30 mutants undergo cell cycle arrest after completing the bulk of DNA replication, but progres- sion through S phase is markedly delayed at the restric- tive temperature. The arrest phenotype of pol30 cells is dependent on the RAD9 DNA damage checkpoint, suggesting the presence of DNA damage. Consistent with this possibility, pol30 cells have high spontaneous mutation frequencies, and they accumulate single- stranded breaks in their DNA at the restrictive tempera- ture.
The phenotypes of the POL30 mutants recovered in this study show some correlation with previous muta- tional analysis of the POL30gene (AWAGARI et al. 1995). Earlier studies revealed that while a majority of alanine scanning mutations have no effect in vivo, a few cause sensitivity to the DNA damaging agents UV and MMS. We also recovered Mmss mutants, but our Mms-sensitive mutants do not appear to be sensitive to UV radiation. Our Cs- mutants, however, exhibit Mms sensitivity at all temperatures, and, interestingly, slight UV sensitivity only at the semi-permissive temperature. This apparent difference in sensitivity to DNA damaging agents may be a result of different assay conditions, or it may reflect real differences in the types of mutants recovered. Taken together, these observations suggest that PCNA plays a role in repairing DNA damaged by either UV light or MMS, and that its efficacy in the processes can be differentially affected by mutation.
The striking clustering of our Cs- alleles in the inter- domain region of the PCNA monomer suggests that this region is functionally important in vivo. One of two mutations (pol3O-6) causing a growth defect isolated by
AWAGARI et al. (1995) also maps to this region. The other (cold-sensitive) growth-defective mutation ($0130-
52) isolated by AWAGARI et al. (1995) maps to a structur- ally similar portion of the molecule: the monomer- monomer interface, which differs from the interdo- main region mainly in its lack of an overhanging pep- tide loop. The interdomain connecting loop has been speculated to be important in the interactions of PCNA with other proteins, because structural predictions sug- gest that it can provide flexibility (KRISHNA et al. 1994). Similar flexibility may be provided by the conserved C- terminal domain of the protein, which is where the majority of the Cs- mutations map. The possibility that the Cs- mutations disturb interactions of PCNA with other proteins is consistent with the general notion that Cs- mutations often perturb protein-protein interac- tions (CANTOR and SCHIMMEL 1980; STRAUSS and GUTHRIE 1991). The clustering of the Cs- mutations is in sharp contrast to the wide distribution of both the previously isolated and the newly isolated Mms" pol30
mutations (Figure 8; AWAGAIU et al. 1995). Thus, it ap- pears that although slight perturbations anywhere in PCNA may affect its ability to metabolize alkylated DNA, the interdomain region may be critical for an additional essential function. An obvious possibility is that muta- tions in this region specifically perturb DNA replica- tion.
Given that PCNA interacts with other members of the replication machinery, how do the various mutations identified in yeast and human PCNA affect these inter- actions? The in vivo interaction of W C l (CDC44) and PCNA (POL30) has been previously shown in yeast through the identification of pol30 mutations that sup- press the phenotypes of rdc44 mutants (MCALEAR et nl.
1994; AWAGARI et al. 1995). Although it uses a different genetic test, the synthetic lethality of the Po130 muta- tions with cdc44 in our study also suggests a functional interaction between RFCl and PCNA. It is interesting that neither the suppression nor the synthetic lethality is allele-specific. This result suggests that the Cs- muta- tions alter overall interactions between the proteins, rather than affecting specifically interacting amino acids.
Although targeted mutagenesis in yeast and human PCNA genes has revealed various mutations that affect the interactions with pol
S
and RFC in vitro, only two of the mutations that cause a recognizable phenotype are closely related to those identified in this study (AWA-In Vivo Anal1
234
FIGURE 8.-Predicted locations of amino acids altered by each cold-sensitive, MMSsensitive, or cdc44suppressor muta- tion of yeast PCNA. The a-carbon backbone of the PCNA trimer is shown (KRISHNA et al. 1994), and the location of mutations conferring the indicated phenotypes are high- lighted as black circles. Numbers indicate the amino acid that has been changed in each mutation. The Cs- mutations show tight clustering (this study) with the exception of the muta- tion in amino acid 115 (AWAGAFU et al. 1995). The Mms” conferring mutations are more loosely distributed around the monomer. It should be noted that many of the Mms5 muta- tions (AYYAGARI et al. 1995) are also UV-sensitive. In addition, all CsC mutations are also Mms’; however, the CsC mutations have not been shown here with the Mms> mutations for rea- sons of clarity. Those PCNA mutations identified as suppres- sors of Cs- cdc44 ($1) mutations are depicted together; these mutations were derived from two separate studies (M&.EAR et al. 1994; AWAGAFU et al. 1995). For clarity, only one of the two adjacent alanine mutations from AWAGARI et al. (1995) are shown in this diagram. This figure is modeled after Figure
1 in AYYAGAFU et al. (1995).
served in yeast, and we have identified a Y114K amino acid change in one of the Mms” mutants (po130-105) in our study. Although, the Y114K mutant yeast protein does not affect trimerization in our studies (unpub- lished data), it results in synthetic lethality when com- bined with a $1 mutation (cdc44-5). A second mutation isolated by FUKUDA et al. K13A in human PCNA was shown to affect DNA synthesis by pol
6
in vitro. In our studies, a K13E mutation in yeast PCNA (po130-114) causes MMS sensitivity alone, without any associated DNA synthesis defects. Overall, the observation that res-idues at the Cterminal tail of human PCNA are re- quired for the stimulation of RFC activity, but not pol
6
activity in vitro is interesting (Fukuda et al. 1995), because five of our eight Cs- mutations are located at the C-terminal.In addition to the interactions of PCNAwith the repli-
isis of PCNA 49 1
cation machinery, it has been suggested that PCNA might be involved in cell cycle regulation through its interactions with cyclins, cyclin-dependent kinases, and p21 (XIONG et al. 1992,1993; ZHANG et al. 1993; FLORES et al. 1994; WAGA et al. 1994). Unfortunately, in vivo evidence for such a role is lacking. Analysis of our cold- sensitive mutants of POL30 did not reveal any specific defects in cell cycle control that were independent of the DNA synthesis defects. It is possible that our screen- ing process did not target cell cycle-specific mutations, or the perturbation of regulatory roles may be subtle compared to effects on DNA synthesis. Alternatively, yeast may differ from other eukaryotes in this area of cell cycle control, because yeast does not need the com- plex regulatory mechanisms necessary in multicellular organisms. Although p21 is known to bind to PCNA in mammalian cells and inhibit DNA replication in vitro, a homologue of p21 in yeast has not been identified. Thus, the role of PCNA in cell cycle control in vivo remains to be elucidated.
How does our in vivo characterization of Po130 mu- tants compare with its previously attributed replication functions from in vitro studies? In vitro studies suggest that pol
6
has appropriate activities to be responsible for the bulk of DNA replication, and pol6
activity is greatly enhanced by its association with PCNA (WAGA and STILLMAN 1994). Thus, it might have been expected that DNA replication would not proceed to completion in Cs- pol30 mutants at the restrictive temperature. In vivo, however, there may be other polymerases that can substitute for the PCNA-dependent polymerase6
activ- ity in yeast. For example, an in vitro study showed that yeast pol E can substitute for yeast or human pol 6 inDNAelongation (LEE et al. 1991). In fact, yeast tempera- ture-sensitive cdc2 (pol